
Abstract
The present survey aimed to investigate flea and tick fauna parasitizing Slovak red fox populations with special emphasis on canine pathogens they transmit. A total of 407 fleas and 105 ticks were collected from 90 red foxes from two geographically distant regions. Seven flea species (Chaetopsylla globiceps, Pulex irritans, Archaeopsylla erinacei, Chaetopsylla rothschildi, Chaetopsylla trichosa, Ctenocephalides canis, and Ctenopthalmus assimilis) and three species of hard ticks (Ixodes ricinus, Ixodes hexagonus, Haemaphysalis concinna) were recorded on sampled animals. Consequently, the DNA of five different pathogen taxa was confirmed in collected arthropod vectors: Bartonella spp. (in P. irritans, Ch. globiceps, and Ct. assimilis), Rickettsia spp. (in A. erinacei, I. ricinus, I. hexagonus, and H. concinna), Anaplasma phagocytophilum (in I. ricinus), Theileria sp. (in Ch. globiceps and H. concinna), and Hepatozoon canis (in I. ricinus and I. hexagonus). Mycoplasma spp., Dipylidium caninum, and Acanthocheilonema reconditum were not found in fleas or ticks in this study.
Introduction
T
While the situation regarding a spread of zoonotic helminths (including Echinococcus multilocularis) by fox population has been explored in detail (e.g., Deplazes et al. 2004), much less is known about the role of red foxes in the transmission of vector-borne pathogens. Fleas and ticks parasitizing red foxes can mainly serve as vectors of bacterial pathogens (e.g., Yersinia pestis, Rickettsia spp., Bartonella spp., Anaplasma spp., Ehrlichia spp., Mycoplasma spp.), and hemoprotozoa (e.g., Hepatozoon spp., Babesia spp., Theileria spp.). Likewise, fleas play an important role as intermediate hosts in the development of canine and human tapeworms (e.g., Dipylidium caninum, Hymenolepis nana, and Hymenolepis diminuta) and filarial worm Acanthocheilonema reconditum (Bitam et al. 2010, Dobler and Pfeffer 2011).
The main aim of the present study was to screen fleas and ticks collected from red foxes for the presence of DNA of several above—mentioned pathogens, to determine the role of arthropods in the eco-epidemiology of vector-borne infections in different regions of Slovakia, Central Europe.
To the best of our knowledge, this is the first monitoring of the presence and diversity of vector-borne pathogens in fleas and ticks parasitizing red foxes in Slovakia. Likewise, information on the diversity of ectoparasites of red foxes living in different regions of Slovakia is still limited. The last large-scale taxonomic study focusing on fleas of wild carnivores was carried out by Cyprich and Lukáš (2006). Therefore, the additional goal of our study was to provide the first complex view on the spectrum of fleas and ticks infesting the red fox populations, with emphasis on regional differences.
Materials and Methods
Fleas and ticks sampling
Fleas and ticks were collected from the carcasses of red foxes delivered to the State Veterinary Institutes (SVI) during the campaign of anti-rabies vaccination effect monitoring, between February and July 2017. Two Institutes joined the research: SVI Košice (south-eastern Slovakia) and SVI Bratislava (south-western Slovakia). All arthropods removed from dead animals were stored in 70% ethanol and transported to the Institute of Parasitology, Slovak Academy of Sciences (IP SAS) in Košice for further investigation. Fleas and ticks were identified to species and sex and/or stage under the stereomicroscope using the morphological keys and published descriptions (Rosický 1957).
The prevalence and mean intensity of an ectoparasitic infestation were not calculated considering that the only data which have been provided were related to the foxes evaluated as positive for the presence of ectoparasites. Moreover, the foxes' corpses were stored frozen for a time what could have a significant impact on the number of ectoparasites found and/or collected.
DNA isolation and molecular analyses
Fleas and unfed ticks were air dried before DNA isolation and individually homogenized. DNA was extracted individually from each flea using alkaline hydrolysis method (Rijpkema et al. 1996).
DNA from engorged ticks was isolated using the DNeasy Blood and Tissue Kit (Qiagen®, Germany) according to the manufacturer's instructions. All DNA samples were stored at −20°C for further analyses.
Subsequently, all DNA samples were screened for the presence of pathogens according to published protocols: Bartonella spp. (Norman et al. 1995, Maggi et al. 2006), Mycoplasma spp. (Varanat et al. 2011), Anaplasma phagocytophilum (Eberts et al. 2011), Babesia spp. (Casati et al. 2006), Hepatozoon canis (Inokuma et al. 2002, Najm et al. 2014), D. caninum (Beugnet et al. 2014), and A. reconditum (Rishniw et al. 2006).
Several positive amplicons from each PCR assay were purified using NucleoSpin® Gel and PCR Cleanup kit (Macherey-Nagel GmbH & Co.) and sequenced.
Obtained electropherograms were manually edited using alignment explorer of MEGA 6 (Tamura et al. 2013). Nucleotide sequences were compared with GenBank entries by Basic Local Alignment Search Tool (BLAST) (Altschul et al. 1990) and submitted to GenBank database.
Results
Fleas and ticks identification
In total, 407 fleas and 105 ticks were collected from 90 red foxes in Slovakia. The most frequent flea species was Chaetopsylla globiceps (n = 223), followed by Pulex irritans (n = 114), Archaeopsylla erinacei (n = 27), Chaetopsylla rothschildi (n = 15), Chaetopsylla trichosa (n = 14), Ctenocephalides canis (n = 13), and Ctenopthalmus assimilis (n = 1).
Five flea species were detected in all surveyed regions: Ch. globiceps, Ch. trichosa, P. irritans, Ct. canis, and A. erinacei. On the contrary, Ch. rothschildi was recorded only in red foxes originated from south-western part of Slovakia; and Ct. assimilis was found only in one fox from the south-eastern region (Table 1).
(n = number of arthropods collected).
Regarding ticks, three species were identified from red fox carcasses, namely Ixodes ricinus (n = 43), Haemaphysalis concinna (n = 36), and Ixodes hexagonus (n = 26). While I. ricinus, the most abundant tick species in Slovakia, was recorded mostly in the eastern part of the country and only two individuals were found in the western region, all H. concinna ticks, except one individual, were collected from foxes originated from south-western Slovakia.
I. hexagonus ticks were distributed more evenly (Table 1).
Molecular analyses
In the present study, the occurrence of five different species of vector-borne pathogens was recorded in fleas and/or ticks removed from red foxes: Bartonella spp., Rickettsia spp., A. phagocytophilum, Babesia/Theileria spp., and H. canis (Table 2). Their geographical distribution is shown in Figure 1.
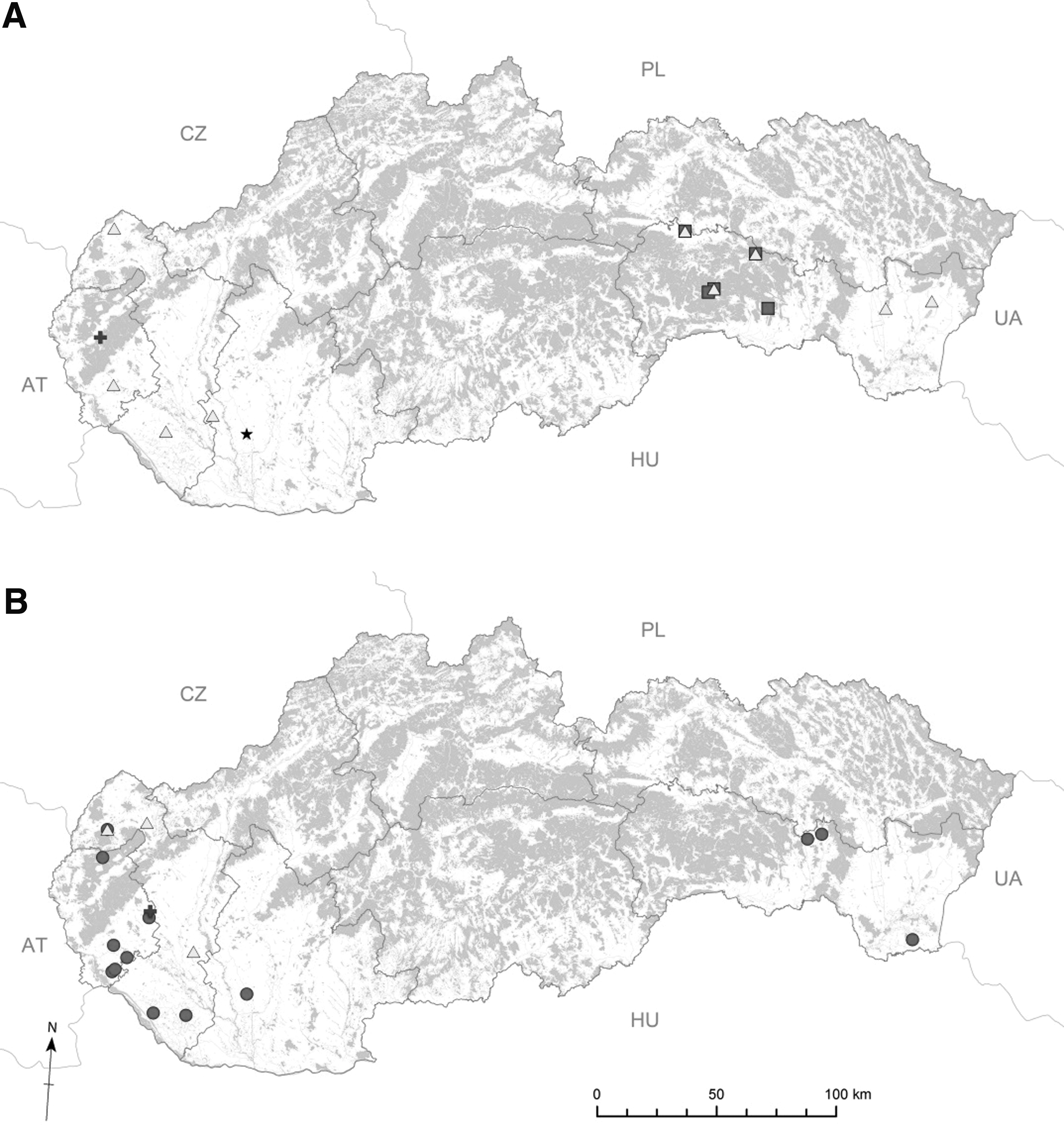
Geographical distribution of bacterial and parasitic pathogens detected in ticks
(n) number of examined ectoparasites.
Bartonella spp. was the most frequently recorded pathogen, and it was detected only in the fleas. P. irritans was the most affected species with this bacterial infection, followed by Ch. globiceps and Ct. assimilis (Table 2). In total, 11 foxes carried Bartonella-infected fleas. Based on the sequencing of 420–780 bp long 16S-23S rRNA gene intergenic spacer region (ITS), 20 sequences were identical to the sequences from the French red fox and P. irritans flea (CP019780; EU191619). One sequence shared 99% similarity with the sequence obtained from the spleen of a Clethrionomys glareolus from Slovakia (KX267690) as well as 98.9% similarity to the Bartonella taylorii strain WM9 (AJ269788). Based on a sequence of a 328 bp long fragment of the citrate-synthase gene (gltA) this sample was again identical to the sequence from a spleen of a bank vole (JQ694012). The other six gltA sequences shared 100% identity with a sequence from a blood sample of Canis latrans (CP019785) and with Bartonella rochalimae isolate BMGH (DQ683195) from a human patient.
Altogether, 14 ticks of three species and four A. erinacei fleas, removed from 12 animals, tested positive for Rickettsia spp. (Table 2). More than half of PCR positive amplicons were sequenced. Only three nucleotide sequences were suitable for further processing. Two of them (MG579967, MG579968), isolated from A. erinacei fleas, were identical and showed 99% similarity with Candidatus Rickettsia senegalensis strain PU01-02 (KF666474) from a cat flea (Ctenocephalides felis) in Senegal and/or Rickettsia felis strain PS120 (AF196973). The third partial nucleotide sequence of a sca4 gene (MG579966), obtained from H. concinna tick collected from red fox in south-western Slovakia, showed 99% similarity with Candidatus Rickettsia kotlanii strain HM-2 (KJ619625) isolated from Haemaphysalis megaspinosa tick in Japan.
Regarding other bacterial pathogens, A. phagocytophilum was detected in one I. ricinus tick from western Slovakia (Table 2). The partial nucleotide sequence of membrane surface protein 2 (msp2) gene, encoding membrane surface protein P44 was compared with GenBank entries and showed 99% similarity with A. phagocytophilum variant 2, strain Norway (CP015376), and/or A. phagocytophilum strain JM (CP006617).
No flea and/or tick included in this survey were positive for hemotropic Mycoplasma spp.
However, hemoprotozoans, Babesia/Theileria spp. and/or H. canis were detected in arthropods removed from red foxes in this study (Table 2). The primers, originally designed for a genus-specific detection of Babesia spp. were used in present work as published by Casati et al. (2006). After 18S rRNA PCR, two samples (one Ch. globiceps flea and one H. concinna tick) tested positive. However, sequencing of positive PCR product from Ch. globiceps did not provide an electropherogram of sufficient quality for further genotyping. In the case of H. concinna tick, BLAST analysis of 18S rRNA nucleotide sequence (MG214907) showed 99% similarity with a complete 18S rRNA sequence of Theileria sp. 3185/02 strain isolated from the spleen of Spanish roe deer (DQ86642) and/or Theileria capreoli clone (KJ188219; KJ188207-209) from west-central China.
H. canis was detected exclusively in ticks from eastern Slovakia. In total, one I. hexagonus and five I. ricinus ticks, removed from five foxes, carried DNA of this protozoon (Table 2). Nucleotide sequences from ticks were identical with each other and with isolates from dogs and/or foxes from all around the world, for example, with H. canis from foxes from Hungary (KJ572979), Slovakia (KX761384; KX887326) and/or Czech Republic (KU893120; KY021180; KU893122; KY021177).
Despite the molecular screening of all samples, canine helminths D. caninum, and A. reconditum were not found in blood-sucking ectoparasites removed from red foxes included in this study.
Simultaneous infections caused by two different species were also confirmed. Three I. ricinus ticks, collected from three different foxes, were coinfected with Rickettsia spp. and H. canis.
Nucleotide sequences submitted to GenBank database
Nucleotide sequences obtained in the present study were deposited in GenBank under the following accession numbers: 18S rRNA—Theileria sp. isolate 458 Hcon/Fox (MG214907); H. canis isolate 42 Iric/Fox (MG214908); H. canis isolate 327 Iric/Fox (MG253001); H. canis isolate 328 Iric/Fox (MG253002); H. canis isolate 329 Iric/Fox (MG253003); H. canis isolate 331 Iric/Fox (MG253004); 44 kDa msp2 gene A. phagocytophilum isolate 443 Iric/Fox (MG334167); surface cell protein 4 (sca4) gene Rickettsia sp. isolate 466 Hcon/Fox (MG579966); Rickettsia sp. isolate 307 Erinacei/Fox (MG579967); and Rickettsia sp. isolate 434 Erinacei/Fox (MG579968)
Discussion
A diversity of fleas and ticks removed from red foxes
During the recent decades, a lot of attention has been paid to arthropods as vectors of serious viral, bacterial and protozoal pathogens and parasites. Nevertheless, as the mosquito-borne and tick-borne diseases have been at the center of scientific attention for a long time, much less is known about fleas and their actual role in the transmission of pathogens. Moreover, in many European countries, only a limited number of studies concerning the diversity of the flea fauna exist.
In Slovakia, Central Europe, the last relevant data related to fleas parasitizing red foxes are dated in the 1950s–1980s (Rosický 1950, Dudich 1982). More recent data aimed at many host species, including foxes, were published by Cyprich and Lukáš (2006).
In their study, as well as in our survey, Ch. globiceps, a specific and the most widespread ectoparasite of red foxes, was the most abundant species. This flea was recorded with relatively high prevalence also in neighboring Hungary (Sréter et al. 2003) and Romania (Foley et al. 2017).
The second most numerous flea species recorded in Slovak red fox population was P. irritans (human flea). In the past, this species was only sporadically found in wild carnivores from Slovakia. However, today P. irritans is being found regularly in various species of domestic and wild animals (Cyprich and Lukáš 2006).
Ch. rothschildi collected from six foxes originating from south-western Slovakia is a relatively rarely occurring species in Europe and it has not been collected from red foxes during recent studies (Foley et al. 2017). It is known from France (Beaucournu 1973), but its current distribution is still unclear. In Slovakia, this flea parasitizes a broad spectrum of wild—living carnivores; most often it has been found on mustelids, for example, beech marten (Martes foina), European pine marten (Martes martes), European polecat (Putorius putorius), steppe polecat (Putorius eversmanni), European badger (Meles meles), or in the wildcat (Felis silvestris). Regarding red foxes, Cyprich and Lukáš (2006) reported the presence of Ch. rothschildi in one female from the western part of the country in 1992.
Ct. canis, the most common flea of domestic dogs, is regularly observed in red fox populations throughout Europe. The other, quite abundant flea species, the hedgehog flea (A. erinacei), is considered a nonspecific for red foxes. We found this species in several regions of Slovakia. The main host of this flea is the European hedgehog (Erinaceus europaeus); however, apparently the flea species is abundant also in the fox population. Surprisingly, the most recent complex study from Romania did not confirm this flea species in a local red fox population. On the contrary, the survey from Romania brought the first record of Ct. assimilis assimilis in red fox (Foley et al. 2017). Likewise, this species, which is quite common in the populations of small mammals, was detected in red fox from the eastern part of the country during the present study. Furthermore, even though Foley et al. (2017) stated that this species has previously not been reported in carnivores, in Slovakia Ct. assimilis was collected from several carnivorous species, such as the beech marten (M. foina), the stoat (Mustela erminea) and/or the least weasel (Mustela nivalis) (Cyprich and Lukáš 2006).
The spectrum of fleas recorded in red foxes from this study is similar to the previously published data from Slovakia (Rosický 1957, Cyprich and Lukáš 2006, Kočišová et al. 2006). However, interesting was the absence in the present study of the badger flea (Paraceras melis) which is considered to be the second most abundant flea species parasitizing red foxes (Rosický 1957). Paraceras melis together with Ch. trichosa is the most common flea species present on European badger (M. meles). Despite the absence of P. melis, 14 individuals of a badger flea, Ch. trichosa, have been collected. This species is quite frequent in Europe (Lassnig et al. 1998, Sréter et al. 2003, Karbowiak et al. 2016, Foley et al. 2017).
Regarding ticks, three species were removed from red foxes in the current survey: I. ricinus, I. hexagonus, and H. concinna. In accordance with a situation in other European countries, it is clear that I. ricinus is the most prevalent tick species infesting red foxes. Likewise, it was found to be a dominant species in red foxes from Austria, Hungary, Germany, or Poland (Lassnig et al. 1998, Sréter et al. 2003, Najm et al. 2014, Karbowiak et al. 2016). I. hexagonus was also found in Germany, Poland, and Romania (Najm et al. 2014, Karbowiak et al. 2016, Sándor et al. 2017). In general, H. concinna, is much less widely distributed in Europe, however, it was reported with relatively high abundance in Hungary, which neighbors the south of Slovakia (Sréter et al. 2003).
Flea-borne and tick-borne pathogens
Fleas and ticks play an important role in the transmission of a broad spectrum of vector–borne parasites and pathogens. Moreover, many ectoparasite species are highly synanthropic and not strictly host-specific, characteristics that may increase the risk of infestation and spread of infections (Bitam et al. 2010). Despite many studies focused on arthropod-borne diseases have been conducted, there is still lack of information regarding the flea and tick vectors parasitizing wild carnivores. Considering the actual trend of a strong urbanization, penetration of wild animals into the urban environment, and their close contact with domestic animals and humans, more detailed epidemiological surveys are needed.
The fleas and ticks collected in this study from red foxes were tested for the presence of DNA of bacteria, protozoa, and helminths.
In total, 21 fleas from three species tested positive for Bartonella spp. Until now, more than 30 Bartonella species and subspecies were described worldwide, and several of them have a considerable zoonotic potential (Paziewska et al. 2011, Kosoy et al. 2012). Fleas are the main vectors responsible for bartonellae transmission among hosts (Gutierrez et al. 2015). This is the first screening of bartonellae in ectoparasites collected from foxes in Slovakia. Based on a sequencing of two genetic markers (ITS and gltA), we have confirmed the presence of zoonotic B. rochalimae and B. taylorii-like genotypes in three flea species. B. rochalimae was for the first time isolated from a human blood sample (Eremeeva et al. 2007). Since then, B. rochalimae and other closely related species have been recovered from a variety of domestic or wild animals and their parasites worldwide (Henn et al. 2009, Gundi et al. 2012, Buffet et al. 2013, Millan et al. 2016, Bai et al. 2016). B. taylorii was found in one Ct. assimilis flea, which is a euryxenous parasite, with a broad host range, mainly parasitizing species of the genus Microtus. Therefore, the presence of rodent-associated B. taylorii DNA in this flea can be explained as a residual DNA acquired during the previous blood meal, probably on a rodent. In Europe, many studies focused on detection of Bartonella spp. in fleas and ticks infesting red foxes have been conducted. The most frequently, bartonellae were reported from P. irritans fleas (Sréter-Lancz et al. 2006a, Márquez et al. 2009).
Less frequently, bartonellae are being recorded in ticks parasitizing foxes, despite a great number of these arthropods having been examined (La Scola et al. 2004). Similarly, in our study, no tick tested positive for Bartonella spp., despite the fact that DNA of Bartonella spp. was confirmed in I. ricinus and D. reticulatus ticks from Poland, which neighbors Slovakia to the north (Podsiadly et al. 2009). The possibility that Bartonella species are transmitted by ticks is not yet proved (Telford and Wormser, 2010).
The most frequently reported rickettsiae in central and western Europe, belonging to the spotted fever group, are Rickettsia slovaca associated with Dermacentor marginatus and D. reticulatus ticks and Rickettsia helvetica transmitted by I. ricinus.
A flea-borne bacterium, R. felis, belonging to the transitional group of rickettsiae is transmitted by the cat flea (C. felis), which is considered to be its most competent vector (Brouqui et al. 2007).
Many Rickettsia species are considered to be actively multiplying symbionts, but they can also be attached to the surface as contaminants. Therefore, rickettsiae can be transmitted during the blood meal as well as mechanically, which is considered an alternative route of transmission (Lawrence et al. 2015).
The results of our study confirmed rickettsial DNA in three tick species (I. ricinus, I. hexagonus, and H. concinna) and one flea species (A. erinacei). In total, 14 ticks and four fleas carried Rickettsia spp. DNA. Two isolates (MG579967, MG579968) from the hedgehog fleas were 99% similar with a R. felis-related Candidatus R. senegalensis strain identified in a cat flea from Senegal (Mediannikov et al. 2015). The second unexpected record of this species was reported from Australia that same year. Interestingly, this species was detected only in cat fleas, which were not surface-cleaned before DNA extraction, suggesting this species might be an external surface contaminant. The study of R. felis-related bacteria is extremely important, considering that these emergent pathogens have a cosmopolitan distribution and are responsible for a flea-borne spotted fever. An eventual zoonotic potential of Cand R. senegalensis and the implications of its detection for public health need to be further investigated. Moreover, the cat flea lacks host specificity and is able to bite humans, thus representing a potential risk of zoonotic transmission of rickettsiae (Torina et al. 2013).
Rickettsia sp. (MG579966) detected in H. concinna removed from red fox was closely related to Candidatus Rickettsia kotlanii (previously Rickettsia RpA4) strain (KJ619625) recorded in H. megaspinosa ticks from Japan. Until now, this candidate species was detected in a questing H. concinna from Hungary, (Sréter-Lancz et al. 2006b). Despite lack of information related to the pathogenicity or virulence of this Rickettsia sp., phylogenetic analysis based on several gene loci, showed a close relationship with some zoonotic species (Andoh et al. 2014). Moreover, both tick species, H. concinna and H. megaspinosa, occasionally attack humans and might represent a risk of infection.
The presence of Rickettsia sp., closely related to Cand. Rickettsia kotlanii identified for the first time in the same tick species in Hungary might suggest spreading of pathogens through the migration of animal hosts between the countries.
A. phagocytophilum was confirmed only in one I. ricinus tick in the present study. No flea tested was positive. In Europe, this bacterium is being detected quite often and the prevalence of infection in questing ticks ranges from 0.4% to more than 15% (Stuen et al. 2013). Bacteria have been confirmed in a wide spectrum of animal hosts worldwide (Stuen 2007), and the ability of red foxes to serve as reservoir hosts has already been demonstrated (Pusterla et al. 1999, Hulínska et al. 2004, Karbowiak et al. 2009). The prevalence of infection in this sentinel species ranges from 2.55% to 16.6% (Sixl et al. 2003, Sréter et al. 2003, Karbowiak et al. 2009, Ebani et al. 2011, Härtwig et al. 2014, Tolnai et al. 2015). However, the data on the prevalence of pathogens in ectoparasites removed from wild animals are relatively scarce (Sréter et al. 2004, Dumitrache et al. 2015). In Slovakia, several molecular studies concerning the prevalence and genetic heterogeneity of A. phagocytophilum in ticks and reservoir hosts have been carried out (Špitálska and Kociánová 2002, Derdáková et al. 2003, 2011, 2014, Nováková et al. 2010, Víchová et al. 2010, 2011, 2014a, 2014b, 2016, Majláthová et al., 2011, Pangrácová et al. 2013, Blaňarová et al. 2014, Svitálková et al. 2015, Reiterová et al. 2016, Slivinska et al. 2016).
Similar to previous studies from Hungary and Switzerland, we failed to detect hemotropic mycoplasmas in ectoparasites removed from red foxes (Willi et al. 2007, Hornok et al. 2010).
The molecular screening of fleas and ticks, collected from red foxes, for the presence of H. canis directly links to our latest research aimed at detection of this protozoan parasite in dogs (1.0%) and red foxes (17.1%) in Slovakia (Miterpáková et al. 2017). H. canis was widely distributed primarily in the south-eastern region of the country. Likewise, similar results were recorded in the present study; H. canis was detected only in I. hexagonus and I. ricinus ticks from this region. It is important to emphasize that the permanent occurrence of R. sanguineus s.l. (the main vector of H. canis in Europe) has not been confirmed in Slovakia yet. On the contrary, a quite high prevalence of canine hepatozoonosis in the fox population expressly points to the long-term circulation of the parasites in this part of Europe. The same phenomenon is observed in other European countries. For example, the latest research from Romania revealed that H. canis was present primarily in I. ricinus ticks collected from dogs and foxes (Andersson et al. 2017). Likewise, in Germany, the presence of Hepatozoon spp. was observed in four different tick species, with the highest positivity in I. ricinus ticks (over 14.0%) (Najm et al. 2014). Moreover, DNA of H. canis was confirmed in questing I. ricinus ticks from western Slovakia as well as from Czech Republic (Hamšíková et al. 2016). Therefore, despite the results of an experimental study published by Giannelli et al. (2013), which reported evidence that I. ricinus cannot act as a vector of H. canis, this possibility should not be completely rejected.
Using primers designed for detection of 18S rRNA fragment of Babesia spp., one H. concinna tick tested positive. The nucleotide sequence obtained from this tick was 99% similar with a complete 18S rRNA sequence of Theileria sp. isolated from the spleen of a Spanish roe deer (DQ86642) and/or Th. capreoli clones obtained from red deer and sika deer from northwestern China (KJ188219; KJ188207-209). Detection of the Theileria species associated with wild ruminants in a tick removed a from red fox was surprising but can reasonably be justified. There is an evidence of two genetically distinct genotypes of Th. capreoli, strongly associated with the host species. The genotype “elaphi-CE1” is associated with red deer and fallow deer, whereas the “capreoli-CE1” genotype is exclusively associated with roe deer (Hornok et al. 2017). With the highest probability, the vector of both genotypes is H. concinna, which is very frequently found on wild cervids. Moreover, Theileria was confirmed in immature as well as adult H. concinna ticks from vegetation in Hungary (Hornok et al. 2012). Considering that Theileria spp. are transmitted transstadially in the tick vectors, it is likely, that H. concinna from our study took a blood meal on a wild ruminant before biting the red fox.
Red foxes are considered to be the main reservoirs of Babesia vulpes (Baneth et al. 2015, Koneval et al. 2017). These blood parasites have been recorded in several ixodid tick species, but obviously their presence in fleas is less frequent.
In addition, all ectoparasites removed from red foxes in this study were screened for the presence of DNA of two helminth species: the tapeworm D. caninum and nematode A. reconditum. None of the examined arthropods tested positive for D. caninum. Although this dog tapeworm is widely distributed among dogs in many countries, its occurrence in Europe is rather sporadic.
Similarly, a filarial nematode, A. reconditum, was not detected in potential flea vectors. However, these results are not surprising, considering that A. reconditum has not been detected in definitive hosts, dogs, or red foxes, despite the high number of animals tested in Slovakia (Hurníková et al. 2015, Miterpáková et al. 2016).
Conclusions
Rising population density and growing urbanization of fox population are hazardous trends, particularly in terms of transmission to various infectious agents. Altogether, seven flea species and three hard-tick species were recorded on sampled red foxes from Slovakia. The occurrence of five different species of vector-borne pathogens was recorded in fleas and ticks removed from foxes. Some of them have a zoonotic potential and represent a risk for animal and human health.
Footnotes
Acknowledgments
Study was realized within the frame of project “Centre of Excellence for Parasitology” no. 26220120022 supported by operating program “Research and Development” funded by European Fund for Regional Development (0.4), and projects of the Scientific Grant Agency of the Ministry of Education of the Slovak Republic and Slovak Academy of Sciences VEGA No. 2/0018/16 and No. 2/0126/16.
Author Disclosure Statement
No competing financial interests exist.