
Abstract
Plague is a lethal zoonotic disease associated with rodents worldwide. In the western United States, plague outbreaks can decimate prairie dog (Cynomys spp.) colonies. However, it is unclear where the causative agent, Yersinia pestis, of this flea-borne disease is maintained between outbreaks, and what triggers plague-induced prairie dog die-offs. Less susceptible rodent hosts, such as mice, could serve to maintain the bacterium, transport infectious fleas across a colony, or introduce the pathogen to other colonies, possibly facilitating an outbreak. Here, we assess the potential role of two short-lived rodent species, North American deer mice (Peromyscus maniculatus) and Northern grasshopper mice (Onychomys leucogaster) in plague dynamics on prairie dog colonies. We live-trapped short-lived rodents and collected their fleas on black-tailed (Cynomys ludovicianus, Montana and South Dakota), white-tailed (Cynomys leucurus, Utah and Wyoming), and Utah prairie dog colonies (Cynomys parvidens, Utah) annually, from 2013 to 2016. Plague outbreaks occurred on colonies of all three species. In all study areas, deer mouse abundance was high the year before plague-induced prairie dog die-offs, but mouse abundance per colony was not predictive of plague die-offs in prairie dogs. We did not detect Y. pestis DNA in mouse fleas during prairie dog die-offs, but in three cases we found it beforehand. On one white-tailed prairie dog colony, we detected Y. pestis positive fleas on one grasshopper mouse and several prairie dogs live-trapped 10 days later, months before visible declines and plague-confirmed mortality of prairie dogs. On one black-tailed prairie dog colony, we detected Y. pestis positive fleas on two deer mice 3 months before evidence of plague was detected in prairie dogs or their fleas and also well before a plague-induced die-off. These observations of plague positive fleas on mice could represent early spillover events of Y. pestis from prairie dogs or an unknown reservoir, or possible movement of infectious fleas by mice.
Introduction
Outbreaks of emerging and re-emerging infectious diseases are challenging to predict. Pathogen dynamics can vary across different host communities, vectors (Mwangangi et al. 2013), and environmental conditions (Harvell et al. 1999, 2002). Because of these variable parameters, it is important to consider host, vector, and environmental determinants at the local level, before pathogen dynamics are extrapolated between areas. Here, we investigated the role of two short-lived rodent species, Northern grasshopper mice (Onychomys leucogaster) and North American deer mice (Peromyscus maniculatus) in plague dynamics on prairie dog (Cynomys spp.) colonies. We assessed whether mice abundance, prevalence of flea infestation, and the presence of Yersinia pestis in fleas collected from mice were related to plague die-offs in black-tailed prairie dog colonies (Cynomys ludovicianus) in Montana and South Dakota, white-tailed prairie dog colonies (Cynomys leucurus) in Utah and Wyoming, and Utah prairie dog colonies (Cynomys parvidens) in Utah.
Plague is a (re)emerging zoonotic disease (Butler 2013) associated with wild and peri-domestic rodents throughout the world. In the United States, outbreaks can decimate local populations of susceptible prairie dogs, a keystone species of grassland ecosystems (Barnes 1982, Cully et al. 1997, Pauli et al. 2006). The causative agent, Y. pestis, can be transmitted via several routes: inhalation of infectious droplets (Doll et al. 1994, Agar et al. 2009), consumption of infectious tissues (Thomas et al. 1989), and bites of infectious fleas (Eskey and Haas 1939), the primary route. In wild rodent communities with plague, an increase in host abundance could aid pathogen transmission; more hosts could lead to an increase in flea abundance, increased host contact, as well as increased movement between rodent burrows. In the United States, plague outbreaks on prairie dog colonies have been associated with higher prairie dog abundance and colony size (Stapp et al. 2004, Brinkerhoff et al. 2010), but neither is predictive for plague outbreaks.
Short-lived rodents living on and near prairie dog colonies could serve several roles in plague ecology, from spillover to amplifying host. Variable susceptibility to Y. pestis infection and the ubiquitous presence of deer mice implicated this species as a potential maintenance host for the bacterium in early plague research (Holdenried and Quan 1956, Lechleitner et al. 1968), However, others found no evidence for active deer mouse involvement in plague outbreaks (Salkeld and Stapp 2008) and Y. pestis had not been detected in fleas collected from mice before plague-induced prairie dog die-offs (Anderson and Williams 1997, Cully et al. 1997, Ubico et al. 1988, Holmes et al. 2006, Brinkerhoff 2008, Thiagarajan et al. 2008a, Stapp et al. 2009).
Grasshopper mice also vary in their susceptibility to plague (Thomas et al. 1988). This species has been implicated as a potential transporter of infectious fleas across and between some prairie dog colonies (Salkeld et al. 2010) as they move between several prairie dog burrows each night (Kraft and Stapp 2013) and carry many flea species, including prairie dog fleas (Stapp et al. 2009). In a simulation model, the number of prairie dogs infected with plague on colonies in northern Colorado increased dramatically when the threshold density of grasshopper mice was reached (Salkeld et al. 2010). It was hypothesized that their large home range provided the movement of fleas between prairie dog burrows needed for effective Y. pestis transmission. On those prairie dog colonies, high grasshopper mouse abundance preceded plague outbreaks, and grasshopper mice carried significantly higher prairie dog flea loads during outbreaks (Stapp et al. 2009).
In contrast to grasshopper mice, prairie dog fleas and plague-positive fleas have rarely been detected on deer mice (Anderson and Williams 1997, Holmes 2003, Thiagarajan et al. 2008a,b) or other short-lived rodent species, which makes these other species unlikely transporters of infectious fleas. The hypothesis that grasshopper mice play a significant role in plague dynamics of prairie dogs was based on studies that took place on colonies where the short-lived rodent community is dominated by grasshopper mice. However, not all prairie dog colonies are dominated by grasshopper mice (Anderson and Williams 1997, Cully et al. 1997, 2010, Brinkerhoff et al. 2010, Maestas and Britten 2017), and it is possible that other short-lived rodent species could play a similar role.
To assess the role of short-lived rodents in plague dynamics, especially on colonies not dominated by grasshopper mice, we trapped and sampled small rodents other than prairie dogs, annually (2013–2016) on black-tailed, white-tailed, and Utah prairie dog colonies included in a study to test the effectiveness of the oral sylvatic plague vaccine (SPV) (Rocke et al. 2017). We hypothesized that mouse abundance would be higher the year before plague outbreaks in prairie dogs. We also hypothesized that if short-lived rodents were transporting plague-positive prairie dog fleas across the colonies, we would (1) detect higher prevalence of flea infestation, (2) more plague-positive fleas, and (3) more prairie dog fleas (Oropsylla spp.) on short-lived rodent hosts during plague outbreaks.
Methods
Field collection
Animal trapping and sampling procedures (EP130214) were reviewed and approved by the Institutional Animal Care and Use Committee of the USGS National Wildlife Health Center (NWHC).
Study area and plots
Mice were trapped on 17 paired plots (34 plots) on prairie dog colonies in six study areas (Buffalo Gap National Park, SD [BGSD], Coyote Basin, UT [CBUT], Cedar City, UT [CCUT], Charles M. Russell National Wildlife Refuge [CMR], high elevation, UT [HEUT], and Pitchfork Ranch, WY [PRWY]) that were part of the study to test the effectiveness of the oral SPV in prairie dogs (Table 1, see Rocke et al. 2017, Bron et al. 2018).
Flea Infestation Prevalence of Deer Mice from 2013 to 2015
The number of deer mice and Western harvest mice sampled (n) and flea infestation prevalence (%) per study plot. Gray-shaded blocks indicate that Yersinia pestis DNA was detected in prairie dog fleas or plague-killed prairie dog carcasses were found (Bron 2017, Rocke et al. 2017).
CMR-1 and CMR-2 had plague outbreaks in prairie dogs in 2016.
Plague-infected fleas were detected in abandoned prairie dog burrows on CCUT-1 in 2015, although the colony was believed to have been decimated by flooding after a large rain storm.
BGSD, Buffalo Gap National Grasslands, South Dakota; BTPD, black-tailed prairie dog; CBUT, Coyote Basin Utah; CCUT, Cedar City, Utah; CMR, Charles M. Russell National Wildlife Refuge, Montana; HEUT, high elevation, Awapa plateau, Utah; PRWY, Pitch Fork Ranch, Wyoming is not included because a different flea sampling method was used; UTPD, Utah prairie dog; WTPD, white-tailed prairie dog.
Trapping and sampling
Paired plots were trapped for two to four summers from 2013 to 2016. One study area, PRWY, was trapped twice each season, and we performed additional trap sessions on HEUT and CCUT in the spring of 2015 and summers of 2014 and 2015 (Supplementary Table S1). Briefly, Sherman traps were set in a 10 × 10 grid with traps spaced 15 meters apart (11 × 12 in Wyoming, spaced 16 meters apart); they were set at sunset and checked at sunrise for three to five consecutive nights. Animals were anesthetized with isoflurane and combed for fleas, except for PRWY. In PRWY, animals were inspected for fleas and anesthetized and combed if fleas were observed. Fleas were placed in a 2% saline solution or 70% ethanol and stored at −20°C until identification.
Y. pestis: flea infection
Fleas were identified to species using morphological keys (Hubbard 1947, Furman and Catts 1982) and revisions (Stark 1970, Lewis and Haas 2001, Lewis 2002, 2008, Lewis and Jameson 2002), pooled by species per individual rodent (not exceeding 12 fleas per pool), and stored at −20°C until further processing.
DNA extraction
Flea pools were homogenized by bead beating. DNA was extracted using a quick gDNA prepkit according to the manufacturer's instructions (Zymo Research microprep DNA-elution in 10 μL), and a subset of fleas collected in 2013 was extracted by using a miniprep (DNA-elution in 30 μL). DNA was stored at −20°C until DNA amplification. For processing voucher specimens and samples for bacterial culture, see Supplementary Data. Voucher specimens were stored in the University of Wisconsin-Madison Insect Research Collection (
Y. pestis PCR protocol
Real-time PCR screening for 112 base-pairs (bp) of the pim-gene on the PcP1 plasmid (Genbank AL109969) of Y. pestis was performed (Table 2). See Rocke et al. 2017 and Supplementary Data for details. Samples with background (nonexponential amplification, common in chipmunk Eumolpianus eumolpi and prairie dog fleas Oropsylla spp.) were repeated with a probe-based pim assay, and CMR 2016 samples were tested with this method only. Samples with a threshold cycle (Ct) of 37 or lower were screened for the presence of the caf1 gene on the pFra plasmid (Genbank NC 006323.1, pG8786) (Supplementary Data, Bron 2017, Rocke et al. 2017).
Summary of Primers Used for Yersinia pestis DNA Detection in Fleas
Bold indicate fluorence label (56FAM) and quenchers (ZEN, 31ABkFQ).
Data analysis
Plague outbreak prediction by rodent abundance
We used logistic regression in package lme4 (Bates et al. 2016) to assess whether relative abundance of short-lived rodents (minimum number of animals known alive during three trap nights) was higher the year before plague was detected in prairie dogs. We used active plague per plot as the response variable (0 if no plague was detected and 1 if plague was detected in prairie dog carcasses or prairie dog fleas and prairie dog abundance declined, Rocke et al. 2017), relative short-lived rodent abundance in the previous year, and SPV treatment (to ensure this was not the underlying driver) as explanatory variables. Relative abundance was calculated by dividing abundance of year t by the mean abundance at the study plot throughout the study (years with plague excluded); this standardization allowed us to compare between study areas.
Prevalence of flea infestation and plague
Prevalence of flea infestation (proportion of sampled hosts with at least one flea) from 2013 to 2015 was modeled in a generalized linear model, with a binomial distribution using package lme4 (Bates et al. 2016), to assess whether prevalence was higher on plots where plague had been detected. Only the first sampling record during summer sampling was included for this analysis (i.e., if an animal was sampled), and additional sample occasions were excluded to avoid inclusion of repeated measures. We constructed a global model with explanatory variables that included plague detection, treatment (vaccine or placebo), short-lived rodent abundance (as a measure of rodent density), year, and study area. Backward selection was used to reduce the model; the least significant parameter by likelihood ratio test (LRT, call drop1) was removed if the reduced model's Akaike's Information Criterion corrected for small sample sizes (AICc) was within two AICc or reduced by two AIC or more (Burnham and Anderson 2004). We used standard diagnostic plots to assess model assumptions, tested for overdispersion by Pearson's correlation coefficient (package aods, function gof), and adjusted our model for overdispersion by adjusting the covariance matrix for the estimated variance (Pearson's χ2/df). For prevalence of flea infestation analyses, PRWY was excluded because flea collection methods were different. We repeated this modeling approach for prevalence of flea infestation on deer mice and Western harvest mice only and for the prevalence of Aetheca wagneri on all rodent host and deer mice and Western harvest mice only.
Results
Plague outbreak prediction by rodent abundance
During annual post vaccine-distribution trap sessions over the 4-year study period, 36,500 trap events yielded 8094 small rodent captures, 3625 individuals of 15 species (Supplementary Table S1). On 10 of 17 study pairs plague was detected in prairie dog carcasses or prairie dog fleas in conjunction with a decline in prairie dog abundance during at least 1 year (Table 1, Rocke et al. 2017). During the first 3 days of trapping, 3155 individuals were caught, of which 2487 were deer mice. Combined deer mouse and western harvest mouse abundance per plot did not predict plague detection in prairie dogs or their fleas on that plot the next year (odds ratio [OR] = 1.064, 95% confidence interval [CI]: 0.395–2.865, p = 0.90). Treatment (vaccine or placebo) was also not a significant explanatory variable in the model (OR = 1.004, 95% CI: 0.345–2.923, p = 0.99). Deer mouse abundance in several study areas (CBUT, CMR, HEUT, and PRWY) was higher the year before first plague detection in prairie dog fleas. However, plague was often detected on one plot in a study area whereas other plots in the area remained plague free (Fig. 1). Thus, high deer mouse abundance did not predict plague at the plot level, but it appears associated with favorable conditions for plague invasion in the area.
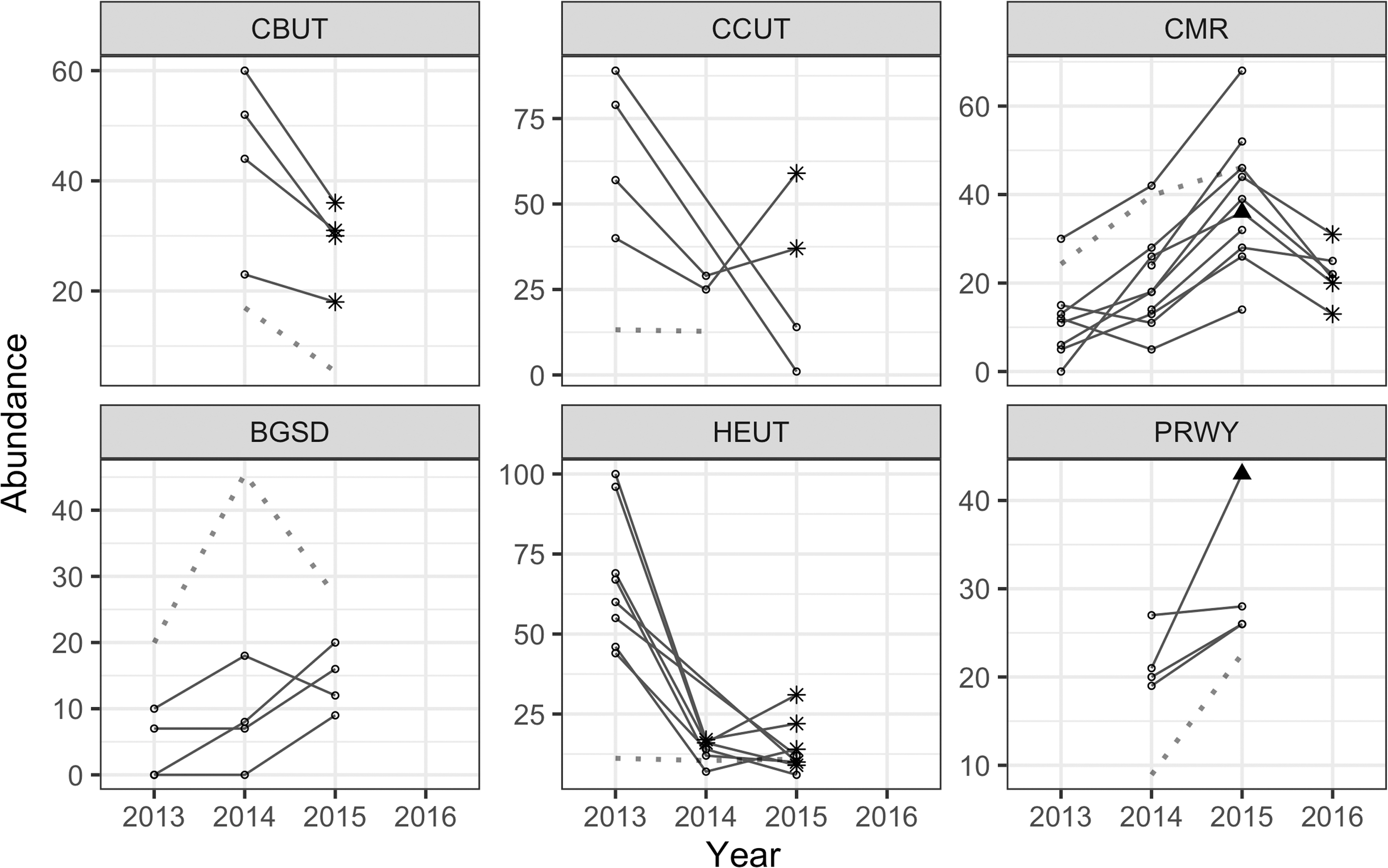
Deer mouse abundance (number of unique individuals caught during three trap nights on all study plots, solid lines) and mean prairie dog abundance (catch per unit effort, Rocke et al. 2017, dashed lines) on CBUT, CCUT, CMR, BGSD, HEUT, and PRWY. Black triangle: Yersinia pestis DNA detection in mouse fleas. Stars: Y. pestis DNA detection in prairie dog fleas. BGSD, Buffalo Gap, South Dakota; CBUT, Coyote Basin area, Utah; CCUT, Cedar City area, Utah; CMR, Charles M. Russell National Wildlife Refuge, Montana; HEUT, high elevation, Utah; PRWY, Pitchfork Ranch, Wyoming.
Grasshopper mouse abundance was low (n = 188) on our study plots (Supplementary Table S1) compared with deer mouse abundance and during other studies describing their possible role in plague dynamics in Northern Colorado (Salkeld et al. 2010). Few individuals (n = 55) were associated with plots that experienced plague outbreaks; thus, analysis was not performed (one grasshopper mouse was caught on a plague-positive plot at CMR in 2016 and nine were caught on a plague-positive plot at PRWY in 2015). This species was most prevalent on black-tailed prairie dog colonies at BGSD where plague was not detected (32% of grasshopper mice caught were caught at BGSD).
Prevalence of flea infestation and plague
In 2013–2015, 3025 hosts were sampled at least once. On first sampling during the summer, 1589 individuals carried at least 1 flea (prevalence 52.5%). The prevalence per plot (PRWY excluded) ranged from 88.9% (CMR5B in 2014, n = 9) to 0 (BGSD2A 2013, n = 3), and mean annual plot level prevalence of flea infestation was 55.1% (n = 80 plot year observations). In our global model, treatment was not a significant predictor of prevalence of flea infestation (LRT p = 0.983), and the reduced model without treatment was our best model (Supplementary Table S2). After correction for overdispersion (dispersion = 2.76), study area and year were significant explanatory variables; CMR had significantly higher flea prevalence than the study area it was compared with in the model (CBUT) (OR = 1.86, 95% CI: 1.09–3.16, p = 0.02), and CCUT prevalence was significantly lower (OR = 0.56, 95% CI: 0.34–0.91, p = 0.019) than CBUT. In addition, prevalence of flea infestation was significantly higher in 2015 compared with 2013 (OR = 1.88, 95% CI: 1.24–2.85, p = 0.003) (Supplementary Fig. S1). The model results were similar for A. wagneri prevalence on all rodent hosts and for prevalence of flea infestation on deer mice only (Supplementary Fig. S1 and Supplementary Table S2).
Prairie dog flea abundance
Of 5713 fleas collected from deer mice, Western harvest mice, and grasshopper mice, 20 (suspect 30) were prairie dog fleas (0.35–0.52%). Nineteen Oropsylla hirsuta were removed from 5 grasshopper mice (maximum flea load 7), 4 deer mice (max. flea load 3), and 1 Western harvest mouse at CMR. One Thrassis francisi was confirmed from a deer mouse on HEUT-1; 10 fleas were identified as Thrassis spp. in 2013 and were likely also T. francisi. In addition, two O. labis were collected from two chipmunks and four Oropsylla idahoensis were collected from a golden-mantled ground squirrel (Callospermophilus lateralis) at HEUT. No common prairie dog fleas were found on mice during plague outbreaks.
Y. pestis flea infection
Including additional sampling on HEUT, PRWY, and CMR plots, we collected 6627 fleas and tested 6123 fleas in 2676 DNA pools (see Table 3 and Supplementary Table S3 for tested specimens). None of the flea pools collected from plots where plague was detected in prairie dogs (718 pools, total of 1441 fleas) were plague positive. Four flea pools were strongly positive for pim and F1 DNA (Table 3): two from CMR and two from PRWY, all collected in 2015, before visible declines of prairie dogs and detection of Y. pestis in prairie dog carcasses.
Sample Size and Maximum Likelihood Estimate of Yersinia Pestis Infection Rate (Determined by DNA Detection) per 100 Fleas Collected from Mice
Mouse flea species also found on prairie dogs are marked with (*), and flea species found on mice that are more commonly associated with prairie dogs are marked with (#). Species that were positive for Y. pestis DNA in our study are bold-faced. The fleas species were collected from prairie dogs at the following plots: A. wagneri BGSD-1A in 2013, CMR-3A, CMR-3B, and HEUT-2A in 2014; C. decipiens HEUT-2B in 2014; E. eumolpi HEUT-4B in 2013, HEUT-1A, HEUT-2B, HEUT-3A, and HEUT-3B in 2014, and HEUT-2A in 2016. Data shared by T.E. Rocke and R.C. Abbott, USGS National Wildlife Health Center, Madison, WI. Point estimates were determined by maximum likelihood estimate or bias-corrected maximum likelihood estimate with the Biggerstaff's PooledInfRate Addin for Excel®. A subset of flea species is shown; see Supplementary Table S3 for additional species.
CI, confidence interval.
From CMR, we tested 2689 fleas, 180 collected in 2013, 570 in 2014, 1570 in 2015, and 369 in 2016. Two pools collected in July 2015 were positive for both pim (threshold cycle [Ct]: 15 and 14) and F1 (Ct: 18 and 17); no other flea pools were plague positive. One pool was collected from a juvenile male and the other from an adult male deer mouse. The flea pool from the juvenile contained two males and two females of Orchopeas leucopus, and the adult male carried two male and four female A. wagneri. From PRWY, we tested 71 fleas, 32 collected in 2014 and 39 in 2015. Two pools collected in June 2015 were positive for both pim (Ct: 14 and 13) and F1 DNA. The positive pools were collected from a female juvenile grasshopper mouse; pools contained three male and three female Pleochaetis exilis, and two female A. wagneri.
Discussion
Deer mice were most abundant in our study, but their abundance and prevalence of flea infestation did not relate to plague-induced prairie dog die-offs. Surprisingly, during plague outbreaks with visible declines and mortality of prairie dogs, we did not detect plague-positive fleas on short-lived rodents. Thus, it appears that deer mice in our study areas were unlikely to have played a similar role as grasshopper mice in Northern Colorado, moving Y. pestis infected prairie dog fleas across the prairie dog colony (Salkeld et al. 2010). Instead, the only plague-positive mouse fleas detected in our study were collected from one grasshopper mouse and two deer mice trapped before plague-induced die-offs that resulted in more than 50% declines in relative abundance on a white-tailed and a black-tailed prairie dog colony, respectively. This provides some limited evidence for the involvement of mice in Y. pestis dynamics on prairie dog colonies, but the significance of this finding remains to be determined. Short-lived rodent fleas are rarely found on prairie dogs, even during plague outbreaks. Of 20,041 fleas collected from 6542 prairie dogs in our same study areas (Russell et al. 2018), only 14 were species associated with short-lived rodents (Table 3).
The observed plague-positive mouse fleas on deer mice, before a plague die-off in prairie dogs, had not been reported earlier. A few studies have reported plague-positive fleas on deer mice during or after plague outbreaks (Holdenried et al. 1955, Lechleitner et al. 1968, Anderson and Williams 1997, Thiagarajan et al. 2008a) and recently in the absence of plague outbreaks in South Dakota (Maestas and Britten 2017). However, few large-scale studies have been conducted in the absence of plague and even in a 46-year surveillance dataset from California (not from prairie dog colonies), samples were most frequently collected during an epizootic, precluding correlation analysis between seroprevalence and subsequent plague outbreaks (Danforth et al. 2018).
On one CMR black-tailed prairie dog colony, we found plague-positive mouse fleas (A. wagneri and O. leucopus) on deer mice in July 2015, without evidence of Y. pestis presence in the colony despite testing prairie dog fleas (n = 35 pools of 56 fleas) collected from 60 prairie dogs 10 days earlier (Rocke et al. 2017). In October 2015, a single plague-killed prairie dog was found (confirmed by USGS National Wildlife Health Center) and the colony had collapsed by the summer of 2016 (observations by R. Matchett, US Fish and Wildlife) (Fig. 2). This does not exclude the possibility that plague was present in prairie dogs at a level that was not detected in 2015, as was observed in PRWY.

Timeline of sampling effort and plague activity on a black-tailed prairie dog colony in Montana (CMR2A) and a white-tailed prairie dog colony in Wyoming (PRWY1B). Prairie dogs represent prairie dog sampling; except in October 2015 in CMR and June 2016 in PRWY, these were observations without systematic sampling. Mice represent mouse sampling effort. (Black animals = no plague detected, light gray = positive carcass(es) and flea silhouette in host = positive flea sample, long-tail = deer mouse, short-tail = grasshopper mouse.
On a white-tailed prairie dog colony at PRWY, we collected plague-positive fleas (A. wagneri and P. exilis) from a juvenile grasshopper mouse in June 2015. Prairie dogs (n = 58) were sampled 10 days after short-lived rodent trapping, and 2 of 35 prairie dog flea pools (n = 42 fleas) tested positive for plague. By the end of July 2015, no visual declines in prairie dogs were observed; however, in June 2016, the colony had collapsed (Boulerice 2017) (Fig. 2). This aligns in part with Stapp et al.'s (2009) observations that grasshopper mice carry plague-positive fleas during plague activity in prairie dogs. In contrast to their study, on our study sites, we found that grasshopper mice were not the most abundant species and their abundance was relatively low compared with Colorado sites described in Stapp et al. (2009).
Plague-positive mouse fleas indicate that the bacterium may be replicating in small rodents to levels that are sufficient to infect feeding fleas. Mouse fleas could have acquired Y. pestis from the same undetermined source as prairie dog fleas, for example, other prairie dogs infected during low-level plague transmission, the start of an outbreak, or another rodent or mammal introducing the bacterium. Interestingly, bloodmeal analysis of the Y. pestis positive fleas by amplicon deep sequencing (described in Bron 2017) yielded no prairie dog DNA, but trace amounts matched DNA sequences from prairie voles (Microtus ochrogaster), a species only trapped rarely on our study areas. We trapped four individuals at CMR prairie dog colonies, but they are more likely to occur off-colony (Holmes et al. 2006).
Local weather patterns varied between and within study areas; unfortunately, no precise measurements are available due to the remote location of most plots. Our first sample year, 2013, was an exceptionally wet year. In HEUT and CCUT, this was associated with highly productive rodent populations, but the wet weather might have also hindered trap success on CMR. In addition, BGSD experienced a very wet and thus green spring in 2013, but no deer mice were captured on those plots that year. The first year of sampling was also the year with the lowest observed flea prevalence. Low flea infestation could indicate that animals had more time to allocate to grooming due to easy accessible food (Eads et al. 2016), but it might also be sampling error as the conditions for animal processing were challenging (i.e., approaching thunderstorms, rain and cold). As with many short-lived rodent studies on prairie dog colonies, our trapping effort was localized (1.8ha grid/colony) and short (3–12 nights, between May and September); thus, the probability of detecting rare events was low. In addition, our abundance estimates were not adjusted for overnight darkness (cloud cover and moon phase) and other plot variables that could have affected trapability of mice (e.g., vegetation type, predator abundance), which may have provided more precise abundance estimates. The distribution of SPV did not have a detectable effect on deer mouse abundance and the protective effect on the mice was minimal (Bron 2017, Bron et al. 2018). Additional concurrent prairie dog and short-lived rodent trapping could help elucidate the possible roles of short-lived rodents in plague ecology; however, these roles likely vary between different prairie dog species and study areas.
In short, Y. pestis maintenance and plague outbreak dynamics on prairie dog colonies are complex, and both have been challenging research topics for decades. The role of short-lived rodent hosts in maintenance and amplification of plague will continue to be questioned, in part due to variation in prairie dog and short-lived rodent communities and their low levels of flea sharing, but also due to feasible, nonexclusive, alternatives such as Y. pestis maintenance in prairie dog meta populations (Salkeld et al. 2016) and survival of the bacterium in soil amoeba (Benavides-Montaño and Vadyvaloo 2017). In our study, we detected a small number of mice with Y. pestis infected fleas before plague-induced die-offs in prairie dogs, an observation not previously reported.
Footnotes
Acknowledgments
The authors thank the field technicians for their stamina, hard work, and enthusiasm. They thank collaborating agencies and their biologists for their support, in particular Randy Matchett and Matthew McCollister (U.S. Fish and Wildlife Service). Colorado Parks and Wildlife graciously provided flea identification training to G.M.B., and the authors thank Dan Tripp and colleagues for their patience and guidance. The authors also thank lab personnel for their tireless assistance and Michael D. Samuel for a critical review of the original article. This work was funded by the Morris Animal Foundation (D14ZO-031, D14ZO-412) and the U.S. Geological Survey. G.M.B. performed this work as a University of Wisconsin–Madison graduate student, supported by U.S.G.S. The use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Data
Supplementary Figure S1
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.