
Abstract
Introduction:
Hantaviruses are a group of globally distributed rodent-associated viruses, some of which are responsible for human morbidity and mortality. Sin Nombre orthohantavirus, a particularly virulent species of hantavirus associated with Peromyscus spp. mice, is actively monitored by the Department of Public Health in California (CDPH). Recently, CDPH documented high (40%) seroprevalence in a potentially novel reservoir species, the cactus mouse (Peromyscus eremicus) in Death Valley National Park.
Methods:
This study was performed in the extremely isolated Mojave Desert Amargosa River valley region of southeastern Inyo County, California, 105 km from Death Valley, approximately over the same time interval as the CDPH work in Death Valley (between 2011 and 2016). Similar rodent species were captured as in Death Valley and were tested for select hantaviruses using serology and RT-PCR to assess risk to human health and the conservation of the endemic endangered Amargosa vole.
Results:
Among 192 rodents tested, including 56 Peromyscus spp., only one seropositive harvest mouse (Reithrodontomys megalotis) was detected.
Discussion:
These data highlight the heterogeneity in the prevalence of hantavirus infection even among nearby desert communities and suggest that further studies of hantavirus persistence in desert environments are needed to more accurately inform the risks to public health and wildlife conservation.
Introduction
H
The Amargosa River valley region is an ecologically important understudied part of California, where increasing ecotourism in the towns of Shoshone and Tecopa Hot Springs in Inyo County potentially increases the risk for pathogen exchange (Pesapane et al. 2013) if SNV prevalence in reservoir species is demonstrated. In this region, north–south mountain ranges divide the Mojave Desert into arid alkaline valleys and desert pockets except where the Amargosa River surfaces creating wetland oases surrounded by alkali scrub and harsh desert playa. Amargosa marshes provide essential habitat for modest populations of humans, rodents, birds, amphibians, and reptiles. There are few permanent residents in Shoshone and Tecopa Hot Springs (31 and 150 people, respectively, with <10 people/mi2) (U.S. Census 2010). Buildings are sparse, but these man-made structures can act as refugia for peridomestic wildlife or offer additional resources favoring wildlife population growth, particularly in harsh environments, and subsequently increase the opportunity for human–wildlife contact (Kuenzi et al. 2001). Some species known to harbor hantaviruses are broadly distributed in the Amargosa River valley region, such as deer mice, cactus mice, and harvest mice, whereas others, such as the endangered Amargosa vole (M. californicus scirpensis), are highly restricted geographically (Cudworth and Koprowski 2010). In contrast to SNV, EMCV and ILV are not known to be pathogenic in humans, but because hantaviruses may also affect rodent fitness (Douglass et al. 2001, 2007, Kallio et al. 2007), the presence of these viruses in ecological communities could compound challenges facing endangered rodents such as the Amargosa vole.
EMCV and ILV are rarely assessed in rodent communities, but SNV is actively monitored by the Department of Public Health in California (CDPH) because of its importance to public health. SNV-reactive antibodies are common in rodent communities of the southwestern U.S. (Mills et al. 1997), and Inyo County had a >21% prevalence among P. maniculatus and >6% across all Peromyscus species from 2007 to 2016 (CDPH 2016), although prevalence varies across the heterogeneous landscape (Jay et al. 1997, Mills et al. 1997). Inyo is the second largest county in California and the ninth largest in the United States, including the lowest point of elevation in the United States (Badwater basin, 86 meters below sea level) and the highest (Mount Whitney, 4421 meters above sea level). From 2010 to 2011, CDPH detected a 40% prevalence of antibody reactive with SNV among cactus mice in Death Valley National Park (DVNP) and the species was confirmed to be SNV through nucleic acid sequencing (Burns et al. 2018). However, rodents in the Amargosa River valley have not been assessed for SNV even though this region is near Death Valley, is a target for ecotourism, and has SNV reservoirs. In this study, we report the results of serological and PCR assays for hantavirus from a neighboring rodent community that differ greatly from the findings in DVNP.
Materials and Methods
Rodents from marshes in Tecopa Hot Springs (3585202000N, 11681305700W) and Shoshone (3585802300N, 11681601600W), California were trapped (Fig. 1) using Sherman live traps (HB Sherman, Tallahassee, FL) baited with oats, peanut butter, and apples in grids as previously described (Klinger et al. 2015, Foley et al. 2016) unless marshes were particularly linear in which case the grid geometry was adjusted to reflect the geometry of the marsh. A subset of rodents captured from 11 marshes between December 2011 and May 2016, plus an additional nine carcasses (seven P. eremicus and two Neotoma lepida) from residences provided by community members in Shoshone were included in this study. These 11 marshes make up at least 56% of all vole-occupied habitat (Foley et al. 2016), are distributed among the northern, middle, and southern range, including isolated island patches, and represent both rural and residential-adjacent vegetation patches as well as areas that might be preferred by Peromyscus species. The number of trap nights for trapping conducted between December 2011 and March 2013 of this study was not available. Trap success for each species was, therefore, based on trapping conducted between November 2013 and May 2016 and calculated by multiplying the number of traps deployed per night by the number of nights in a sampling event for a total sum of 6207 trap nights during this period alone.
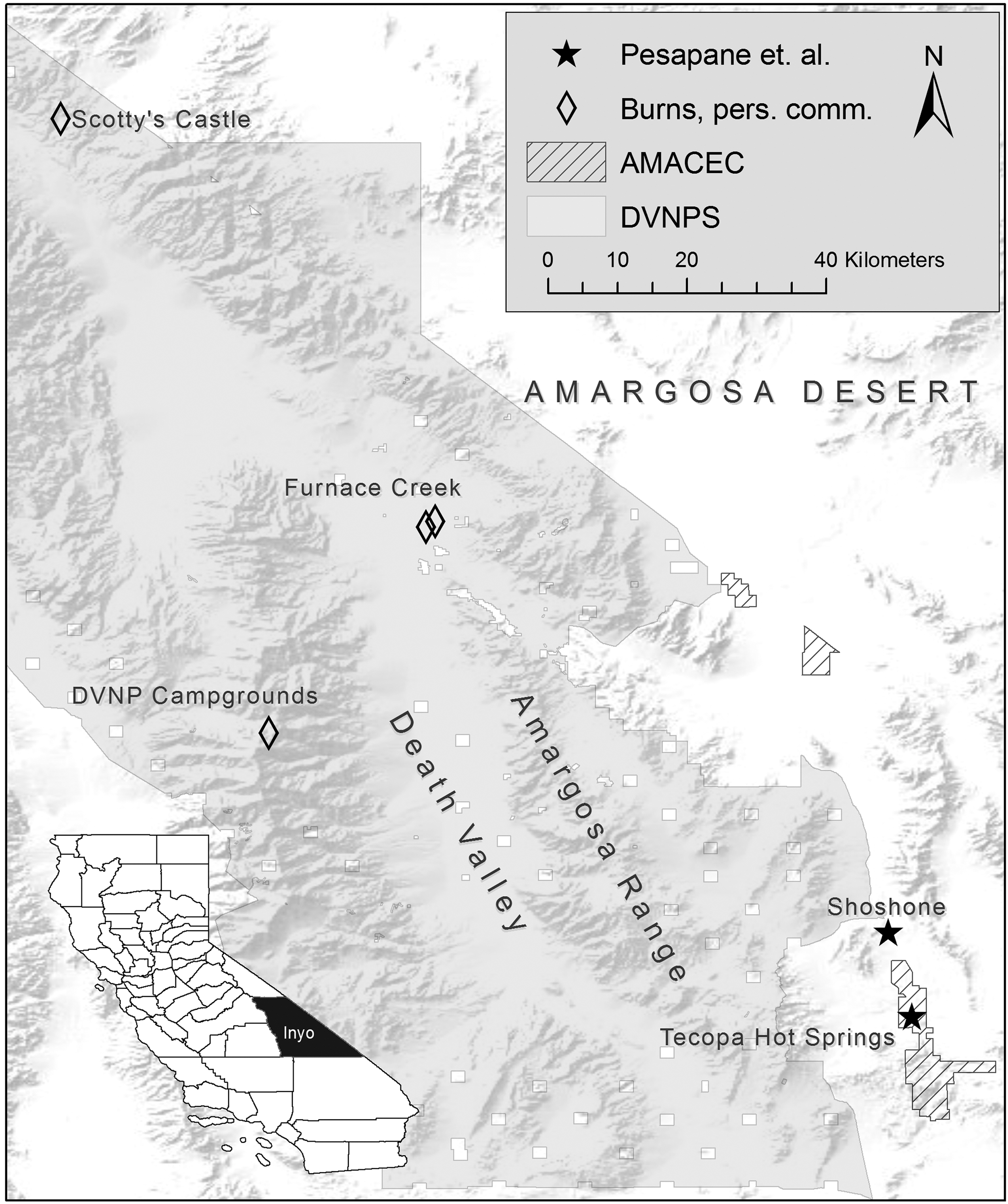
Hantavirus sampling sites in the Amargosa valley and Death Valley regions of southeast Inyo County, California. Rodents sampled in this study in the AMACEC and those sampled by Joe Burns at the California Department of Public Health in areas run by the DVNPS are shown in relation to topography in the Mojave Desert. An inset of Inyo County, California, is included for geographical reference. AMACEC, Amargosa Area of Critical Environmental Concern; DVNPS, Death Valley National Park Service.
Data were collected on the species, age, body mass, gender, and reproductive status of captured rodents. Each rodent was given a uniquely numbered ear tag (1005-1 Monel; National Band and Tag Co., Newport, KY) and only sampled once. For serology, blood samples were collected by retro-orbital bleeding using a heparinized capillary tube into BD Microtainer tubes with EDTA (Becton Dickson, Franklin Lakes, NJ). For PCR, rodents (except endangered voles) were euthanized using 100 mg/kg ketamine:10 mg/kg xylazine, and >100 mg/kg pentobarbital followed by cervical dislocation. Sampling was conducted with permission from the California Department of Fish and Wildlife, U.S. Fish and Wildlife Service, Bureau of Land Management, and UC Davis IACUC (agency permits TE54614A-1 and SC-000854; IACUC protocol 19905).
Blood samples were submitted to the CDPH Viral and Rickettsial Disease Laboratory (VRDL) for screening for IgG in response to SNV using an enzyme-linked immunosorbent assay (ELISA) (Ksiazek et al. 1995). This SNV ELISA reacts with antibodies against several New World hantaviruses (notably Sin Nombre, El Moro Canyon, and Isla Vista) (CDPH 2013).
Kidney tissue (25 mg) was extracted for PCR using the DNeasy Kit (Qiagen, Valencia, CA) following manufacturer's instructions. Immediately after extraction, RNA was reverse transcribed and amplified using the Titan One Tube RT-PCR System (Sigma-Aldrich, St. Louis, MI) with primers SNV-F (CAGCTGTGTCTGCATTGGAGAC) and SNV-R (TARAGYCCGATGGATTTCCAATCA) (Kramski et al. 2007), and then visualized on a 1% agarose gel. As a positive control, 9.10E+10 copies of a gBlock gene fragment (Integrated DNA Technologies, Skokie, IL) consisting of nucleotides 176–425 of the SNV nucleocapsid protein gene (GenBank ID: L25784.1) were used.
Prevalence and 95% confidence intervals were calculated with the function prop.test in R (v. 3.3.1, R Core Team 2016). We compared Amargosa data from 2011 to 2016 with data on SNV test results from neighboring DVNP from 2010 to 2013 provided by CDPH (Joseph Burns, pers. comm.) and with statewide data from 2010 to 2016 CDPH annual reports. The same serological diagnostic tests were performed by CDPH across all three data sets.
Results
In the Amargosa River valley from December 2011 to January 2014, 71 blood samples from deer mice, cactus mice, and Amargosa voles were antibody negative, whereas one harvest mouse was antibody positive (11% prevalence, 95% CI 0–49.3 among harvest mice) (Table 1). From May 2014 to May 2016, 121 kidney samples from five rodent species, including those from community residences, were PCR negative (Table 1).
SNV antigen in the serologic assay can cross-react with antibodies to several New World hantaviruses (notably EMCV and ILV). Data shown as number sampled and positives in parenthesis.
Data from this study. Serology completed by the California Department of Public Health, Viral and Rickettsial Disease Laboratory. RT-PCR conducted at UC Davis.
Data from trapping conducted by the California Department of Public Health Viral and Rickettsial Disease Laboratory in Death Valley National Park from 2010 to 2013 (Joseph Burns, pers. comm.), partially published in Burns et al. 2018.
Data summarized from California Department of Public Health Vector-Borne Disease Section Annual Reports 2010–2016.
Based on trapping conducted between November 2013 and May of 2016.
EMCV, El Moro Canyon orthohantavirus; ILV, Isla Vista virus; n, number of samples; pos, positive; prev., prevalence; SNV, Sin Nombre orthohantavirus.
Discussion
Serological assays for hantaviruses detect antibodies generated by hosts in response to exposure to a broad range of hantavirus species associated with human disease and, fortuitously, also detect species known only to occur in rodents. Based on rodent community composition at our study site in the Amargosa River valley, we could expect to detect SNV, EMCV, or the hantavirus species associated with Microtus previously known as ILV. Although antibodies reactive to SNV have been detected in harvest mice (Mills et al. 1997), the single SNV-antibody reactive harvest mouse in this study was presumably the result of cross-reactivity due to infection with EMCV for which harvest mice are the reservoir (Calisher et al. 2005), because spillover of SNV was unlikely given the consistently low numbers of deer mice sampled at these locations. Unfortunately, no additional sample was available from this individual for molecular analysis to conclusively determine the viral species.
None of our Amargosa voles were SNV-antibody reactive compared with 30.5% prevalence of SNV-antibody-reactive Microtus spp. throughout the state from 2010 to 2016 (Table 1). The absence of SNV-reactive antibodies in Amargosa voles suggests that exposure to endemic hantaviruses occurs rarely, if at all, in these voles, and the potential for deleterious effects associated with hantavirus infection in this endangered species is likely minimal.
House mouse blood from the Amargosa River valley had no detectable SNV antibodies in agreement with other California studies (Otteson et al. 1996, Bennett et al. 1999). Although Old World hantavirus-positive house mice have been described (Diglisic et al. 1994), neither the serological nor molecular assay used in our study would have detected exposure to Old World hantaviruses.
Although deer mice are documented reservoirs of SNV and the cactus mice population just across the Amargosa Mountains had 40% prevalence (Burns et al. 2018), our study found no evidence of SNV in Peromyscus spp. from the Amargosa River valley. Although CDPH annual reports indicate a 5.8% seroprevalence in cactus mice statewide from 2010 to 2016 (Table 1), that would include the 40 seropositive cactus mice reported in DVNP. Thus, we can conclude that the seroprevalence of cactus mice at all other locations in California was just 5/671 (0.7%) and that our results in the Amargosa reflect the overall limited number of SNV-antibody reactive cactus mice in the state. These findings lend support to the novelty of the DVNP location or the cactus mouse population at that site.
Although our limited sampling of the primary SNV reservoir, and potential secondary reservoir, yielded no evidence of SNV in the Amargosa River valley, endemic hantaviruses can disappear from rodent populations in isolated or fragmented habitats or even large patches when populations are sufficiently small (Calisher et al. 2005, Luis et al. 2015). The habitat patches in the Amargosa River valley are indeed ecological islands surrounded by harsh desert landscapes, which may be small enough that hantavirus cannot be maintained. Other factors that could account for the discrepancy, even with the presence of the same rodent species, between Amargosa where there is no SNV and DVNP where it is abundant, include population demographics and environmental attributes that influence food resources and virus persistence (Mills et al. 2010, Gorosito and Douglass 2017).
Conclusion
Our findings emphasize how landscape heterogeneity can drive variability in pathogen prevalence. Although there is no evidence of hantavirus in the towns of Shoshone and Tecopa Hot Springs at this time, it is not known whether infected rodents dispersing from the neighboring DVNP region could reintroduce hantavirus to the Amargosa River valley. Pathogen surveillance in wildlife is an important tool for the prevention of human morbidity and mortality related to zoonotic diseases. Our results highlight the importance of evaluating hantaviruses across heterogeneous landscapes as opposed to county lines. Because hantaviruses, specifically the pathogenic variant SNV, fluctuate with environmental and ecological interactions, longitudinal assessments with consistent sampling as discussed in Douglass and Vadell (2016) are also recommended to accurately reflect current threat of spillover to humans. Furthermore, considering studies that suggest there are fitness costs such as reduced reproduction for those that seroconvert (Douglass et al. 2001, 2007, Kallio et al. 2007), monitoring the prevalence of hantaviruses in threatened and endangered rodent populations is important for informing management decisions. Understanding not only local disease risk but also the regional context could contribute to an improved understanding of the interdependent human–animal–environmental health of Mojave Desert systems.
Footnotes
Acknowledgments
We thank Susan Sorrells from Shoshone Village, and Chris Otahal, Russ Scofield, and Brian Croft for logistical support. Amanda Poulsen and Caitlin Ott-Conn provided field and laboratory assistance. Hantavirus data sets from Inyo County were generously provided by Joseph Burns and details on serology methodology by Kristina Hsieh, from the Ontario and Richmond, CA offices of CDPH, respectively. Funding was provided by the Bureau of Land Management, the California Department of Fish and Wildlife, and the National Science Foundation Graduate Fellowship Program for R.R.P.
Author Disclosure Statement
No competing financial interests exist. The findings and conclusions in this report are those of the authors and do not necessarily represent the views of the California Department of Public Health.