
Abstract
This study investigated the prevalence of Staphylococcus aureus enterotoxin genes and shiga toxin -producing Escherichia coli (STEC) in fish and evaluated quality parameters of examined fish. A total of 150 fish samples belonging to 6 species (25/each species) were cultured on Baird–Parker agar and eosin methylene blue agar. Staphylococcal enterotoxin genes and virulence genes (stx1, stx2, and eaeA genes) in E. coli serotypes were determined by multiplex PCR. Aerobic plate count (APC), Enterobacteriaceae count, coliform count, and Pseudomonas count were performed. Also, levels of total volatile base nitrogen and histamine in fish were determined. The prevalence of S. aureus ranged from 4% to 36% and count from 2 to 4 log10CFU/g. The sed, sea, and seb genes in S. aureus isolates were detected with percentages of 40%, 26.6%, and 20%, respectively. The E. coli serotype O26 carried stx1, stx2, and eaeA. The APCs, Enterobacteriaceae counts, and Pseudomonas counts ranged from 5.1 to 7.2, from 2.01 to 3.9, and from 2.1 to 3.1 log10 CFU/g, respectively. The most probable number (MPN) of coliform ranged from 1.3 to 3.6 log10/g. Levels of total volatile basic nitrogen and histamine ranged from 29.2 to 12.2 and from 0.6 to 4.6 mg/100 g, respectively. Also, the value of thiobarbituric acid was highly significant (1.1 ± 0.084 mg MDA/kg) in Trachurus mediterraneus samples compared with those levels obtained from other fish species. Our findings concluded that those fish species could constitute a public health hazard as fish are reservoirs for enterotoxigenic S. aureus and Shiga toxin producing E. coli strains. This study highlighted the importance of screening of fish for enterotoxigenic S. aureus strains and STEC isolates, and also assessing the quality parameters of fish.
Introduction
F
Many outbreaks of food borne illness associated with seafood are derived from consumption of raw or inadequately heat-treated fish, which might be contaminated with different bacterial pathogens, including Salmonella spp., Escherichia coli, S. aureus, and Vibrio spp. from water sources, or the fish and fish products might be contaminated after the deferent processing stage (Pal et al. 2014). Humans could contract food poisoning through handling of infected fish or contact with contaminated water or other components of aquatic environment (Acha and Szyfres 2001, Gauthier 2015). The non- O157 Stx producing E. coli that produce only Stx2 are more often related to the hemolytic uremic syndrome (HUS) than those strains that produce only Stx1 or isolates that produce both Stx1 and Stx2 (Ethelberg et al. 2004).
S. aureus could grow in foods due to the wide range of growth parameters such as temperature from 7°C to 48.5°C with an optimum of 30°C to 37°C (Schmitt et al. 1990), pH from 4.2 to 9.3, with an optimum of 7 to 7.5 (Bergdoll 1983) and tolerate sodium chloride up to 15%. Staphylococcal food poisoning is attributed to staphylococcal enterotoxins (SEs) (Le Loir et al. 2003). The production of SEs occurs in the exponential phase of S. aureus growth, with the quantity dependent on the strain. Typically, SE induces food poisoning in humans when S. aureus is found at least 105–108 CFU/g in food (Seo and Bohach 2007).
The major etiology of fish spoilage is the activities of enzymes and microorganisms, resulting in unacceptability for human consumption. Microorganisms harbored in fish are usually associated with the fishing site, environmental factors, harvesting methods, the storage, and transportation. Also, the microbial growth during the storage will rely on the preservation condition (Ghaly et al. 2010).
The aim of current study was to determine the prevalence of enterotoxigenic S. aureus isolates and Shiga toxin producing E. coli (STEC) isolates from different fish species in Egypt, and also to evaluate the bacterial quality and freshness parameters of fish.
Materials and Methods
Collection and preparation of fish samples
One hundred fifty samples of Tilapia nilotica, Clarias lazera, Mugil cephalus, Synodus aurus, Pagrus pagrus, and Trachurus mediterraneus (25, each) were randomly collected from different fish markets at Zagazig city, Sharkia Province, Egypt, during May–October 2017. Twenty-five fish samples from each fish species were purchased from different retail markets on different days. The purchased fish samples were identified and separately packaged in sterile plastic ziplock bags (Mayank Plastics, Delhi, India), then kept in an insulated box with ice packs (4°C), and immediately transported to the Food Control Laboratory within 1 h. The scales and skin of each fish sample were aseptically removed by using sanitized scissor and scalpel. The scissor and scalpel were regularly sanitized between each fish sample by using ethyl alcohol 70%.
Afterward, 25 g of each fish sample was placed in a sterile Whirl-Pak® sample bag made of polyethylene (Thomas Scientific, Inc.) that contained 225 mL sterile buffered peptone water (Oxoid, CM509). Each bag was sealed and then followed by homogenization using a stomacher (Merck, Darmstadt, Germany) for 1 min. The homogenate was transferred into a sterile test tube. One milliliter was transferred into a sterile test tube containing 9 mL of 0.1% peptone water (Conda, Spain) and then followed by preparation of 10-fold serial dilution up to 107.
Enumeration, isolation, and identification of S. aureus
A quantity of 0.1 mL from the prepared serial dilution was inoculated on the dry surface of duplicate Baird–Parker agar (Oxoid CM 275) plates followed by incubation at 37°C for 24 h. The black-colored colonies surrounded by a wide clear area with opacity were recorded. Counting was repeated after reincubation at 37°C for further 24 h. S. aureus CFU/g was calculated as presumptive count (APHA 1992). The suspected colonies, revealing positive egg yolk reaction, were biochemically identified. Colonies demonstrating coagulase-positive, gram-positive cocci, and catalase-positive were biochemically identified in S. aureus.
Multiplex polymerase chain reaction for detection of enterotoxigenic S. aureus
Bacterial DNA was extracted by the Gene JET Genomic DNA Purification kit (# k0722; Ferments) following the manufacturer's protocols. The oligonucleotide sequences of se genes were chosen as previously described, as shown in Table 1, by Mehrotra et al. (2000). PCR conditions included an initial denaturation at 94°C/5 min; 35 cycles of denaturation at 94°C/2 min., annealing at 57°C/2 min, and extension at 72°C/1 min with a final extension at 72°C/7 min. Gel electrophoresis was carried out using 1.5% agarose gel in 0.5% Tris-acetate- EDTA (TEA) buffer stained with ethidium bromide. The amplified products were visualized on ultraviolet (UV) transilluminator.
Isolation, biochemical identification, and serotyping of E. coli
After homogenization of 25 g of fish samples in 225 mL buffered peptone water, samples were subjected for pre-enrichment at 37°C for 18 h. Afterward, 1 mL of the pre-enriched samples was directly inoculated into 9 mL of MacConkey broth (CM5a; Oxoid) and then followed by incubation at 37°C for 18 h (Cruickshank et al. 1975). A loopful from enriched broth was directly streaked on eosin methylene blue agar, EMB (CM69; Oxoid) plates and then subjected to incubation at 37°C for 18–24 h. The suspected E. coli colonies were subjected to biochemical identification (Barrow and Feltham 1993). The biochemically confirmed isolates were serotyped by using rapid diagnostic E. coli antisera sets (DIFCO Laboratories, Detroit Michigan 48232-7058) at the Food Analysis Center, Faculty of Veterinary Medicine, Benha University, as previously explained (Kok et al. 1996). E. coli O antisera consist of polyclonal antibodies, utilized for serotyping of E. coli O antigens.
Multiplex PCR for detection of Shiga toxin and intimin genes
The molecular identification of shiga toxins (stx1 and stx2) and intimin (eaeA) genes in E. coli isolates was performed by oligonucleotide primers (Pharmacia Biotech), as shown in Table 1. The oligonucleotide primers and program of PCRs were carried out as previously explained (Fagan et al. 1999). The program of PCRs included an initial denaturation at 95°C/3 min and then followed by 35 cycles at 95°C/20 s, 58°C/40 s, and 72°C/90 s. The final cycle was followed by a 72°C incubation/5 min. The amplicons were run on 1.5% of agarose gel electrophoresis stained with ethidium bromide. The PCR products were visualized on UV transilluminator.
Aerobic plate count
The aerobic plate counts (APCs) were detected by the surface spread technique according to APHA (1992). In brief, 0.1 mL of sample homogenate, from the selected dilutions, was inoculated on the duplicate sterile plates of prepoured and dried Standard Method Agar (Nissui Pharmaceutical Co., Ltd., Tokyo, Japan) followed by incubation of plates at 35°C for 48 h.
Enterobacteriaceae count
The Enterobacteriaceae counts (EBCs) were performed by pour plating method on violet red bile glucose agar. These plates were covered with a growth medium layer before their incubation at 37°C for 24 h (ICMSF 1986).
Coliform count
The coliform counts were determined according to Thatcher and Clark (1978). Positive tubes with acid and gas production were recorded for each dilution. The results were presented as a fraction as follows: number of positive tubes/number of inoculated tubes. The most probable number (MPN) was estimated using MPN index (USDA 2008) and the concentration of coliform bacteria was presented as MPN/g of the sample.
Pseudomonas count
Pseudomonas counts were carried out on the pseudomonas agar base (CM 559; Oxoid) supplemented with cetrimide, fucidin, and cephaloridine supplements (SR 103; Oxoid, Basingstoke, Hampshire, United Kingdom). Pseudomonas colonies were counted after incubation of plates at 25°C for 48 h.
Chemical parameters of fish freshness
Total volatile basic nitrogen (TVB-N) content of fish was detected as previously explained by Malle and Tao (1987). TVB-N content was expressed as milligram nitrogen/100 g fish. The four replicates were prepared for each tested fish sample. The thiobarbituric acid (TBA) assay was performed according to the procedure of Schmedes and Holmer (1989). Briefly, the fish sample (10 g) was mixed with 25 mL of 20% trichloroacetic acid (w/v) and homogenized in a blender for 30 s. After filtration, the filtrate (2 mL) was added to 2 mL of 0.02 M aqueous TBA in a test tube. Test tubes were subjected to incubation at room temperature in the dark for 20 h. The absorbance was measured by UV-vis spectrophotometer at 532 nm. The TBA value was expressed as milligram malonaldehyde per kilogram fish. ELISA was utilized to determine histamine by using RIDASCREEN® histamine kit (R-Bipharm AG, Germany) according to the manufacturer's recommendation. The limit of detection for fresh fish and canned fish is 2.5 ppm. Optical density was estimated at 450 nm on the ELISA plate reader. The complex amount bound to the plate and the optical density were inversely proportional to the concentration of histamine in the fish sample.
Statistical analysis
The data concerning bacterial counts, TVB-N, TBA, and histamine were subjected to analysis by one-way analysis of variance, followed by the post hoc Duncan's test to compare bacterial counts and values of chemical parameters between different fish species (IBM SPSS Statistics, version 22). The test was significant at p value <0.05. The data of APC, EBC, coliform count, and Pseudomonas count were expressed as average of log ± standard deviation (SD).While data of TVB-N, TBA, and histamine were reported as average of value mean ± SD.
Results
The prevalence of S. aureus was 8%, 4%, 16%, 24%, 36%, and 12% and counts ranged from 2 to 4 log CFU/g in examined T. nilotica, C. lazera, M. cephalus, S. saurus, P. pagrus, and T. mediterraneus, respectively (Table 2). Multiplex PCR was performed to detect enterotoxin genes of S. aureus from fish species. The highest prevalence belonged to sed gene (Fig. 1a). Also, the sed, sea, and seb genes of S. aureus isolates were detected with percentages of 40%, 26.6%, and 20%, respectively. Moreover, in two isolates, the sea plus sed (6.6%) and seb plus sed (6.6%) genes were detected simultaneously. In our study, se genes were found in 60% of recovered S. aureus isolates. The overall prevalence of E. coli was 15.33% (23/150). The descending prevalence of E. coli started with C. lazera (32%), M. cephalus (20%), T. nilotica (16%), P. pagrus (12%), S. aureus (8%), and T. mediterraneus (4%) (Table 3). The serotyping of E. coli isolates was distinguished into O125 (21.73%), O119 and O44 (17.4%, each), O26 and O128 (13%, each), and O111 serotype (8.6%). There were two untyped isolates of E. coli as illustrated in Table 3. The virulence genes associated with E. coli strains (stx1, stx2, and eaeA genes) from fish were illustrated (Fig. 1b). Multiplex PCR screening of isolates revealed that E. coli O26 carried stx1, stx2, and eaeA genes at molecular weights of 614, 719, and 890 bp, respectively. The E. coli serotype O111 was positive for stx1 and eaeA genes. The E. coli serotype O128 was positive for stx1 and E. coli O125 was positive for eaeA gene (Fig. 1b). While E. coli serotypes O44 and O119 were negative for those virulence genes. The untyped E. coli serotypes were not subjected to molecular detection of stx and eaeA genes.
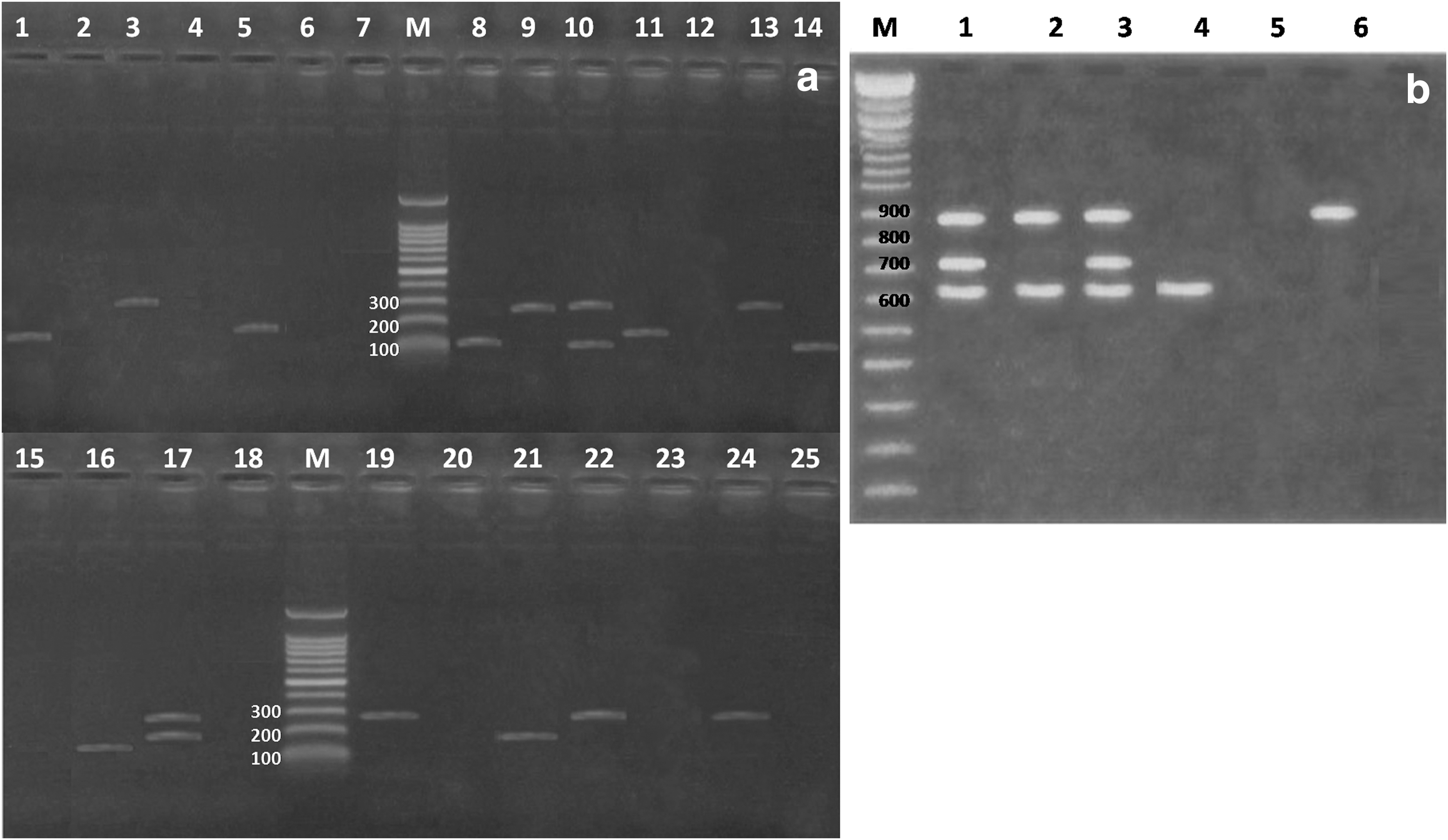
Agarose gel electrophoresis of multiplex polymerase chain reaction for detection of enterotoxin genes in Staphylococcus aureus and virulence genes in Escherichia coli strains isolated from fish.
APC mean values were 5.7 ± 2.1, 6.1 ± 2.22, 6.0 ± 1.98, 6.3 ± 2.44, 7.2 ± 2.96, and 5.1 ± 1.36 log10 CFU/g in the examined T. nilotica, C. lazera, M. cephalus, S. saurus, P. pagrus, and T. mediterraneus, respectively (Fig. 2a). The EBC mean values in the descending manner were 3.9 ± 1.42, 2.9 ± 0.91, 2.6 ± 0.93, 2.3 ± 0.96, 2.0 ± 0.97, and 2.0 ± 0.94 log10 CFU/g in the investigated C. lazera, M. cephalus, T. nilotica, P. pagrus, T. mediterraneus, and S. saurus, respectively (Fig. 2b). The MPN of coliforms in T. nilotica and C. lazera samples was 2.9 ± 1.04 log10 MPN/g and 3.6 ± 1.11 log10 MPN/g, respectively (Fig. 2c). However, M. cephalus harbored MPN of coliforms within an average of 2.8 ± 1.09 log10 MPN/g. The MPN of coliforms in the examined S. saurus, P. pagrus, and T. mediterraneus was 1.8 ± 0.77, 1.8 ± 0.84, and 1.3 ± 0.52 log10 MPN/g, respectively. In Figure 2d, Pseudomonas count mean values were 2.3 ± 0.98, 2.1 ± 0.69, 2.5 ± 0.94, 2.3 ± 1.12, 3.5 ± 1.05, and 3.1 ± 1.02 log10 CFU/g in T. nilotica, C. latera, M. cephalus, S. saurus, P. pagrus, and T. mediterraneus, respectively. In Table 4, there was a significant difference (p < 0.05) in TVB-N levels between P. pagrus compared with other fish species. In our study, the level of TBA showed a higher significance in T. mediterraneus fish samples compared with other species. There was a significant difference in the level of histamine in T. mediterraneus compared with other fish species (p < 0.05) (Table 4).
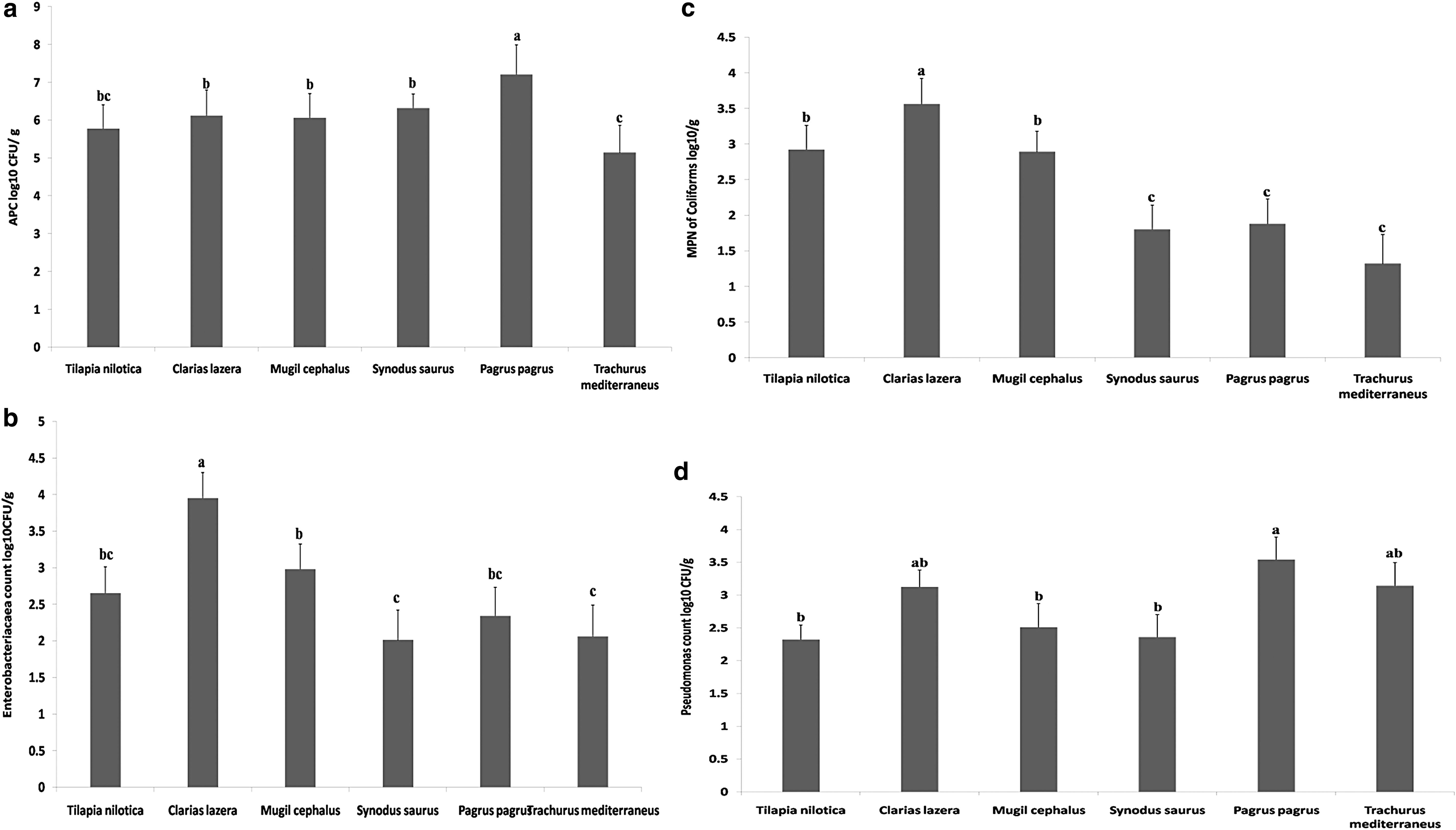
The rows carrying different letters showed a statistical significance (p < 0.05) according to the one-way analysis of variance.
TVB-N, total volatile basic nitrogen;
Discussion
Staphylococci are one of normal flora that inhabit humans and animals, because of their ubiquitous occurrence in the environment, and have a public health hazard. These pathogens were detected in various raw foods. Therefore, Staphylococcus enterotoxins (SE) are considered a worldwide problem (Vorster et al. 1994). Staphylococcal food poisoning usually arises due to the contamination of food with SE produced by S. aureus. Also, food handlers carry enterotoxin-producing S. aureus in their noses or on their hands, which are the main sources for food contamination (Argudín et al. 2010). The severity of symptoms of food poisoning is dependent on dose of ingested SE in food and health status of human. The older and younger ages of humans usually suffer from more complicated symptoms (FDA 2012). In our study, the prevalence of S. aureus was higher (36%) in P. pagrus, while it was lowest in T. nilotica (8%) and C. lazera (4%). Comparable prevalence was reported in Galicia (10-43%) in the examined fishery products (Vazquez-Sanchez et al. 2012). The counts of S. aureus ranged from 2 to 4 log CFU/g in examined six fish species. S. aureus concentration of 106CFU/g in food is required to produce enterotoxins at hazardous concentrations, possible when S. aureus grows at optimum conditions (3.5% NaCl and 37°C) as previously reported (ICMSF 1996). However, the growth of S. aureus may occur at 7°C, its toxin is not produced at temperatures below 10°C, and limited amounts of toxins are produced at a temperature ranging from 10°C to 20°C (Adams and Moss 2000).
A nearly similar count (3 log CFU/g) was found (Gonzalez-Rodriguez et al. 2001). Concerning the detection of enterotoxigenic S. aureus, the sed, sea, and seb genes of S. aureus isolates were recovered with percentages of 40%, 26.6%, and 20%, respectively. Also, in two isolates, the sea plus sed (6.6%) and seb plus sed (6.6%) genes were detected simultaneously. Our result indicated that existence of two enterotoxin genes in S. aureus isolates will result in more intoxication for humans. This was proved by the finding of Denayer et al. (2017), where the presence of multiple enterotoxin genes (sea, sed, seg, sei, sed, sej, and ser), in S. aureus isolates from human outbreak in Belgium, could have synergistic effects, and produces diarrhea and vomition, although a lower amount of enterotoxin (1.9 ng/100 mashed potatoes) was quantified. In the present study, the highest prevalence belonged to sed genes. Our results were consistent with the finding of Cha et al. (2006), where sea and sed genes were recovered from fish samples in Austria. In northwest Spain, Vazquez-Sanchez et al. (2012) found that sea gene was the most common enterotoxin gene in S. aureus isolates from fish and fish products. In this study, se genes were detected in 60% of examined S. aureus isolates. Conversely, a lower prevalence of S. aureus-se positive was cited in isolates from fishery products (Corrente et al. 2007, Oh et al. 2007).
The highest prevalence of E. coli was found in C. lazera (32%), followed by lowest percentage in T. mediterraneus (4%) and S. saurus (8%). The overall prevalence of E. coli was 15.33%. A nearly similar prevalence (13%) was reported in raw fish at Khartoum State fish market (Yagoub 2009). Otherwise, a lower prevalence of E. coli (5.9%) was recorded in Tilapia (Pao et al. 2008). A higher prevalence (48.9%) was found in India (Gupta et al. 2013). In our study, the highest prevalence of E. coli in C. lazera samples might be attributed to the habitat of this fish species, near to sediment. This explanation was augmented by Spinu et al. (2014), whereas a higher prevalence of E. coli in the sediment (70%), fish (80%), and water (30%). E. coli isolates were identified into different serotypes, including O125, O119, O44, O111, O128, and O26. Similarly, different serotypes of E. coli were isolated from fish sold in South Korea (Koo et al. 2012). Shiga toxin producing E. coli (STEC) isolates pose a zoonotic risk as they cause food poisoning outbreaks and HUS. The most common STEC serotype related with human illness is O157:H7, while there are more than 100 non-O157 serotypes that are implicated in causing HUS and hemorrhagic colitis (HC) (Coombes et al. 2011). In our study, E. coli O26 carries stx1, stx2, and eaeA virulence genes. Our data were concurred with the finding of Balière et al. (2015), who recovered one isolate of E. coli serotype O26:H11 bearing three genes (stx1, eaeA, and ehxA) from shellfish in France. However, E. coli serotypes O44 and O119 were negative for those virulence genes, While E. coli O111 was positive for stx1 and eaeA genes. E. coli O128 was only positive for stx1 and E. coli O125 was only positive for eaeA gene. In India, sxt1 and sxt2 genes were detected during the investigation of raw fish samples (Gupta et al. 2013). Previous studies found that Stx2 producing E. coli are strongly associated with HUS than those isolates producing Stx1 (Paton and Paton 2002).The production of Stx is correlated to pathogenesis of bloody diarrhea and HUS (Ethelberg et al. 2004). Moreover, Stx2 is the most common virulence factor related to severe human diseases (Pradel et al. 2008). The enterohemorrhagic E. coli (EHEC) belonged to the following serotypes: O26: H11, O103: H2, O111: H8, O118: H16, O121: H19, O145: H28, and O157: H7 were the most frequently isolated serotypes from food poisoning outbreaks, HC, and HUS cases (CDC 2009). There were scarce data on EHEC infection associated with the consumption of fish. In Japan, Asai et al. (1999) recovered an outbreak in eight patients infected with STEC O157: H7, and this outbreak could be attributed to ingestion of processed salmons infected with 84 isolates of E. coli O157: H7 carrying both stx1 and stx2 genes.
It was clear that the mean value of APC was highly significant in P. pagrus compared with other examined fish species. The International Commission on Microbiological Specification for Foods (ICMSF 1986) classifies fish into three categories according to total bacterial count, including good quality below 5 × 105, marginally accepted ranged from 5 × 105 to 107, and unacceptable fish more than 107. Most examined fish samples in this study were closely related to the marginally accepted group with the exception of P. pagrus fish samples that are related to the unacceptable group. While T. nilotica fish samples were mostly located in the good quality group. The significant value of APCs in P. pagrus compared with other fish species might be attributed to the lower marketability of this species in Zagazig fish markets, more exposure time to contaminated ice and utensils, and the temperature fluctuation in retail stores. The obtained results are within the range of previous studies in different countries. Pao et al. (2008) detected the mean counts for cat fish and tilapia at Virginia, were 7–6.5 log10 CFU/g and Gonzalez-Rodriguez et al. (2001) in Spain reported 5.27 log10 CFU/g in fish samples. These variations in APCs are attributed to the postharvest handling, packaging conditions, and storage temperature before and during the marketing process as previously explained by Ahmed et al. (2013). Enterobacteriaceae are implicated in fish spoilage during refrigerated storage (Sallam 2007). In this study, C. lazera samples showed a higher significance in the total Enterobacteriaceae count compared with other fish species (p < 0.05). This finding could be due to the feeding of C. lazera on nonprocessed chicken viscera. Nearly similar EBCs of 3 log CFU/g were detected in Spain (Gonzalez-Rodriguez et al. 2001). Our finding was in agreement with results reported for different fish species, including sea bass (Papadopoulou et al. 2007).
Higher MPN of coliforms was detected in C. lazera (3.6 ± 1.11 log10 MPN/g) and T. nilotica (2.9 ± 1.04 log10 MPN/g). Also, this study clarified that the MPN of coliforms in examined S. aurus, P. pagrus, and T. mediterraneus was 1.8 ± 0.77, 1.8 ± 0.84, and 1.3 ± 0.52 log10 MPN/g, respectively. The MPN of coliforms was nearly relevant to counts (Oku and Amakoromo 2013) in Nigeria during investigation of bacteriological status of fresh fish. Also, Begum et al. (2010) found that total coliform ranged from 1 to 2 log10 MPN/g. Similarly, Pao et al. (2008) detected 1.9 log10 MPN/g of marketed fish. The MPN of coliforms was significantly higher in fresh water examined species than marine water species (p < 0.05). The ability of coliform bacteria to survive in fresh water is responsible for higher density of these bacteria in fresh water fish. According to ICMSF (1986), the acceptable level for MPN of coliform is 102/g. Therefore, most fresh water fish species exceeded this level. Many microorganisms were grown on muscle and able to secrete wide varieties of hydrolytic enzymes, especially proteinases. Pseudomonas spp. is the main pathogen responsible for the deterioration process of food proteins (Pantazi et al. 2008). Pseudomonas growth causes limited shelf life of chilled fish under aerobic conditions (Ravi Sankar et al. 2008). The psychrotolerant gram-negative bacteria, including Pseudomonas spp., have the ability to grow on chilled fish causing spoilage (Gram and Huss 1996). In the present study, the mean values of Pseudomonas counts were 3.5 ± 1.05 and 3.1 ± 1.02 log10 CFU/g in examined P. pagrus and T. mediterraneus, respectively. A nearly similar count of 3.3 ± 0.27 log10 CFU/g was recovered in muscles of T. mediterraneus in Greece (Tzikas et al. 2007). Sallam (2007) detected 3.2 log10 CFU/g in salmon slices, Sapporo, Japan. However, a higher Pseudomonas count (4.80 × 107 CFU/g) was reported in Sarpunti fish in Bangladesh, and this higher finding could be attributed to poor handling, improper fish storage, and poor sanitary conditions during fish processing (Begum et al. 2010).
The quality changes of fish resulted from growth of spoilage microorganisms and biochemical reactions during fish storage. Many spoilage indicators are commonly utilized to determine the fish quality. TVB-N refers to the summation of ammonia, dimethylamine, trimethylamine, and the other basic volatile nitrogenous compounds. TVB-N content is superior to 10 mg/100 g in freshly caught fish, but does not exceed 15 mg/100 g. The TVB-N is an indicator of spoilage in fish species. Therefore, legal requirements have been established in different countries. When the TVB-N value is up to 25 mg/100 g, it indicates very good quality of fish. However, TVB-N value over 35 mg/100 g refers to spoilage (EEC 1995). In our study, there was a significant difference (p < 0.05) in TVB-N levels in P. pagrus compared with other examined fish species. This finding suggested that a higher total bacterial count could harbor many spoilage bacteria that are responsible for volatile base production particularly during fish storage at aerobic conditions. Most examined fish samples were categorized into the good quality according to Egyptian legislation 3495/2005, whereas it recommends that good-quality fish contains a level of TVB-N less than 30 mg/100 gm. TBA is widely utilized as an indicator to assess the degree of secondary lipid peroxidation (Jeon et al. 2002). Conversely, lower values of TBA were detected in trout fish (Ojagh et al. 2010). In our study, TBA value was significantly higher (1.0 ± 0.084 mg MDA/kg) in T. mediterraneus samples compared with other fish species. This might have accounted for the increase of oxidative deterioration in oily fish than other fishes. All examined fishes are regarded acceptable compared with other works, which suggested that 1–2 mg MDA/kg fish flesh is usually considered the limit, beyond that fish will develop the objectionable odor. The maximum level of TBA referring good quality of the fish is 5 mg of MDA/kg fish tissue, while the fish may be suitable for consumption up to 8 mg MDA/kg fish tissue (Sallam 2007).
A level of histamine is a guide for spoilage and the potential of the sample for inducing scombroid poisoning. During fish spoilage, particularly if the temperature is increased to above 10°C, histidine might be converted into histamine. Ingestion of human to ∼100 mg/100 g of fish flesh could induce histamine food poisoning (Edmunds and Eitenmiller 1975). The maximum permissible limit of histamine set by FDA is 50 mg/kg (FDA 2011). Also, in Egypt, the maximum permissible limit of histamine must not exceed 200 mg/kg for frozen, smoked, and salted fish, as previously listed by Egyptian Standard Specifications (ESS 2005a, 2005b, and 2005c). The European Food Safety Agency has reported a maximum daily intake of histamine (50 mg) in foods for healthy adult persons (EFSA 2011). Our study revealed that the histamine level ranged from 0.6 ± 0.06 mg/100 g in C. lazera to 4.6 ± 1.6 mg/100 g in T. mediterraneus. This finding is consistent with the international reference and maximum allowed limit of histamine (0.49–21.29 mg/kg) in fish and fish products, which is a marker for more freshness and shelf life of examined fish, safe for human consumption (Talab et al. 2017). However, higher levels of histamine were detected in some studies: Fletcher et al. (1988) reported 346.4–681.8 mg/kg in smoked fish in New Zealand; Laurent et al. (1995) found histamine level of 730–1200 mg/kg in yellow fin tuna at ambient temperature storage after 72 h in Melbourne; and Rahimi et al. (2012) detected a level of 17–2010 mg/100 g in canned tuna fish in Iran. In the current study, T. mediterraneus showed higher significance of histamine level (4.6 ± 1.65 mg/100 g) compared with other fish species (p < 0.05). These variations might have resulted from a higher amount of amino acid histidine, which is a precursor of histamine under bacterial carboxylation effect. This result fully coincided with the finding of Kim et al. (2001), whereas mackerel and tuna fish had larger quantities of histidine. During the spoilage, histidine is converted to histamine, particularly if the temperature of storage rises above 8°C.
Conclusion
This study confirmed that the examined fish are reservoir for enterotoxigenic S. aureus (carrying sea, seb, and sed genes) and for STEC isolates (bearing stx1, stx2, and eaeA genes), constituting a zoonotic potential for consumers and fish handlers. Also, a higher Enterobacteriaceae count was detected in C. lazera due to feeding on viscera of nonprocessed chicken, while higher MPN of coliform was found in P. pagrus. The maximum level of TBA value is an indicator for good quality of the fish. Also, detection of histamine level in examined fish within the permissible limit indicated fish freshness and safety for consumers. Therefore, this study highlighted the necessity for continuous fish screening for enterotoxigenic S. aureus and virulence genes associated with E. coli serotypes, besides the assessment of quality and freshness parameters of fish to prevent food poisoning outbreaks in humans and fish spoilage.
Footnotes
Author Disclosure Statement
No conflicting financial interests exist.