
Abstract
Leishmania mexicana is an intracellular parasite that causes cutaneous leishmaniasis (CL) in some countries, including Mexico. Recently, we identified the elongation factor-1α (EF-1α) of L. mexicana by immunoproteomic analysis. In Leishmania donovani, this molecule has been reported as a virulence factor involved in downregulation of macrophages by no-canonical function when EF-1α interacts with protein tyrosine phosphatase-1 (SHP-1). However, in L. mexicana the key role of EF-1α in host–parasite relationship has not been elucidated, by this reason we started with cloning and recombinant expression of this antigen. A sequence of 1350 bp corresponding to EF-1α (EF-Lm) full-length gene was amplified from genomic DNA of L. mexicana (GenBank: MG256973); this gene contains only one nucleotide change: C464T, compared with L. mexicana reference sequence (GenBank: FR799570.1). The gene cloned (EF-Lm) codes for a protein of 449 residues. It was expressed in Escherichia coli and purified as 63 kDa sumo-fusion protein, which was recognized in the sera of patients diagnosed with CL. Our results show that EF-Lm is immunogenic during infection, and the rEF-Lm could be used for further analyses in the host–parasite relationship.
Introduction
B
To start a more detailed analysis of EF-1α in host–parasite relationship evoked in CL, our aim was to clone and express a recombinant of EF-1α from L. mexicana.
Materials and Methods
To clone the EF-1α gene, located on the chromosome 17 of the genomic sequence of L. mexicana strain MHOM/GT/2001/U1103 (GenBank: FR799570.1), optimal primers designated as Z-F (5′-TGGGCAAGGATAAGGTG-3′) and Z-R (5′-TTACTTCTTCGCAGCCTT-3′) were designed in silico using Oligo 7® software, and analyzed in the Workbench, Gene DB, and NCBI databases, afterward were synthesized (Sigma-Aldrich Mexico). Genomic DNA L. mexicana was isolated using the PureLink Genomic DNA kit (Qiagen), and was visualized on agarose gel 1% w/v with SybrGold (Invitrogen) staining on UV transilluminator.
Each PCR-mix (all reactives from Invitrogen) microtube contained 1 U of Taq polymerase, 1 μL of 2 mM dNTPs, 1.5 μL of 2 mM MgCl2 and 40 mM of each forward and reverse primers, 5 μL of Leishmania DNA or not related DNA (as negative control) and nuclease-free water with a final volume of 25 μL. The PCR was performed using a Mastercycler Epgradient (Eppendorf), programmed as follows: an initial DNA denaturing step at 95°C for 5 min; 25 cycles of 95°C for 2 min, primers annealing at 55°C for 2 min and 72°C for 2 min with a final extension period at 72°C for 5 min, and the products were resolved on 1% agarose gel as described above. Specific amplicons were excised with sterile scalpels and purified using a Qiaquick Gel kit (Qiagen).
DNA sequencing was performed at LANGEBIO (Laboratorio Nacional de Genómica para la Biodiversidad in Irapuato, Guanajuato, México) and confirmed by sequence obtained using Macrogen (South Korea).
L. mexicana EF-1α full-length gene (EF-Lm) was inserted into pETSUMO expression system (Life Technologies) using T4 ligase. The recombinant DNA obtained was amplified by growing the transformed Escherichia coli Mach-1 in LB broth medium supplemented with 25 μg/mL of kanamycin, and then was purified using NucleoSpin® Plasmid column (MN); the purified DNA was used for transformation of E. coli BL21 cells. The recombinant expression of EF-1α as sumo-fusion protein containing His6-tag was induced by 1 mM of IPTG until the optic density was 0.6 at 600 nm. Recombinant EF-Lm (rEF-Lm) was partially purified from inclusion bodies, solubilized in 5 M-guanidine hydrochloride, and then purified by affinity chromatography using a HiTrap chelating column (GE), following the protocol described previously by our group (López-Moreno et al. 2003).
Purified protein was separated by conventional 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) following standard protocols. The antigenicity of the protein was evaluated by Western blot using CL sera of patients infected with L. mexicana (confirmed by PCR-RFLP) (Pérez-Vega et al. 2009, Ochoa-Díaz et al. 2012), diluted 1:100, followed by incubation of the samples with antihuman IgG peroxidase conjugated (Bio-Rad) at 1:3000 dilution. The immunoreaction was evidenced by H2O2 (JT Baker) and diaminobenzidine (Research Organics) as previously described (Salazar et al. 2010).
Results
Since the immunodominant antigen p29 was identified as EF-1α of L. mexicana (Salazar et al. 2010, Piedra-Quintero et al. 2015), we focused on obtaining the nucleotide sequence of the EF-1α gene. As predicted in silico, a 1350-bp amplicon was obtained (Fig. 1A). As expected, a nucleotide sequence of 1350 bp was obtained and was 99.9% identical to that of the EF-1α from L. mexicana strain MHOM/GT/2001/U1103; they only differed in one nucleotide: T to C, this sequence was deposited in GenBank: MG256973 (Fig. 1B).
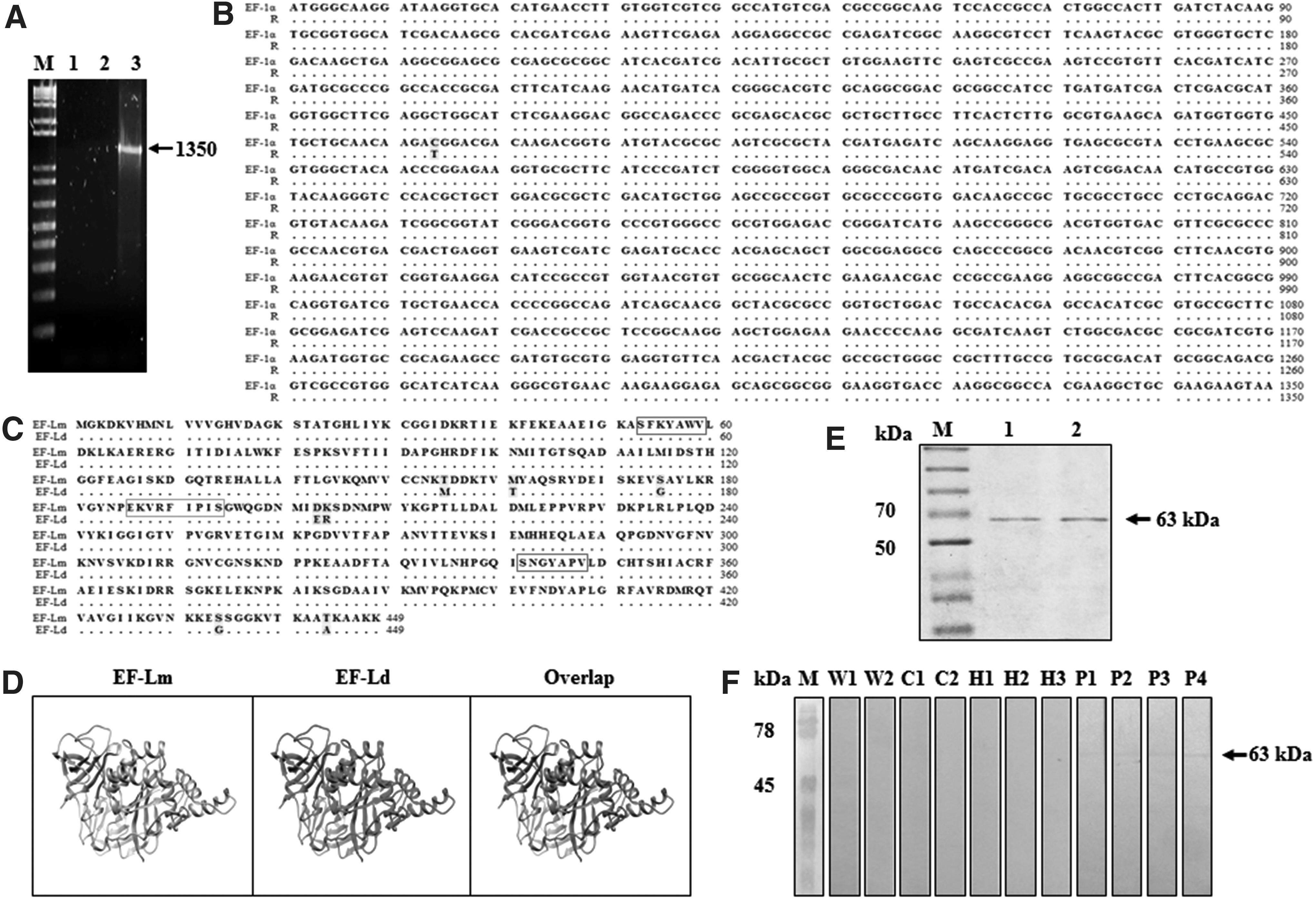
Cloning and recombinant expression of elongation factor-1α of Leishmania mexicana.
In the BLAST analysis of the amino acid sequence of the EF-Lm protein of the strain MHOM/MX/92/UAY68 with the EF-Ld protein (GenBank: AAL08019), a 98.4% similarity was observed when differing in 7 amino acids of the 449 that make up the complete protein (Fig. 1C). In the amino acid sequence of EF-Lm, an exposed region was identified (EKVRFIPIS, 186
Likewise, the amino acid sequence analysis of EF-Lm revealed the presence of two immunoreceptor tyrosine-based inhibitory motifs, which are also conserved (xxxYAxV) (shown in red boxes, Fig. 1C), since in other studies focused on the structural analysis of EF-1α of L. donovani these conserved regions had been identified by Nandan et al. (2003). These motifs are potential binding sites to SH2-domain-containing proteins, which explains the ability of Leishmania EF-1α to bind SHP-1 and perform its noncanonical function. In addition, the prediction of the tertiary structure of the EF-Lm and EF-Ld proteins was performed, obtaining a 98.8% identity in the structural comparison between both proteins; however, this difference in a single nucleotide does not generate structural modification of the protein (Fig. 1D) (Kelley et al. 2015).
The gene was inserted into pETSUMO expression system, and expressed as recombinant sumo-fusion protein that was purified and evidenced as 63-kDa band, which includes the 50 kDa of EF-1α plus 13 kDa of sumo-protein (Fig. 1E). Likewise, the antigenicity of rEF-Lm was confirmed by Western blot, where a 63-kDa reactive band was observed, showing that rEF-Lm presents antigenicity reacting with the sera of patients with parasitological diagnosis of CL (Fig. 1F).
Discussion
The production of rEF-Lm gives the possibility of increasing our knowledge about this molecule, including a more detailed study of the interaction with SHP-1 in macrophages or dendritic cells, contributing to elucidate the immunological mechanisms involved in host–parasite relationship of CL caused by L. mexicana as was reported in VL by L. donovani (Nandan et al. 2003). The antigenicity of rEF-Lm evidenced in this study was similar to that reported for other parasites, such as Giardia duodenalis, where EF-1α has been identified as an immunoreactive protein during acute infection (Skarin et al. 2011).
In addition, the immunogenicity properties of rEF-Lm suggest that it can be used in a more extensive study of host–parasite relationship, or it can be evaluated as a candidate for designing a diagnosis method of CL, as proposed for other recombinant proteins such as rLdPDI and rKLO8 from L. donovani (Kushawaha et al. 2012, Abass et al. 2013). However, the possible cross-reactivity with other related parasites such Trypanosoma cruzi should be evaluated. Finally, rEF-Lm can also be evaluated as a vaccine candidate against CL or other Leishmaniasis variants.
Conclusion
This study showed for the first time the cloning of L. mexicana EF-1α full-length gene, as well as its recombinant expression and purification. The purified recombinant protein (rEF-Lm) presents immunoreactivity against sera of CL patients, showing that it can be used in further studies that involve EF-1α and supports further research in host–parasite interaction.
Footnotes
Acknowledgment
The authors especially acknowledge Zayda Piedra, MS, for technical laboratory assistance.
Author Disclosure Statement
No conflicting financial interests exist.