
Abstract
Historically, Bartonella spp. have been associated with febrile illness (Oroya fever, trench fever, and cat scratch disease), endocarditis (numerous Bartonella spp.), and vasoproliferative lesions (Bartonella bacilliformis, Bartonella quintana, Bartonella henselae, and Bartonella vinsonii subsp. berkhoffii), occurring most often but not exclusively in immunocompromised patients. Recently, bloodstream infections with various Bartonella spp. have been documented in nonimmunocompromised individuals in association with a spectrum of cardiovascular, neurologic, and rheumatologic symptoms. As documented in this family, symptoms for which the medical implications remain unclear can occur in multiple family members infected with one or more Bartonella spp. Serial serologic and molecular microbiological findings supported exposure to or infection with Bartonella spp. in all seven family members. Either antibiotics failed to eliminate bacteremic infection, resulted in partial resolution of symptoms, or potentially reinfection occurred during the 19-month study period. There is a substantial need for clinical research to clarify the extent to which Bartonella spp. bacteremia induces nonspecific cardiovascular, neurologic, or rheumatologic symptoms, for ongoing improvement in the sensitivity and specificity of diagnostic testing, and clarification as to if, when, and how to treat patients with documented Bartonella spp. bacteremia.
Introduction
T
In this study, individual case histories are reported for a family residing in Quebec, Canada, in conjunction with Bartonella spp. serology and molecular microbiological findings.
Materials and Methods
Epidemiological investigation
The family consisted of a father, mother, maternal grandmother, two daughters, and two sons, residing in a small rural town surrounded by farms and forest in the Lanaudière region, Quebec, Canada. Historically, with the exception of the eldest daughter and mother, no unusual symptoms or illnesses were reported in remaining family members before August 2013. The family had lived in the current home surrounded by fields and forest since 2006. With the exception of the father, who had visited Maine, no other family member had traveled outside of Quebec before illness onset. Occasionally, ticks, fleas, biting flies, spiders, ladybugs, Larder beetles, book lice, mosquitoes, and mice were found around or inside the house. Bats, deer, fish, foxes, frogs, mice, rabbits, squirrels, and toads were frequently observed in the peridomestic environment.
At the onset of illness in the youngest daughter (August 2013), the family pets consisted of a domestic rabbit, a pet rat, cat, and a dog. All pets had outdoor and indoor access and would sleep with the children. All family members reported cat, dog, and rabbit bites or scratches for months to years before illness onset. The pet rabbit died in February 2014 after being infested with ticks found within the house. A tick, identified by the regional public health laboratory as Ixodes cookei (a tick species often found on rabbits and other small mammals), was subsequently removed from the family cat during the summer of 2014. Due to ongoing illness problems, the cat (9-year-old castrated male domestic shorthair) and dog (8-year-old castrated male Labrador retriever) were euthanized in February 2015. At necropsy, the cat had lymphoplasmacytic inflammation involving the kidney, liver, pancreas, and synovium, whereas the dog had a cutaneous mast cell tumor and lymphoplasmacytic multifocal chronic arthritis, consistent with historical lameness that began in October 2014.
Between June 2014 and August 2016, family members were tested for Bartonella henselae antibodies at the CHU Sainte-Justine Laboratory using an indirect immunofluorescence antibody (IFA) assay and commercially available reagents (Focus Laboratories, DiaSorin Molecular LLC, Cypress, CA). Individual serology results for each family member are reported in conjunction with the medical histories in Supplementary Data. Although the youngest daughter was the first of the four children to develop lymphadenopathy (August 2013), it was not until the youngest son was hospitalized in June 2014 with axillary lymphadenopathy that Bartonella serology was first obtained for any family member. B. henselae seroconversion was documented in the youngest son (Supplementary Data). During the first 6 months of 2015, B. henselae IFA titers (1:640–1:1280) were reported for the remaining five family members. Given Bartonella seroreactivity in all family members, the mother contacted the corresponding author on April 21, 2015, requesting to enroll all family members in a vector-borne disease research study (Bartonella spp. infection in high-risk individuals with arthropod exposure and animal contact, North Carolina State University Institutional Review Board approval 1960-11) With the exception of September 2015, when the family traveled to North Carolina to facilitate blood collection (second research testing time point), all other blood specimens were collected in Canada and shipped to North Carolina.
Overall family health considerations
Detailed medical histories for each family member are reported in Supplementary Data. Selected symptoms for each individual are summarized in Table 1. The youngest daughter and youngest son experienced recurring lymphadenopathy (Fig. 1). The youngest son also experienced auricular swelling and hyperemia (Fig. 2). Beginning in 2013, the eldest son and both daughters experienced periodic epistaxis, with the youngest daughter experiencing nasal bleeding events multiple times a week. Starting in early 2014, dental disease, lingual papillitis (more severe in the children), generalized itching, painful pimple-like eruptions in the scalp, and persistent sore throats were symptoms ultimately shared by all family members. During 2015, the entire family was treated for pinworms with brexpiprazole. All family members reported intense “needle-like” pain that was acute at onset, short in duration, and resolved spontaneously. The mother, youngest daughter, and eldest son developed wrinkles involving the skin overlying the feet. Despite repeated testing, there were generally no hematological or biochemical abnormalities among family members. Neutropenia, thrombocytosis, and echinocytes were occasionally reported. Erythrocyte sedimentation rate and C-reactive protein were infrequently increased. Hyperbilirubinemia occurred without concurrent increases in alanine aminotransferase and aspartate aminotransferase activities. Through November 2018, the health of all family members has remained relatively stable. The mother uses essential oils, herbs, muscle massage for pain relief, and encourages dietary consumption of nuts, fruits, and vegetables for all family members. In addition, she encourages relaxation time and exercise for the children.
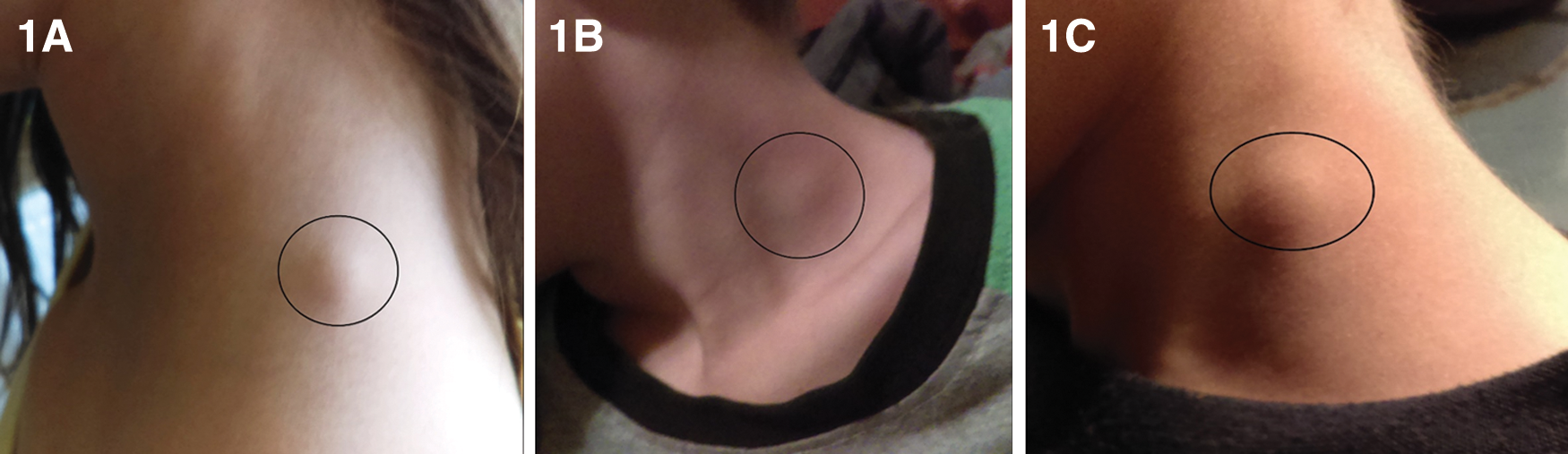
Swelling of the cervical lymph nodes in the youngest daughter, August 2016
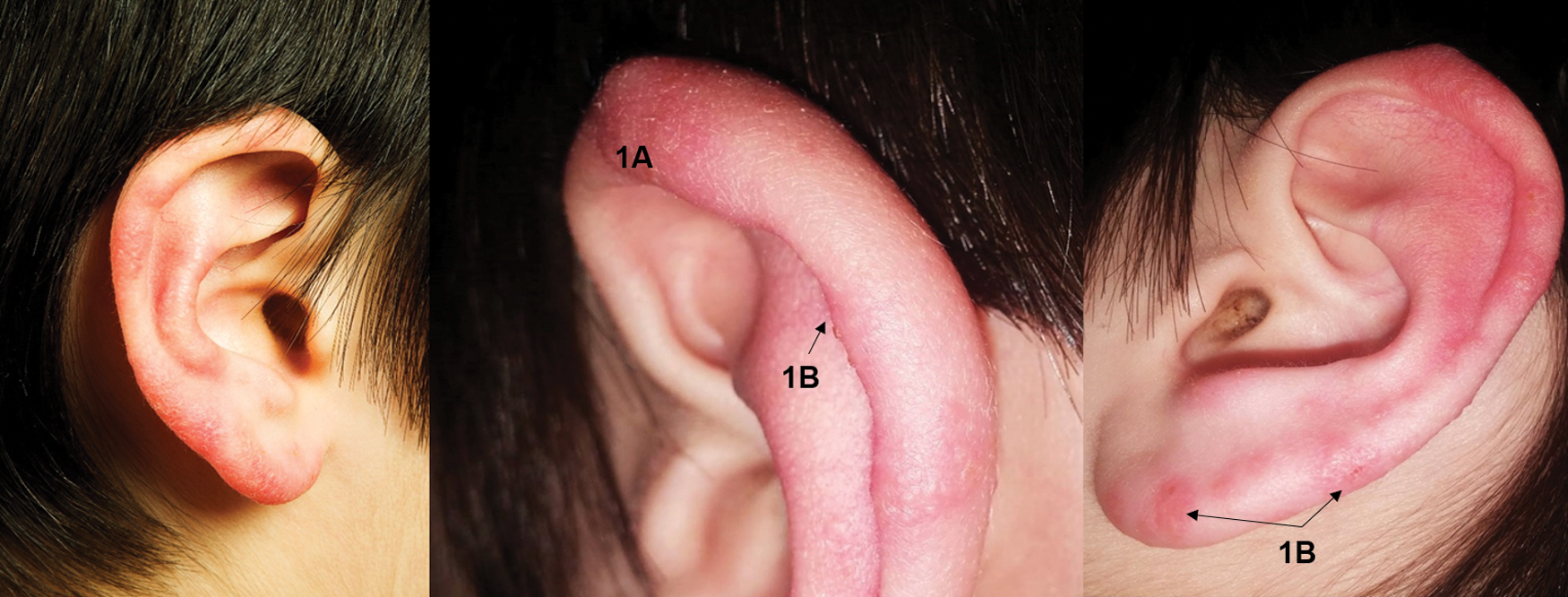
Auricular hyperemia with swelling
Vector-borne disease research testing methods
During 2015, family members were tested in May (first blood samples collected and shipped from Canada, overnight express, to the North Carolina State University, College of Veterinary Medicine, Intracellular Pathogens Research Laboratory [NCSU-CVM-IPRL]) and again in September, when the family traveled to Raleigh, North Carolina, for sample collection at a local hospital. Subsequently, blood and serum specimens were collected in Canada and shipped to the NCSU-CVM-IPRL. At each testing occasion, whole blood and serum sample sets were obtained on Monday, Wednesday, and Friday (three sample sets) for Bartonella sp. serology and inoculation of whole blood into Bartonella alpha-Proteobacteria growth medium (BAPGM), as previously described (Duncan et al. 2007, Breitschwerdt et al. 2010b, Mascarelli et al. 2011, 2013, Lantos et al. 2015, Oteo et al. 2017).
PCR for Bartonella internal transcribed spacer (ITS) region detection
Specifically, Bartonella spp. PCR was performed following DNA extraction from whole blood and serum; PCR following BAPGM enrichment blood culture incubated for 7, 14, and 21 days; and PCR was performed from subculture isolates if they were obtained after subinoculation from the BAPGM flask onto plates containing trypticase soy agar with 10% sheep whole blood, incubated for 4 weeks. PCR specimen preparation, DNA extraction, and PCR amplification were performed in three separate rooms with unidirectional workflow to avoid DNA contamination. In addition, BAPGM cultures were processed in a biosafety cabinet with HEPA filtration in a limited access biosafety level III laboratory.
Conventional PCR analysis
Conventional PCR (cPCR) screening of Bartonella internal transcribed spacer (ITS) region was performed using oligonucleotides 325 s: 5′-CTT CAG ATG ATG ATC CCA AGC CTT CTG GCG-3′ and 1100as: 5′-GAA CCG ACG ACC CCC TGC TTG CAA AGC A-3′ as forward and reverse primers, respectively. Amplification was performed in a 25 μL final volume reaction containing 12.5 μL of MyTaq Premix (Bioline), 0.2 μL of 100 μM of each forward and reverse primer (IDT® DNA Technology), 7.5 μL of molecular-grade water, and 5 μL of DNA from each sample tested. PCR-negative controls were prepared using 5 μL of DNA from blood of a healthy dog. Positive controls for PCR were prepared by using 5 μL of DNA from a serial dilution (using dog blood DNA) of B. henselae genomic DNA at a concentration equivalent to 0.001 pg/μL. cPCR was performed in an Eppendorf Mastercycler EPgradient® under the following conditions: a single hot-start cycle at 95°C for 2 min followed by 55 cycles of denaturing at 94°C for 15 s, annealing at 68°C for 15 s, and extension at 72°C for 18 s. Amplification was completed by an additional cycle at 72°C for 1 min, and products were analyzed by 2% agarose gel electrophoresis with detection using ethidium bromide under ultraviolet light. Amplicon products were sequenced to establish species strain identification.
Quantitative PCR analysis
Quantitative PCR (qPCR) screening of Bartonella ITS region was performed using oligonucleotides 325 s: 5′-CTTCAGATGATGATCCCAAGCCTTCTGGCG-3′ and 543as: 5′-AATTGGTGGGCCTGGGAGGACTTG-3′ as forward and reverse primers, respectively, and oligonucleotide BsppITS438probe: 5′ FAM-GGTTTTCCGGTTTATCCCGGAGGGC-BHQ1 3′ as TaqMan probe. Amplification was performed in a 25 μL final volume reaction containing 12.5 μL of SsoAdvanced™ Universal Probes Supermix (Bio-Rad), 0.2 μL of 100 μM of each forward primer, reverse primer, and TaqMan probe (IDT® DNA Technology), 7.5 μL of molecular-grade water, and 5 μL of DNA from each sample tested. PCR-negative controls were prepared using 5 μL of DNA from blood of a healthy dog. Positive controls for PCR were prepared by using 5 μL of DNA from a serial dilution (using dog blood DNA) of B. henselae genomic DNA equivalent to 0.1, 0.01, and 0.001 pg/μL. qPCR was performed in an CFX96® (Bio-Rad) under the following conditions: a single hot-start cycle at 95°C for 3 min followed by 45 cycles of denaturing at 94°C for 10 s, annealing at 68°C for 10 s, and extension at 72°C for 10 s. Amplification was completed by an additional cycle at 72°C for 30 s. Positive amplification was assessed by analysis of detectable fluorescence versus cycle values. Positive amplicon products were sent for sequencing to establish species and strain identification.
In no instance was B. henselae or DNA of any other Bartonella spp. amplified in the negative control lane on any PCR gel. To assess for potential contamination during blood sample processing into liquid BAPGM flasks, an uninoculated BAPGM culture flask was processed simultaneously and in an identical manner with each batch of patient blood and serum samples tested. For all components of the BAPGM platform (PCR from blood, serum, enrichment cultures at 7, 14, and 21 days), PCR-negative controls remained negative throughout the course of the study (Varanat et al. 2009). In addition, subcultures of uninoculated BAPGM (culture control) at 7, 14, and 21 days did not yield bacterial growth. Using previously described primers and assay conditions (Maggi et al. 2013a, Breitschwerdt et al. 2014), PCR was performed targeting Anaplasma, Babesia, Ehrlichia, and hemotropic Mycoplasma spp.
Serological analyses
As previously described (Maggi et al. 2011, 2012, 2013, Mascarelli et al. 2011), Bartonella vinsonii subsp. berkhoffii genotypes I, II, III, B. henselae (SA2 strain), and Bartonella koehlerae and B. quintana antibodies were determined by indirect immunofluorescence antibody assay (IFA) using fluorescein-conjugated goat anti-human IgG (Pierce Biotechnology, Rockford, IL). Briefly, Bartonella organisms of feline (B. koehlerae [NCSU 09FO-01, Trillium] and B. henselae SA2 [NCSU 95FO-099]), canine (B. vinsonii subsp. berkhoffii genotype I [NCSU 93CO-01 T'weed], II [NCSU 95CO-08, Winnie], and III [NCSU 06CO-01 Klara]), and monkey (B. quintana [NCSU 11MO-01]) origin were passed from 5% sheep blood agar into cell lines to obtain antigens for IFA testing as described previously (Hegarty et al. 2014). Heavily infected cell cultures were spotted onto 30-well Teflon-coated slides (Cel-Line/Thermo Scientific, Waltham, MA), air-dried, acetone fixed, and stored frozen until tested. Serum samples were diluted in phosphate-buffered saline (pH 7.4) containing normal goat serum, Tween-20, and 0.5% of powdered nonfat dry milk to block nonspecific antigen binding sites. Sera were tested at dilutions of 1:16–1:8192. To avoid confusion with possible nonspecific binding found at low dilutions and to standardize with other laboratories such as the Centers for Disease Control and Prevention, a cutoff titer of ≥1:64 was used to define a seroreactive titer.
Research Testing Results
Bartonella spp. serology results varied among individual family members and individual results varied during the study (Table 2). Overall, the most consistent IFA seroreactivity was to B. vinsonii subsp. berkhoffii genotype II and B. henselae, less seroreactivity to B. vinsonii subsp. berkhoffii genotype III and B. koehlerae, and minimal seroreactivity to B. vinsonii subsp. berkhoffii genotype I or to Bartonella quintana antigens. The youngest daughter was initially nonseroreactive, but subsequently developed low antibody reactivity to B. vinsonii subsp. berkhoffii genotype II and B. henselae. The eldest son tended to become seroreactive to an increasing number of Bartonella spp. over time, ultimately becoming seroreactive to all six Bartonella spp. IFA test antigens in December 2016. Similarly, the eldest daughter was initially nonseroreactive, but subsequently became seroreactive to multiple Bartonella sp. antigens. In contrast, the youngest son had a progressive decrease in seroreactivity to the IFA antigen panel over time and was nonseroreactive between March and December 2016. The mother and grandmother were reactive to nearly all six antigens at all testing time points, whereas the father was nonseroreactive with the exception of one antigen at two time points.
Sequential Bartonella spp. Serology and PCR Results from Blood, Serum, and Bartonella Alpha Proteobacteria Growth Medium Enrichment Blood Cultures for Each of the Seven Canadian Family Members
Bold-faced reciprocal titers are considered seroreactive, reflecting support for exposure to a Bartonella spp. Bartonella serology results reported in the individual case histories (Supplementary Data) were generated at the CHU Sainte-Justine Laboratory, Quebec, Canada, before beginning the research testing from the Intracellular Pathogens Research Laboratory at North Carolina State University reported in this table. As per the Methods section, BAPGM enrichment cultures were tested by PCR at 7, 14, and 21 days after blood inoculation. The day that the culture was PCR positive is reported.
Bartonella conventional or qPCR product successfully sequenced.
BAPGM, Bartonella alpha Proteobacteria growth medium; Bvb, Bartonella vinsonii subsp. berkhoffii; Bh, Bartonella henselae; Bk, Bartonella koehlerae; Bq, Bartonella quintana; cPCR, conventional PCR; NT, not tested as culture was discarded due to obvious contamination; qPCR, quantitative PCR.
Using the BAPGM enrichment blood culture platform, cPCR, qPCR, and enrichment blood culture PCR (ePCR) (Duncan et al. 2007, Oteo et al. 2017), Bartonella spp. DNA was amplified from blood specimens or BAPGM enrichment blood cultures from the father, mother, and all four children at various study time points. Bartonella spp. DNA was not amplified from blood, serum, or BAPGM enrichment blood cultures from the maternal grandmother. No subculture isolates were obtained at any time during the study. Based on qPCR testing, Bartonella DNA was amplified and sequenced directly from the blood of the father, two sons, and youngest daughter. Bartonella DNA was only amplified from the mother and eldest daughter after BAPGM enrichment blood culture.
Species identification was attempted for all PCR-positive samples by DNA sequencing of qPCR or conventional 16S–23S ITS or rpoB (β-subunit of bacterial RNA polymerase) PCR products as previously described (Carrasco et al. 2014, Oteo et al. 2017). Negative and positive PCR results for individual family members over time are summarized in Table 2. Due to the small amplicon size, discrimination of the Bartonella sp. infecting the father was not possible, and the mother's qPCR amplicon failed sequencing. By qPCR ITS amplicon sequence analyses, B. henselae DNA (100% similarity, 139/139 bp to GenBank accession AF369527) was amplified from the eldest daughter's enrichment blood culture. Following cPCR, sequences of the larger ITS region amplicons from the blood and blood culture of the eldest son also aligned with B. henselae strain California 1 with 100% similarity (525/525 bp to GenBank accession no. AF369527), 99% similarity (524/525 bp) to B. henselae strain Houston I (GenBank accession no. BX897699), 92.3% similarity (485/525 bp) to B. koehlerae strain bothieri (GenBank accession no. KF437497), and 63.5% similarity (344/525 bp) to B. quintana (GenBank accession no. AF368391).
A Bartonella sp. most closely related to B. quintana (98% similarity, 119/122 bp to GenBank accession no. AF368391) was amplified from the youngest son's blood specimens. DNA sequence alignments of rpoB PCR products from the eldest son's blood and youngest daughter's enrichment blood culture were also most closely related to B. quintana (627/656 bp, 95.6% similarity to GenBank accession no. AF165994) and to B. koehlerae strain bothieri (93.8% similarity to GenBank accession no. KX499340). With the exception of two base pairs, the 656 bp rpoB sequences from these two children were identical. Amplification of Bartonella DNA by cPCR was possible only for qPCR-positive samples with Ct (cycle threshold) values of ≤32, suggesting that higher Ct values indicated DNA copy numbers below the threshold limit for cPCR amplification. Anaplasma, Babesia, Ehrlichia, and hemotropic Mycoplasma spp. DNA was not amplified from blood specimens from any family member.
B. quintana DNA was amplified and sequenced from the family cat's postmortem tissues with 100% similarity (330/330 bp to B. quintana strain Oklahoma, GenBank accession no. AF368391), Bartonella DNA was not amplified from the dog's postmortem tissues.
Discussion
In this family, and in other familial case reports to date, the route(s) of transmission of Bartonella spp. to or among family members remains unclear. As illustrated by the spectrum of potential animal reservoir and vector exposures among individuals within this family, it is possible that the source(s) and mode(s) of transmission vary among and within families and are influenced by poorly defined environmental, social, microbiological, and epidemiological risk factors. The initial goal of this investigation was to determine which Bartonella sp. or spp. were responsible for B. henselae seroreactivity documented in all family members when tested by the CHU Sainte-Justine Laboratory. Initially, bacterial DNA sequences amplified from three sick children aligned with either a known (B. henselae) or an unreported (B. quintana-like) Bartonella sp. In contrast to the B. henselae antibody titers reported from Canada, when first tested on a research basis in May 2015 using a panel of 6, cell culture grown Bartonella spp. antigen, IFA antibodies were either not detected or were only detectable at a 1:64 dilution in two of six family members. As all four children had been treated with antibiotics for 2 months before submitting blood for research testing, it was possible that antimicrobial therapy influenced the initial research-based IFA serology and compromised achieving BAPGM enrichment blood culture/PCR results; however, neither the mother nor father was PCR positive at this testing time point and neither had been treated with antibiotics.
We have previously reported discrepancies between IFA serology and PCR amplification results, through which antibodies were not detected by a broad antigen panel, despite PCR amplification of Bartonella spp. DNA from blood, BAPGM enrichment blood cultures, or from subculture isolates. For example, a cross-sectional study (Maggi et al. 2012) found that 75% of Bartonella-infected humans did not have IFA antibodies to the infecting Bartonella species or genotype. The discrepancy between Bartonella spp. serology and PCR may represent falsely negative serology due to unresolved technical IFA testing issues, immunological anergy induced by chronic intravascular infection, or other as yet undetermined factors. Regardless of cause, contradictory serology and PCR findings became the basis to pursue further One Health collaborative investigations (Breitschwerdt 2014, 2017) spanning nearly 2 years, and were supported to a substantial degree by numerous attending and consulting physicians, veterinarians, microbiologists, regional public health officials, and the family, particularly the mother.
The ePCR platform was initially developed to enhance the microbiological confirmation of Bartonella spp. bacteremia in dogs (Duncan et al. 2007), but has subsequently been used to document Bartonella spp. infections in cerebrospinal fluid, joint fluid, and various pathological effusions in companion and wild animals, and humans (Cherry et al. 2011, Breitschwerdt 2014, 2017). Despite the use of multiple Bartonella spp. primer sets, no amplicons were generated from the grandmother or from any PCR-negative control. Also, no Bartonella or non-Bartonella isolates were obtained following subculture from the BAPGM liquid flask onto blood agar, reflecting the well-recognized fastidious nature of Bartonella spp. As PCR results from the initial blood specimens provided microbiological support for Bartonella spp. bacteremia, testing was repeated in September 2015, with no family member receiving antibiotics between May and September. Four of seven family members were now seroreactive to B. vinsonii subsp. berkhoffii genotypes II and III and B. henselae. Also, the eldest daughter and father were Bartonella spp. PCR positive, but only following direct extraction from blood, whereas all PCR results from blood, serum, and BAPGM enrichment blood cultures for the remaining family members were negative. Collectively, these results illustrate ongoing sensitivity limitations with the growth of Bartonella spp. from patient blood specimens.
As is common among many families in rural areas, this family had extensive exposure to numerous pets, to peridomestic wild animals, and to a spectrum of potential arthropod vectors. In recent years, an increasing number of documented or suspected modes of Bartonella spp. transmission have been reported, in conjunction with an expanding number of animals (reservoir and incidental hosts), in which Bartonella spp. have been identified (Regier et al. 2016, Breitschwerdt 2017). These and other factors have created clinical, epidemiological, and diagnostic challenges for microbiologists, physicians, animal health workers, and public health officials attempting to clarify the role and risks of Bartonella spp. infections between animals and humans. In addition to vector-competent arthropods such as sand flies, lice, fleas, biting flies, bedbugs, rat mites, and spiders, needle sticks, intraocular inoculation, blood transfusions, and potentially other modes of transmission seemingly occur (Breitschwerdt 2014, 2017, Regier et al. 2016). Previously, we described ecological diversity of Bartonella spp. infecting pet dogs and their owners in Virginia (Cherry et al. 2011). Similar to the Virginia report, DNA of more than one Bartonella spp. was amplified and sequenced from the Canadian family members and B. quintana from their cat. As the youngest daughter and eldest son had similar timing in illness onset after visiting the family vacation cottage where feral cats and numerous wildlife species were located, it is possible that initial exposures for these children occurred there rather than at the family residence. It is also possible that individual family members experienced additional Bartonella spp. exposures during the course of this study, which might have influenced temporal trends in serology and PCR findings.
Consistent with the prototypical manifestations of cat scratch disease (CSD), lymphadenopathy and fever were prominent findings in two children; however, these abnormalities were not self-limiting as expected with CSD. Recently, French investigators identified three novel Bartonella spp. strains that may have contributed to undifferentiated chronic illness in humans who were bitten by ticks (Vayssier-Taussat et al. 2016). Although nonspecific, the symptom constellation described by members of this family is consistent with reported findings in other recent studies involving Bartonella bacteremic individuals (Cherry et al. 2011, Maggi et al. 2011, 2013, Lantos et al. 2015, Vayssier-Taussat et al. 2016). In contrast to self-limiting CSD, “immunocompetent” humans can develop Bartonella spp. bacteremia that persists for months or perhaps years (Breitschwerdt et al. 2010a and Balakrishnan et al. 2013). Also, bacteremia has been reported in outwardly healthy blood donors (Pitassi et al. 2015, Oteo et al. 2017) as well as healthy or sick veterinary workers (Lantos et al. 2015). Based on multivariate logistic regression, bloodstream infection with B. henselae or B. clarridgeiae in Brazilian blood donors was associated with cat contact or history of tick bite (Diniz et al. 2016).
Despite PCR documentation of Bartonella spp. DNA in blood, serum, or enrichment blood cultures in this study, we do not know if these bacteria were a cause, a cofactor, or were totally unrelated to the reported illnesses. However, as reflected by the large number of specialists who provided medical care to this family, no other definitive infectious, noninfectious, or environmental cause of illness was found. Toxin exposure was not found during an environmental screen of the family residence and well water testing for toxic chemicals. Despite near-simultaneous treatment of all family members with an extended course of antibiotics, clinical improvement was often incomplete or similar symptoms reoccurred months after the course of antibiotics was completed. Due to variability or a lack of sensitivity, neither sequential serological testing nor BAPGM ePCR triple draw PCR results proved useful to guide clinical decision-making as to whether infection persisted or had been eliminated immunologically or in association with antibiotic administration.
Enhanced understanding of the pathogenesis of naturally occurring Bartonella spp. bacteremia may ultimately benefit from a comparative infectious disease causation approach (Breitschwerdt et al. 2013). With the advent and application of molecular diagnostic techniques (polymerase chain reaction, whole-genome sequencing, and others), and ongoing efforts to enhance the microbiological culture techniques to identify fastidious bacteria in patient blood samples in diagnostic and research laboratories around the world, it has become increasingly clear that a spectrum of bacteria gain access to the vasculature of healthy and sick humans (Vayssier-Taussat et al. 2015, Païssé et al. 2016). Because flea, tick, and other arthropod vectors require blood for nutrition, various bacteria, protozoa, and viruses have evolutionarily adapted to infect mammalian blood cells, thereby resulting in persistent intravascular infections that facilitate subsequent vector acquisition and perpetuation of the vectored microbial species (Vayssier-Taussat et al. 2015, Regier et al. 2016, Breitschwerdt 2017). Immunologically, many vector-borne organisms have concurrently developed a highly sophisticated repertoire of immune evasive mechanisms, so as to facilitate long-standing infection within the host, without inducing serious or life-threatening pathology. In contrast to most vector-borne bacteria that preferentially infect only one cell type, Bartonella spp. are unique in their ability to infect numerous cell types within the host, including dendritic cells, erythrocytes, monocytes, neutrophils, pericytes, and endothelial cells (Vayssier-Taussat et al. 2015, Regier et al. 2016, Breitschwerdt 2017). The ability to invade numerous cell types may contribute to a varying and diverse spectrum of symptoms reported by patients.
Conclusion
As illustrated by the findings reported for this family, the extent to which Bartonella spp. represent a component of the microbiome of healthy individuals or the pathobiome of sick individuals awaits future research studies utilizing more sensitive diagnostic techniques. However, based on recently reported microbiological observations, physicians and microbiologists should be aware of the potential for familial bloodstream infections with one or more Bartonella spp.
Footnotes
Acknowledgments
The authors thank the family, in particular the mother, and the many physicians, public health officials, and veterinarians for contributing information summarized in this study. Specifically we thank Marie-Christine Leduc, DMV IPASV, Hôspital Veterinaire des Moulins, Mascouche, QC, for providing medical record data for the family's cat and dog and the McGill University Health Centre, Vaccine Study Centre research staff who collected blood from family members, and carefully packed and shipped the blood draws to North Carolina for Bartonella testing.
Research Funding
This research was supported, in part, by donations to the North Carolina State University Veterinary Medical Foundation Fund for Bartonella and Vector-Borne Diseases Research and the state of North Carolina. Canadian contributors donated their time and resources in support of this study.
Author Disclosure Statement
In conjunction with Dr. Sushama Sontakke and North Carolina State University, Edward B. Breitschwerdt, DVM, holds U.S. Patent No. 7,115,385; Media and Methods for cultivation of microorganisms, issued October 3, 2006. He is a cofounder, shareholder, and chief scientific officer for Galaxy Diagnostics, a company that provides advanced diagnostic testing for the detection of Bartonella spp. infections. Dr. Ricardo Maggi is the chief technical officer for Galaxy Diagnostics. All other authors have no potential conflicts of interest to report.
Supplementary Material
Supplementary Data
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.