
Abstract
Nipah virus (NiV) and Hendra virus (HeV) are closely related members within the genus Henipavirus, family Paramyxoviridae, for which fruit bats serve as the reservoir. The initial emergence of NiV infections in pigs and humans in Malaysia, and HeV infections in horses and humans in Australia, posed severe impacts on human and animal health, and continues threatening lives of humans and livestock within Southeast Asia and Australia. Recently, henipavirus-specific antibodies have also been detected in fruit bats in a number of sub-Saharan African countries and in Brazil, thereby considerably increasing the known geographic distribution of henipaviruses. Africa is progressively being recognized as a new high prevalence zone for henipaviruses, as deduced from serological and molecular evidence of past infections in Madagascar, Ghana, Republic of Congo, Gulf of Guinea, Zambia, Tanzania, Cameroon, and Nigeria lately. Serological data suggest henipavirus spillover from bats to livestock and human populations in Africa without reported clinical disease in any of these species. All virus isolation attempts have been abortive, highlighting the need for further investigations. The genome of the Ghanaian bat henipavirus designated Ghana virus (GhV), which was detected in a pteropid Eidolon helvum bat, is the only African henipavirus that has been completely sequenced limiting our current knowledge on the genetic diversity and pathogenesis of African henipaviruses. In this review, we summarize the available data on the circulation of henipaviruses in Africa, discuss potential sources for virus spillover, and highlight existing research gaps.
Introduction
The genus Henipavirus within the order of Mononegavirales, family of Paramyxoviridae comprises five species that have all been detected within the past 25 years. Of these, Hendra virus (HeV) and Nipah virus (NiV) are closely related novel emerging zoonotic RNA viruses, to which three species have been added more recently, namely Cedar virus (CedV), Ghana virus (GhV), and Mojiang virus (MojV). With the extension of the known geographic distribution of henipaviruses outside Southeast Asia and Australia into Africa and lately also into Brazil, the global attention and relevance of these viruses has considerably increased.
Thus the objectives of this review are to summarize the existing data on the molecular and serological evidence confirming the circulation of henipaviruses in Africa and their spillover from bats to livestock and humans, and second to discuss possible risk factors associated with the emergence and spillover of henipavirus infection in Africa.
Morphology and Genome Organization of Henipaviruses
Henipaviruses harbor a nonsegmented, single-stranded negative sense RNA genome consisting of a helical nucleocapsid encased in an envelope, forming spherical to pleomorphic virus particles. Their genome codes for six major structural proteins from the 3′ to 5′ terminal (Fig. 1); nucleoprotein (N), phosphoprotein (P), matrix protein (M), fusion glycoprotein (F), attachment glycoprotein (G), and large polymerase protein (L) (Wang et al. 2000, 2001, Bellini et al. 2005, ICTV 2011, Rota and Lo 2012, Afonso et al. 2016). The P gene encodes for accessory proteins or nonstructural proteins designated C, V, and W, which can be translated by the use of alternative translation starts or by messenger RNA (mRNA) editing mechanisms and which are able to compromise the host's immune system (Yoneda et al. 2010, Basler 2012).
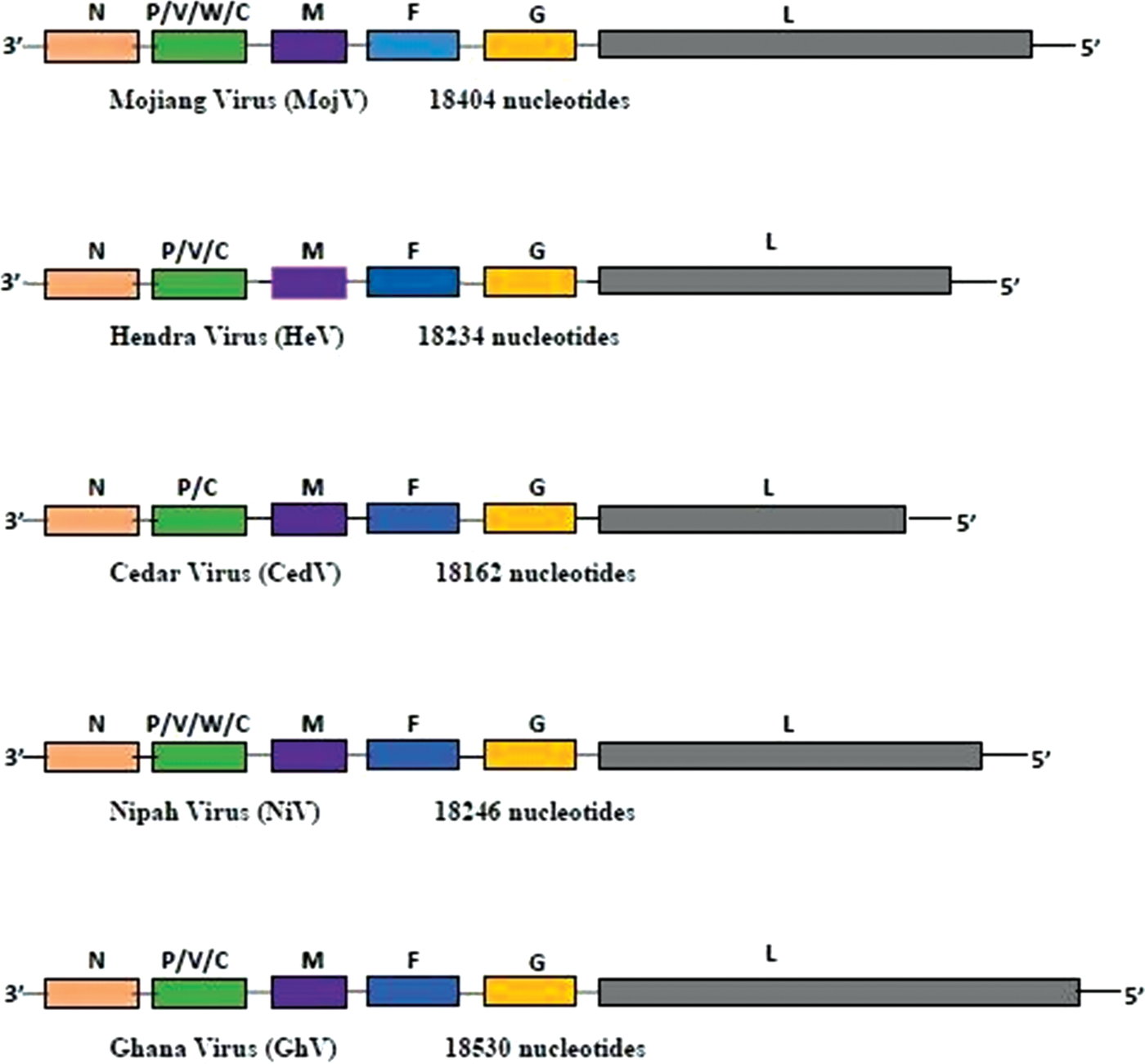
Organization of henipavirus genes. F, fusion glycoprotein; G, attachment glycoprotein; L, large polymerase protein; M, matrix protein; N, nucleoprotein; P, phosphoprotein; V/C/W, nonstructural proteins. Color images are available online.
The N, M, and L genes of NiV and HeV are highly conserved and are exploited for molecular diagnostic (PCR) purposes. The nucleotide homologies between NiV and HeV are 79% for the N gene, 77% for the M gene, and 87% for the L gene (Harcourt et al. 2000, Wang et al. 2001). Meanwhile, the amino acid sequence identity between the N proteins of NiV and HeV is 92%, whereas the G proteins of both viruses display an amino acid sequence identity of 83% (Harcourt et al. 2000), which is relevant for serological diagnostic assays.
The cellular receptor used by the different representatives of this virus family may influence the virulence, pathogenesis, and pathogenicity according to the respective tissue distribution of the receptor. The viral surface G protein facilitates the binding of the virus to the cellular surface, which is mediated by cellular ephrin B2 and ephrin B3 in the case of NiV and HeV (Bonaparte et al. 2005, Negrete et al. 2005, Xu et al. 2012), only ephrin B2 in the case of CedV (Marsh et al. 2012) and most probably GhV (Lee et al. 2015), whereas the cellular receptor for the MoV G protein is still to be elucidated, because the ephrin receptor binding motif is absent in this protein (Rissanen et al. 2017). The fusion protein, F0, is synthesized as an inactive precursor that requires cleavage by a host cell protease to become active subunits F1 and F2. Thus these subunits mediate the fusion of the virion membrane with the plasma membrane of the host cell (Wang et al. 2001, Diederich et al. 2005).
Although antibodies against the N protein are generated early after infection because of the abundant expression of this protein (Chua et al. 2000, Harcourt et al. 2000), the G protein is expressed on the surface of infectious viral particles and thus induces neutralizing antibodies (Zhu et al. 2006). Although the serum neutralization test that is considered the gold standard in henipavirus serology requires Biosafety Level 4 (BSL-4) facilities, N and G proteins are broadly used as suitable antigens for serological screening by enzyme-linked immunosorbent assay (ELISA) (Tamin et al. 2002, McNabb et al. 2014, Fischer et al. 2018). After the implementation of a commercial vaccine against HeV infections in horses in Australia that is based on the soluble HeV G protein, the parallel determination of antibodies against HeV N and G proteins can be used for the differentiation between infected and vaccinated animals (DIVA approach).
Henipaviruses in Southeast Asia, Australia, and China
NiV and HeV can cause severe respiratory illness, encephalitis, and neurodegenerative diseases in humans and animals with case fatality rates reaching up to 40–75% (Selvey et al. 1995, Chua et al. 1999, Chua 2003, McCormack 2005, Eaton et al. 2006, ProMED-mail 2011). Because of their zoonotic potential, broad host range, and high virulence and also the current absence of vaccines and therapeutics, both viruses have been classified as BSL-4 agents as the only members of the Paramyxovirus family. Fruit bats of the genus Pteropus, suborder Megachiroptera, are widely described as the natural reservoir for henipaviruses (Young et al. 1996, Halpin et al. 1999, 2011, Enserink 2000, Field et al. 2001, Yob et al. 2001), except MojV, which is considered to have a rodent reservoir host, Rattus flavipectus (Wu et al. 2014).
HeV first emerged in 1994 in Hendra a suburb of Brisbane, Australia, leading to an outbreak of severe respiratory disease in 21 horses of which 14 died, and including the fatal infection of a horse trainer (Murray et al. 1995a, 1995b, Nowak 1995, Selvey et al. 1995). Since its initial detection in Australia, HeV spillover from flying foxes to horses has regularly occurred, with an increase in the disease events in 2011–2012 (
NiV was first identified during a major outbreak of an acute respiratory disease in pigs in Sungai Nipah, Malaysia, between 1998 and 1999, which resulted in the death of 105 persons/pig-farm workers among 265 human infections and the extirpation of 1.1 million pigs, which were considered to act as intermediate hosts (Centers for Disease Control and Prevention [CDC] 1999, Chua et al. 1999, Parashar et al. 2000, AbuBakar et al. 2004). This was followed by a related smaller outbreak in Singapore in 1999 (Chew et al. 2000). Again, flying foxes were identified as the reservoir host of this virus (Chua et al. 2000, 2002, Enserink 2000, Yob et al. 2001).
Since 2001, NiV spillover infections from bats to humans are being reported from Bangladesh and India, where, without any involvement of pigs in the transmission cycle, humans became infected through the consumption of NiV contaminated date palm sap (Luby et al. 2006, 2009, Rahman and Chakraborty 2012, Islam et al. 2016). Sequence analysis revealed a high degree of similarity between the NiV strains circulating in Bangladesh and India (AbuBakar et al. 2004, Harcourt et al. 2005). Very recently, another NiV outbreak was reported in India and led to widespread public anxiety (Paul 2018), underlining the importance of public relations regarding zoonotic diseases. During these outbreaks, human-to-human infections occurred mostly by direct contact with infected patients or by contact with contaminated surfaces in hospitals and homes (Gurley et al. 2007, Sazzad et al. 2013, Chakraborty et al. 2016).
In the following years, NiV infections have also been detected in livestock species such as cattle, goats, and pigs in Bangladesh (Chowdhury et al. 2014), whereas the infection of dogs had already been reported in Malaysia in 2009 (Mills et al. 2009). Furthermore, the first NiV infection of horses was observed in the Philippines in 2014, and horse-to-human infections were postulated to have occurred through handling and consumption of horse meat, whereas human-to-human infections occurred by direct contact with infected subjects (Ching et al. 2015).
Ever since the emergence of NiV and HeV, studies on henipaviruses have extensively been conducted in Southeast Asia and Australia. In fact, fruit bats in Malaysia (Yob et al. 2001), Cambodia (Olson et al. 2002, Reynes et al. 2005), Thailand (Wacharapluesadee et al. 2005, 2016), Indonesia (Sendow et al. 2010, 2013), Vietnam (Hasebe et al. 2012), Papua New Guinea (Breed et al. 2010, 2013, Field et al. 2013), China (Li et al. 2008), and India (Yadav et al. 2012, 2018) were tested positive for the presence of NiV or HeV-specific antibodies and/or henipavirus-related RNA, whereas only HeV RNA and HeV-specific antibodies were detected in pteropid fruit bats in Australia (Field et al. 2011, Smith et al. 2011, Edson et al. 2015).
Results from studies in Australia indicated that urine is the most plausible source of HeV infection from bats to bats and bats to horses (Smith et al. 2011, Barr et al. 2015, Edson et al. 2015) and that virus excretion rather occurs periodically than continuously (Field et al. 2011). In the course of these screening efforts, CedV was detected and successfully isolated from Pteropus alecto fruit bats in Australia (Marsh et al. 2012). The lack of the alternative editing of the P gene, resulting in the translation of the V gene, may contribute to its majorly reduced, if not absent, pathogenicity in bats as well as in ferrets and guinea pigs after experimental exposure (Marsh et al. 2012).
Sequence analysis revealed nucleotide homologies between CedV and NiV or HeV to be 59% and 58% for N, 60% in both cases for M, and 50% in both cases for L (Marsh et al. 2012), which is considerably lower than the similarities between these proteins of HeV and NiV. Moreover, the considerably lower antigenic similarity of the CedV G protein and the G proteins of HeV or NiV (29% and 30%, respectively) results in a lack of cross-reactivity of antibodies raised against HeV G or NiV G proteins with CedV G (Marsh et al. 2012). Shortly afterward, another member of the Henipavirus genus designated MojV was detected in rats (R. flavipectus) in China in the course of a molecular screening of bats and rodents after fatal respiratory infections in three mine workers in the Yunnan Province (Wu et al. 2014).
Sequence analysis revealed nucleotide homologies between MojV and the other known henipaviruses to be 53–57% for N, 60–63% for M and 56–59% for L (Wu et al. 2014), which is again considerably lower than the similarities between these proteins of HeV and NiV (Harcourt et al. 2000, Wang et al. 2001), but comparable with the similarities between HeV or NiV and CedV (Marsh et al. 2012). Virus isolation attempts remained unsuccessful in this study (Wu et al. 2014).
Of interest, virus isolation attempts have been highlighted in several studies across Southeast Asia and Australia, but successful isolations were only reported for HeV in Australia (Marsh et al. 2010, Smith et al. 2011) and NiV in Malaysia (Chua et al. 1999, 2001, 2002, AbuBakar et al. 2004) and Bangladesh (Harcourt et al. 2005). Of late, antibodies against henipaviruses were also detected in bats in Brazil (De Araujo et al. 2017), thereby expanding the geographic distribution of henipaviruses to new territories. However, Africa is becoming another hotspot for the circulation of henipaviruses, for example, from the indigenous Eidolon helvum bat that is widely distributed across the continent.
Serological and Molecular Evidence of Henipaviruses in Bats and Potential Spillover into Livestock and the Human Population in Africa
An ample number of studies in Madagascar (Iehlé et al. 2007), Ghana (Hayman et al. 2008, Drexler et al. 2009, 2012), the Republic of Congo (Weiss et al. 2012), Gulf of Guinea (Peel et al. 2012), Zambia (Muleya et al. 2013), Tanzania (Peel et al. 2013), Cameroon (Pernet et al. 2014), and Nigeria (Olufemi et al. 2015) have effectively revealed the presence of henipavirus-specific antibodies and henipavirus-related RNA in African fruit bats. Some of these studies have shown that there has also been exposure of pigs, horses, and humans to henipaviruses in Africa suggesting spillover events from bats to livestock and to human populations (Hayman et al. 2011, Pernet et al. 2014, Olufemi et al. 2015). Figure 2 illustrates the locations in sub-Saharan Africa where henipavirus-specific antibodies or henipavirus-related RNA were detected so far.
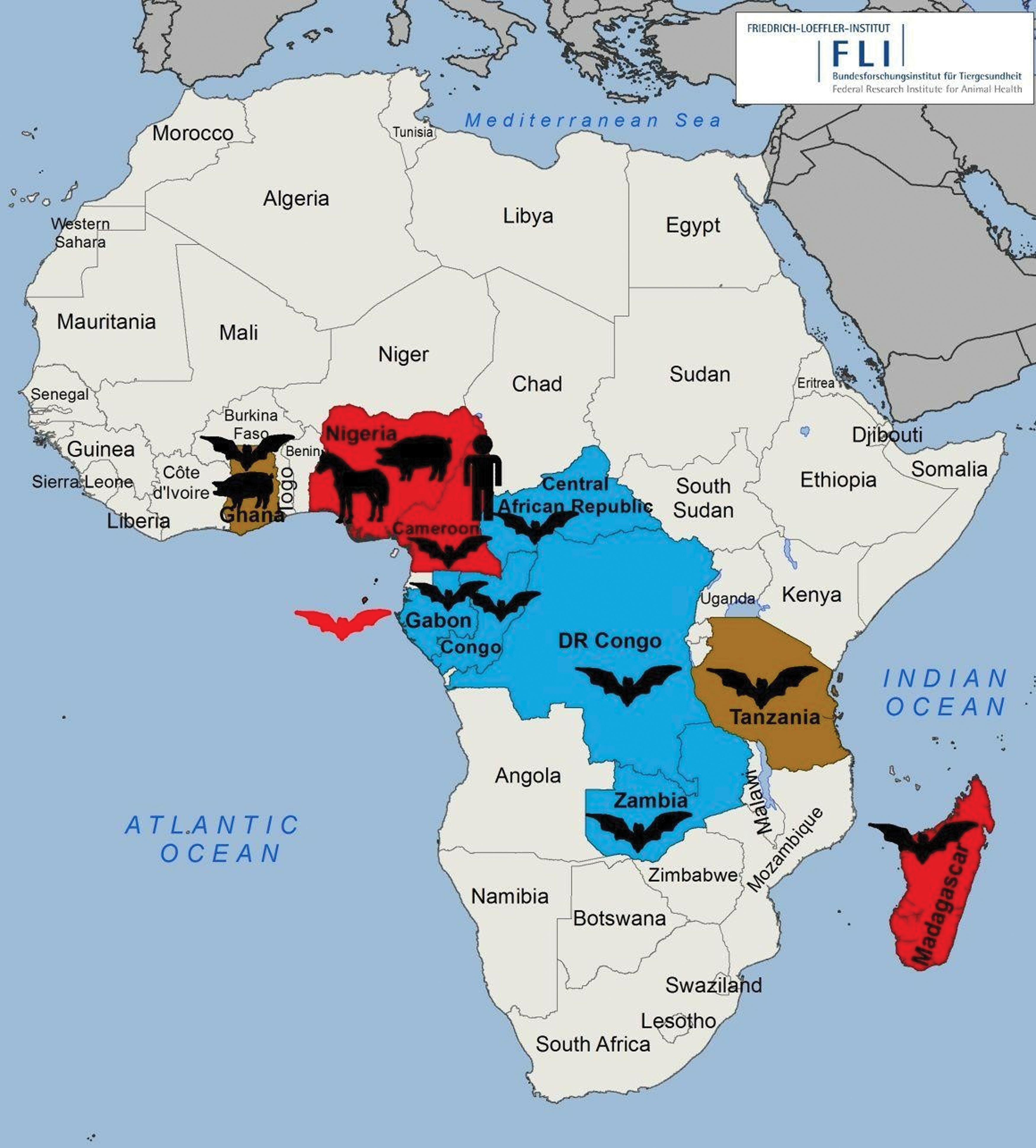
Serological and molecular evidence of henipavirus infections in sub-Saharan Africa. Red: countries with positive results in serological screening; brown: countries with positive results in serological and PCR screening; blue: countries with positive results in PCR screening. Color images are available online.
The first molecular evidence of henipavirus infection in bats in Africa was reported in a West African country, Ghana, in which fruit bats of the species E. helvum were sampled for their fecal droppings. A total of 215 fecal samples were obtained from an estimated colony of ∼400,000 bats in the Zoological Garden of Kumasi, Ghana (Drexler et al. 2009). Three of 215 were confirmed positive by a broad-spectrum RT-PCR targeting the L-gene of henipavirus, morbillivirus, and respirovirus (Tong et al. 2008). Sequencing of the amplicons revealed that the three viral sequences were closely related to the so far known henipaviruses but one was more closely to NiV. However, virus isolation was unsuccessful (Drexler et al. 2009).
A serological study in the same country revealed henipavirus-specific antibodies in Ghanaian fruit bats (Hayman et al. 2008), whereas a later study in 2011 confirmed a seroprevalence of 5% in 97 domestic pigs tested by Luminex binding assay but with no cross-neutralizing antibodies to NiV or HeV, suggesting previous exposure of Ghanaian pigs to serologically related but distinct henipaviruses (Hayman et al. 2011).
The entire genome of an African henipavirus, GhV, was successfully sequenced in a study that involved sampling of bats in Ghana, Republic of Congo, Gabon, the Central African Republic (RCA), and the Democratic Republic of Congo (DRC) (Drexler et al. 2012).
The first serological evidence of henipavirus-specific antibodies in African horses was revealed during a cross-sectional survey of horses and pigs in Zaria and environs in Kaduna state, Nigeria (Olufemi et al. 2015). A total of 200 and 310 sera were sampled from horses and pigs, respectively. All samples were analyzed using an indirect ELISA based on HeV-sG antigen (CSIRO Australia). The species-specific seroprevalence was 15.5% and 20% for horses and pigs, respectively. It was noticed that horses managed under intensive system, used for sports and traditional purposes, consumed homemade food, consumed well water, and were kept outdoors were at higher risk to henipavirus infection. The highest risk factors for pigs to contract a henipavirus infection included import, intensive management with homemade feed and well water, and being kept outdoors without a feed storage plan (Olufemi et al. 2015). However, results from this study should be interpreted carefully because the study lacks confirmatory testing, for example, by Western blot or immunofluorescence assay detecting antibodies against other viral antigens such as the nucleoprotein.
Moreover, the first evidence of henipavirus spillover into human populations of Africa was detected in Cameroon (Pernet et al. 2014). In this study, sera were sampled from 44 E. helvum fruit bats and 497 humans. These samples were subjected to a vesicular stomatitis virus (VSV)-based pseudoparticle seroneutralization assay to detect NiV (cross-) neutralizing antibodies. A seroprevalence of 48% and 3–4% was observed in bats and humans, respectively. Consequently, the seropositive human sera were found almost exclusively in individuals involved in butchering bats for bushmeat and residing in proximity to areas undergoing deforestation (Pernet et al. 2014). Furthermore, another study conducted in Brazzaville, Republic of Congo, aimed at investigating the risk of zoonotic disease emergence through hunting of bats and preparation and consumption of bushmeat. A total of 339 samples collected from 42 E. helvum bats destined for human consumption were obtained from local hunters at the market. All samples were tested by a broad-spectrum RT-PCR targeting the L-gene of respirovirus, morbillivirus, and henipavirus (Tong et al. 2008), and by a quantitative RT-PCR (RT-qPCR; Weiss et al. 2012) that was used to determine the viral load in the tissues. Henipavirus-related sequences were detected in 15 samples from 11 individual bats and virus load ranged from 1.1 × 102 to 3.4 × 104 copies per tissue sample of ∼0.3 cm3 and 4 samples could not be quantified because of low viral load in tissues (Weiss et al. 2012).
In East Africa, during an epidemiological study to screen for paramyxoviruses in frugivorous E. helvum bats in Zambia, 312 bats were captured at the Kazanka National Park from which the corresponding number of spleen samples were collected (Muleya et al. 2013). Samples were tested for paramyxovirus-related sequences using a semi-nested RT-PCR (Tong et al. 2008) and further sequenced and phylogenetic analysis performed. Among the seven novel paramyxovirus sequences detected, five were closely related to the genus Henipavirus. The presence of these viruses in bats, livestock, and human populations in Africa poses a potential public health risk although no outbreak is yet reported within this continent and as such continuous surveillance of these viruses in bat–livestock–human populations is primordial. Table 1 presents an overview of serological and molecular studies performed so far in Africa.
Overview of Serological and Molecular Studies Performed in Africa on the Presence of Henipaviruses in Different Host Species
∼, closely related to; Afr, African; CI, confidence interval; DRC, Democratic Republic of Congo; ELISA, enzyme-linked immunosorbent assay; HeV, Hendra virus; HeV sG iELISA; Hendra virus recombinant soluble G-glycoprotein indirect immunosorbent assay; LBA, Luminex binding assay; N/A, not applicable; NGS, Next Generation Sequencing; NiV, Nipah virus; qPCR, quantitative PCR; RCA, Central African Republic; SNT, serum neutralization test; VNT, virus neutralization test; VSV, vesicular stomatitis virus.
Possible Risk Factors Associated with Henipavirus Emergence and Spillover in Africa
Disease emergence results from the dynamic interactions between pathogen, animal, and/or human hosts and the perpetually changing environment. These interactions are at the core of disease emergence; therefore, understanding these drivers and impacts will allow an evidence-based risk assessment and the development of mitigation strategies that will enable an effective and timely response to new disease outbreaks (Wang and Crameri 2014).
The frugivorous bat species E. helvum is highly abundant particularly across the sub-Saharan region and also present on some offshore and isolated islands (Thomas and Henry 2013). Fruit bat colonies undergo annual transcontinental migration after the rainfall gradient to suitable feeding grounds or the seasonal changes in food availability, with reports of individual bats covering >2500 km yearly (Richter and Cumming 2008, Ossa et al. 2012, Fahr et al. 2015). Generally, they roost in urban settlements and feed in close proximity to human and livestock populations, thereby increasing the possibility of disease agents such as henipaviruses to cross species barriers and infect new hosts, resulting in putatively harmful or even fatal disease outbreaks (Peel et al. 2013).
In addition, husbandry practices and management in developing nations in Africa allow close contact between bats and livestock populations because animals are mostly kept free-ranging. There is a high probability for bats to contaminate their food, pasture, and water with secretions and excretory products thus culminating to infection (Peel et al. 2013, Olufemi et al. 2015).
Moreover, deforestation causes habitat fragmentation and severely impacts natural habitats of bats, forcing them to further migrate toward urban centers and thereby increasing the possibility of contact with domestic animals and humans, potentially leading to cross-species transmission of viral agents (Weiss and McMichael 2004, Wolfe 2005, Lloyd-Smith et al. 2009, Murray and Daszak 2013). The altered habitat may also influence the density of the local bat population, thereby affecting the spillover risk (Plowright et al. 2015). Bat migration toward urban centers could also alter their feeding habits, which might subsequently affect their immune responses by limiting their ability to combat or clear the viruses they harbor and possibly increase the probability of virus spillover to other species (Plowright et al. 2015, 2017, Giles et al. 2018, Kessler et al. 2018, Páez et al. 2018).
Indeed, socioeconomic and cultural activities like bat hunting, sale, handling, butchering, and consumption as bushmeat is considered one of the primordial risk factors associated with henipavirus spillover to human populations in Africa (Weiss et al. 2012, Pernet et al. 2014). However, humans may also be infected through contact with environmental virus contamination by, for example, inhaling virus particles from bat secretions and excretions upon intrusion into bat roosting areas (Weiss et al. 2012). Although no specific disease symptoms have been associated with henipavirus infections in bats, pigs, and humans in Africa (Hayman et al. 2008, Drexler et al. 2009, Pernet et al. 2014), there is a possibility of underreporting of clinical cases connected with such infections. This is especially the case because febrile flu-like infections in humans that may also develop into encephalitis occur frequently in sub-Saharan Africa without further investigation into the infectious source. It is therefore important to continue the screening for such infections and to associate these virological results with clinical data of the sampled individuals.
Limits or Gaps in the Study of Henipaviruses in Africa
So far, GhV is the only African henipavirus that has been completely sequenced (Drexler et al. 2012). Not much is known on the real prevalence of henipavirus infection in Africa and only very little is known on the genomic diversity of henipaviruses circulating at the interface between bats, livestock, and humans in sub-Saharan Africa. All efforts to isolate African henipaviruses and even the attempt to generate a recombinant replication competent GhV have failed so far, making it impossible to really assess their pathogenic, antigenic, and zoonotic potential (Hayman et al. 2008, Drexler et al. 2009, Weiss et al. 2012) and their public health risk.
Cross-sectional serological studies provide no details on viral transmission dynamics within populations. However, valuable information on the presence or absence of virus infection may be obtained and consequently, further studies are required to understand virus–host interaction and mechanisms of cross-species transmission. Only one study indicated a potential spillover of an African henipavirus into the human populations in Cameroon without reporting on clinical cases (Pernet et al. 2014). It is worth mentioning that the human sera used in this study were archival samples collected between 2001 and 2003, thus suggesting that human exposure to henipaviruses or henipa-like viruses might have occurred before the first known report of a circulation of these viruses.
Therefore, more studies should be undertaken as henipaviruses with a higher pathogenicity might be circulating in Africa, possibly causing isolated clinical cases in humans and/or animals. Multidisciplinary studies on henipaviruses involving bats, livestock, and humans have not yet been conducted throughout Africa; the previous studies targeted just individual aspects.
So far, almost all serological screening and confirmatory tests have been performed using the glyco- and nucleoproteins of either HeV or NiV, thus possibly detecting cross-reactive antibodies that are not specifically targeting GhV. However, our present knowledge suggests that the applied diagnostic measures cannot fully identify the assumed diversity of circulating henipaviruses in Africa. Only one assay has been used to date that is able to specifically detect antibodies against GhV and to correlate these results to HeV or NiV-specific serological results (Pernet et al. 2014). Therefore, there is still a lack of serological studies from different areas in Africa and different (animal) species to specifically detect antibodies against GhV, and to correlate the cross-reactivity of those sera to HeV or NiV-specific antigens, which would enable a better assessment of the variety and prevalence of different henipaviruses in Africa.
Conclusion and Recommendations
This review summarizes the current knowledge on the circulation of henipaviruses on the African continent, particularly in sub-Saharan Africa. Besides a number of studies in indigenous bat species, there is evidence for the exposure of livestock and humans to henipaviruses. In this light there is need for intensified national and regional surveillance and monitoring programs to provide adequate information on the epidemiology of henipaviruses in Africa. The programs should target wildlife, livestock, and companion animal species as well as human populations and their ecosystem in view of promoting the One Health concept. For a noninvasive sampling approach, molecular analysis is most promising from urine samples. However, if tissue samples like kidney, spleen, and brain are available for molecular analysis along with serology, the results might become much more rewarding.
There is also need for more serological and molecular studies in Africa using reliable and specific diagnostic tools that could discriminate the known species of henipaviruses to reveal the true prevalence of henipavirus infections in Africa. To complement that, a very important milestone would be to isolate the respective virus(es).
Livestock owners, farmers, hunters, traders, veterinarians, epidemiologists, foresters, health personnel, researchers, military personnel, and other stakeholders should be stringently trained on biosafety and biosecurity measures to be respected during sample collection in the field. Multiministerial and multidisciplinary teams should work in close collaboration with international organizations to effectively implement the correct biosafety and biosecurity measures. Hence, this integrated and collaborative approach will serve as an efficient tool in the detection and further characterization of henipaviruses that are circulating within the continent.
Footnotes
Acknowledgments
This work was made possible through the financial support of the German Research foundation—DFG (DFG grant GR 980/4–1). Special thanks to the Friedrich-Loeffler Institut (FLI), Germany, through its fruitful collaboration with the National Veterinary Laboratory (LANAVET), Cameroon and the Laboratory for Public Health Research Biotechnologies (LAPHER Biotech.) under the Biotechnology Centre—University of Yaoundé 1 (BTC-UY1), Cameroon. The authors also thank Nicole Reimer for preparing the map displaying the sampling and henipavirus screening studies in African countries that have been published so far.
Author Disclosure Statement
The authors declare no competing financial interests.