
Abstract
Sporadic outbreaks of Rift Valley fever virus (RVFV), a zoonotic, mosquito-borne Phlebovirus, cause abortion storms and death in sheep and cattle resulting in catastrophic economic impacts in endemic regions of Africa. More recently, with changes in competent vector distribution, growing international trade, and its potential use for bioterrorism, RVFV has become a transboundary animal disease of significant concern. New and sensitive techniques that determine RVFV presence, while lessening the potential for environmental contamination and human risk, through the use of inactivated, noninfectious samples such as formalin-fixed, paraffin-embedded (FFPE) tissues are needed. FFPE tissue in situ hybridization (ISH) enables the detection of nucleic acid sequences within the visual context of cellular and tissue morphology. Here, we present a chromogenic pan-RVFV ISH assay based on RNAscope® technology, which is able to detect multiple RVFV strains in FFPE tissues, enabling visual correlation of RVFV RNA presence with histopathologic lesions.
Introduction
Rift Valley Fever virus (RVFV) (Family: Phenuiviridae, genus: Phlebovirus) is a zoonotic arbovirus endemic to Africa and the Arabian Peninsula. It is an enveloped, negative-sense RNA virus with a tripartite genome, consisting of the ∼6.4 kb large (L), ∼3.8 kb medium (M), and ∼1.7 kb small (S) segments (Faburay et al. 2017).
RVF is considered to be a high consequence transboundary animal disease and a potential public health concern. Work with it requires BSL-3 containment and in the US, also Select Agent clearance. RVF outbreaks in ruminants are characterized by abortion storms and neonatal deaths, and to a lesser degree, morbidity and mortality in adult ruminants. Infected tissues must be handled with care due to their zoonotic risk of causing human infections, which can vary from mild febrile illness to severe liver disease, encephalitis, or even death, often occurring following exposure to infected animal fluids or tissues (Faburay et al. 2017).
RVFV has a broad tissue tropism but is typically found in the liver, where it causes a necrotizing hepatitis, in lymphoid tissues, and in the kidney (Odendaal et al. 2018). In experimental animal studies with virulent strains, viral nucleoprotein is readily detectable by immunohistochemistry (IHC) in tissues until 5 days postinfection (Faburay et al. 2016, Wilson et al. 2016). While, histopathologic lesions consistent with RVF are seen at later time points postinfection, current virus detection techniques typically yield negative results.
Formalin-fixed, paraffin-embedded (FFPE) tissues offer a safe format for handling and testing viral pathogens (Haddock et al. 2016). IHC and to a lesser extent PCR and in situ hybridization (ISH) have been used for pathogen detection in these tissues. However, to date, only IHC, the detection of viral antigen, has been used for RVFV and serves as a confirmatory diagnostic test in South Africa (Odendaal et al. 2018). While both ISH and PCR detect viral RNA (vRNA) in tissues, ISH, similar to IHC, additionally provides histomorphologic context, thereby enabling the correlation of microscopic lesions with the presence of vRNA (Cassidy and Jones 2014).
Recent technological advances have improved the accuracy and accessibility of RNA ISH. One such technology, RNAscope® (Advanced Cell Diagnostics [ACD], Hayward, CA), uses short branched DNA probes to enhance the detection of vRNA in FFPE tissues and may be as sensitive as existing qPCR technology (Wang et al. 2012, Vasquez et al. 2018). To our knowledge, so far RVFV ISH has only been used to detect RVFV RNA in fixed cells (Wichgers Schreur and Kortekaas 2016). Therefore, the objective of this study was to develop a highly sensitive ISH assay for the detection of RVFV RNA in FFPE tissues.
Materials and Methods
The RVFV-infected FFPE ruminant tissues used in this study originated from previous experimental studies (Faburay et al. 2016, Wilson et al. 2016) and the 2010 South African outbreak (Odendaal et al. 2018). RVFV infection was previously confirmed using either RVFV-specific triplex or M segment RT-qPCR, for the experimentally and naturally infected animals, respectively (Faburay et al. 2016, Wilson et al. 2016, Odendaal et al. 2018). In the experimental studies, 4–5-month-old lambs and calves were inoculated subcutaneously with 2 mL of 1 × 106 plaque-forming units of RVFV Kenya 128-15B 2006 (Ken06) or Saudi Arabia 2001-1322 (SA01) (Faburay et al. 2016, Wilson et al. 2016). We tested pan-RVFV FFPE tissue ISH on liver from a Ken06 4 days postinoculation (dpi) sheep and a liver and lymph node from 5 dpi Ken06 sheep, lambs #55 and #44, respectively (Faburay et al. 2016). We also tested one liver sample from a 3 dpi Ken06-infected calf, #41, and two liver samples from a 4 dpi SA01-infected calf, #37 (Wilson et al. 2016). Finally, we tested a mixed tissue block, from SA2010 outbreak adult sheep 10-3268, containing lung, liver, spleen, and kidney (Odendaal et al. 2018). Uninfected control tissues for all tissue types and species tested came from either mock-inoculated animals in the aforementioned experimental studies or were species matched necropsy tissues from the Kansas State Veterinary Diagnostic Laboratory (KS VDL).
ACD synthesized the pan-RVFV ISH probe according to the investigator-provided target sequence and a list of excluded sequences, including host species and nearest neighbor viruses. Ten sequences of RVFV representative of phylogenetic variance of up to 5% in the target region of the L segment were aligned, including MP-12, Ken06, Clone 13, ZH548, ZH501, Smithburn, Lunyo, SA-57, SA-51, and SA01. The resulting probe (516121-V-RVFV-L) targets a consensus sequence located between nucleotides 20-997 of the RVFV segment L, complete genome (NCBI Reference Sequence: NC_014397.1). ACD recommended negative and positive control probes targeting dihydrodipicolinate reducatase B of Bacillus subtilis and sheep housekeeping gene peptidylprolyl isomerase B (PPIB), respectively, were also used.
The chromogenic ISH assays were conducted according to the manufacturer's protocol. In brief, 4 μm tissue sections were prepared using standard procedures of the KS VDL. These were rehydrated, pretreated, hybridized, and detected using the RNAscope 2.5 HD Brown Kit and then hematoxylin counterstained and mounted. For RVFV RT-qPCR on the previously untested SA2010 FFPE tissue block, we used a single tube deparaffinization and tissue digestion protocol followed by automated magnetic bead extraction or silica based spin column protocol for recovery of RNA and the RVFV triplex RT-qPCR assay (Wilson et al. 2013). Additional modifications made were reduction of the final concentration of primers and probes, halving of the volume of nucleic acids used, and use of a 20 min reverse transcriptase reaction with the PCR master mix (Quanta BioSciences). Cycle threshold (Ct) values reported are the means of three replicates for RVFV L, M, and S gene detection. IHC results were previously published (Faburay et al. 2016, Wilson et al. 2016, Odendaal et al. 2018).
Results
The pan-RVFV ISH assay readily identified vRNA in lesions in both species' livers (Fig. 1A, B) and did not label uninfected liver (Fig. 1C). The assay also worked well on the other infected tissue types (Fig. 1D). The negative and positive control probes performed as expected, including the sheep PPIB positive control probe on cattle tissue, as expected due to close gene homology (data not shown).
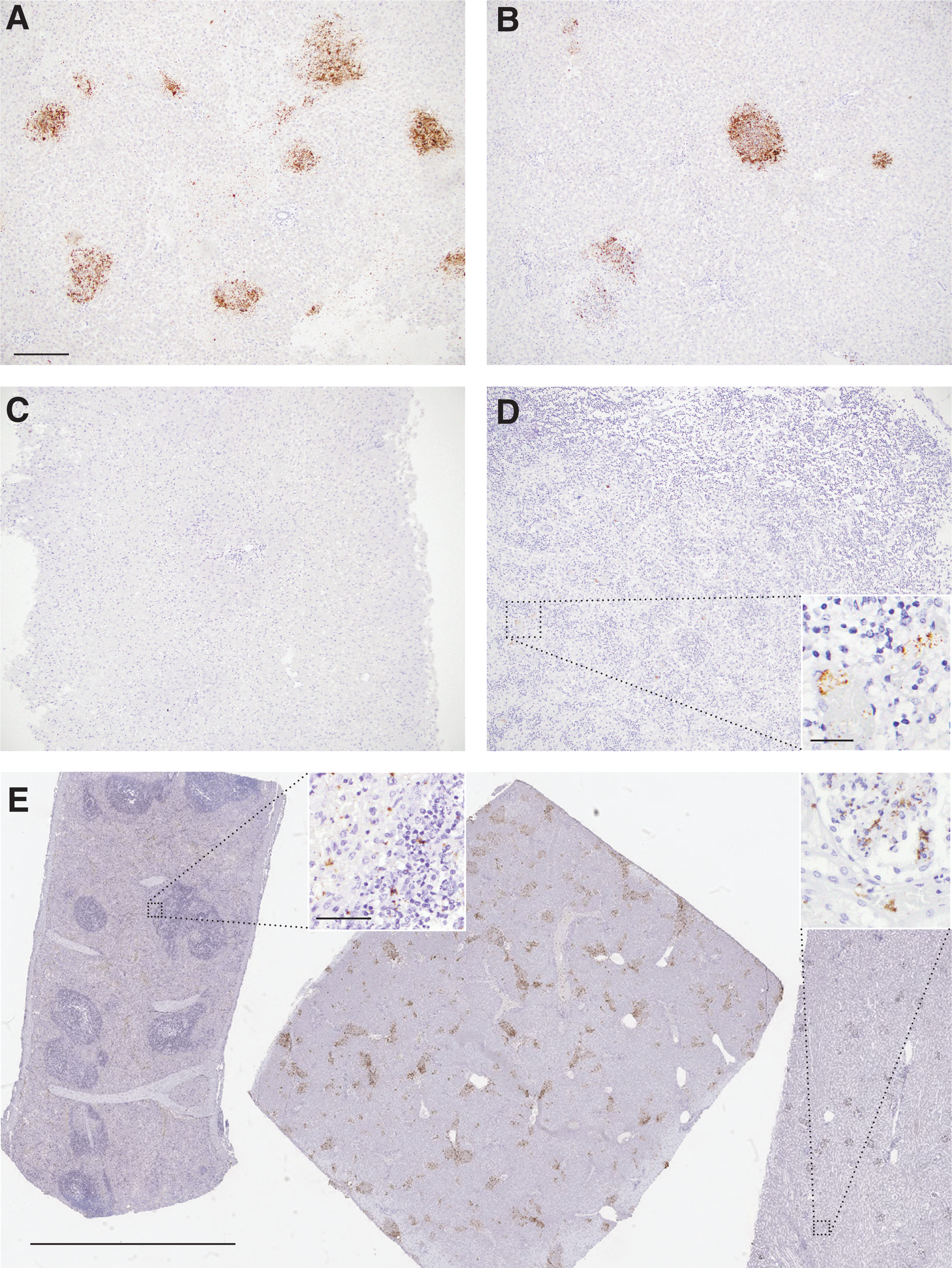
Pan-RVFV ISH detects multiple strains of RVFV in ruminant tissues. The pan-RVFV ISH probe labeled RVFV L segment vRNA (brown) in Ken06 infection-induced hepatic necrosis lesions in a sheep
Table 1 summarizes IHC, PCR, and ISH results for all tissues. The ISH results correlated 100% with the triplex RT-qPCR and prior IHC (Faburay et al. 2016, Wilson et al. 2016) with the exception of one liver sample from calf #37 SA01 and sheep 10-3268's lung, kidney, and spleen. The Ken06 tissue blocks were all heavily infected as indicated by low mean RT-qPCR Ct values, ranging from 21.3 to 23.6. In the liver, ISH labeling correlated with histopathology lesions and in the lymph node vRNA was found in the cytoplasm of circulating macrophages as reported previously by IHC (Faburay et al. 2016). Subjectively, ISH produced a stronger, more widespread and more readily interpreted signal than seen previously with IHC (Faburay et al. 2016, Wilson et al. 2016, Odendaal et al. 2018).
In Situ Hybridization Correlates with Immunohistochemistry and RT-qPCR
Mean Ct of triplicates, bead extraction.
Separately collected sample from same organ that was processed into a separate FFPE block.
PCR was run on all tissues as a mixed tissue block specimen; consequently, all PCR results listed are the same for each organ.
Ct, cycle threshold; FFPE, formalin-fixed, paraffin-embedded; IHC, immunohistochemistry; ISH, in situ hybridization; N/A, nonapplicable; ND, not detected; RVFV, Rift Valley fever virus.
The SA01 liver samples from calf #37 were selected for their low vRNA level (Ct 30), determined using RT-qPCR on fresh unfixed tissue, and observed viral antigen in only one tissue block (Wilson et al. 2016). Moreover, the IHC negative block was also initially negative when tested by FFPE tissue triplex RT-qPCR using both column and bead extraction techniques with 100 μL lysate. However, increasing the lysate volume to 200 μL for bead extraction yielded a suspect result, specifically a mean Ct of 35.1 for the M segment. By pan-RVFV ISH, both liver blocks were positive, although the vRNA labeling was less pronounced for the block that was previously negative by the other tests.
The SA2010 outbreak sheep mixed tissue block 10-3268, from a sheep with a positive liver by M segment RT-PCR on fresh unfixed tissue (Odendaal et al. 2018), was positive by triplex RT-qPCR (mean Ct values of 20.9/20.4/23.7 for L/M/S segments respectively). By IHC, the liver was strongly positive, while the lung, kidney, and spleen were negative (Odendaal et al. 2018). However, on histopathologic review, there was a moderate amount of necrosis in the spleen, and the kidney had a moderate amount of acute tubular injury. In the liver, stronger labeling with similar intralesional distribution was seen by ISH when compared with IHC. Interestingly, all four organs were ISH positive, although lung, kidney, and spleen were more sparsely labeled than the liver (Fig. 1E).
Discussion
This is the first report of a pan-RVFV ISH for detection of RVF vRNA in FFPE tissues. RNAscope ISH technology is a highly sensitive RNA ISH platform previously shown to accurately detect viral nucleic acids in FFPE tissues (Vasquez et al. 2018). We found that pan-RVFV ISH readily identified vRNA, regardless of infecting strain in all species' tissues tested (Fig. 1). In addition, in this study, the ISH results correlated well with results from IHC and RT-qPCR results. Finally, the SA01 low vRNA experimental block and the SA2010 outbreak mixed tissue block results suggest that pan-RVFV ISH is more sensitive than IHC and possibly RT-qPCR on FFPE tissues.
Further evaluation of pan-RVFV ISH, including establishment of the assay's diagnostic accuracy and precision, is underway.
Conclusions
The RNAscope pan-RVFV ISH assay has great potential as a confirmatory diagnostic test or research tool, including for use with archival FFPE tissues. Importantly, ISH may offer superior sensitivity to IHC while still enabling testing to be conducted outside high containment on verified inactivated RVFV-infected FFPE tissues. Unfortunately, high costs currently limit the application of the RNAscope pan-RVFV ISH assay as a routine diagnostic test, but the authors are hopeful that these costs will drop as the technology matures.
Footnotes
Acknowledgments
The authors thank the KS VDL histology laboratory for slide sectioning. This work was funded by the USDA Agricultural Research Service and the Department of Diagnostic Medicine/Pathobiology, College of Veterinary Medicine, Kansas State University's startup funds for Dr. Davis. This work was also supported through the Department of Homeland Security (grant no. 2010-ST061-AG0001), Center of Excellence for Emerging and Zoonotic Animal Diseases (CEEZAD). This work was done in partial completion of Izabela Ragan's PhD dissertation.
Author Disclosure Statement
No competing financial interests exist.