
Abstract
Introduction:
Chikungunya virus (CHIKV) and West Nile virus (WNV) have previously been reported from several African countries, including those bordering Rwanda where they may have originated. However, there have been no serosurveillance reports from Rwanda regarding these two viral pathogens.
In this article, we present the first study of immunoglobulin G (IgG) seroreactivity of CHIKV and WNV in Rwandan blood donor samples.
Methods:
Blood donors from Rwanda (n = 874) and Sweden (n = 199) were tested for IgG reactivity against CHIKV, using an in-house enzyme-linked immunosorbent assay with the E1 envelope protein fused with p62 as antigen, and against WNV using a commercial kit. Data on mosquito distribution were obtained from the 2012 assessment of yellow fever virus circulation in Rwanda.
Results:
Seroreactivity to CHIKV was high in Rwanda (63.0%), when compared with Swedish donors, where only 8.5% were IgG positive. However, a cross-reactivity to O'nyong'nyong virus in neutralization test was noted in Rwandan donors. No significant difference in WNV seroreactivity was found (10.4% for Rwandan and 14.1% for Swedish donors). The relatively high seroreactivity to WNV among Swedish donors could partly be explained by cross-reactivity with tick-borne encephalitis virus prevalent in Sweden. Donors from the Eastern Province of Rwanda had the highest IgG reactivity to the two investigated viruses (86.7% for CHIKV and 33.3% for WNV). Five genera of mosquitoes were found in Rwanda where Culex was the most common (82.5%). The vector of CHIKV, Aedes, accounted for 9.6% of mosquitoes and this species was most commonly found in the Eastern Province.
Conclusions:
Our results showed high seroreactivity to CHIKV in Rwandan donors. The highest IgG reactivity to CHIKV, and to WNV, was found in the Eastern Province, the area reporting the highest number of mosquito vectors for these two viruses. Infection control by eliminating mosquito-breeding sites in population-dense areas is recommended, especially in eastern Rwanda.
Introduction
Awareness of mosquito-borne viruses, including chikungunya virus (CHIKV) and West Nile virus (WNV), is limited among health care institutions in Sub-Saharan Africa. In this region, arboviruses are often misdiagnosed due to lack of laboratory facilities and diagnostic tests (Stoler et al. 2014). Knowledge concerning the distribution of arbovirus mosquito vectors is often incomplete or absent (Kraemer et al. 2015), although such knowledge could be of help in assessing risk areas for such viruses (Nsoesie et al. 2016). Human-to-human transmission through administration of blood products contaminated with these arboviruses has previously been reported (de Mendoza et al. 2012, Appassakij et al. 2016). This is why investigation of blood donors is well motivated.
Little is known about the prevalence and clinical burden of mosquito-borne viruses in Rwanda, including the risk of arbovirus transmission through blood transfusion. Currently, the Rwandan National Center of Blood Transfusion systematically screens for four transfusion-transmissible infections: HIV, syphilis, hepatitis B virus (HBV), and hepatitis C virus (HCV). In 2005, serosurveillance of blood-borne viral infections among blood donors in Rwanda showed a prevalence of 1.05% for HIV, 2.76% for HBV, 1.4% for HCV, and 0.7% for syphilis. At that time, 55% of blood donors came from rural areas, 40% from schools, and 5% from urban areas. A large proportion (75%) of these blood donors were <25 years of age, and the majority were male (88%). Regular donors donated blood on average twice a year (MOH-Rwanda 2006).
Although the blood donor screening program in Rwanda is at present limited to these four infectious agents, the program might be extended to include other potentially transfusion-transmissible viruses such as human T cell lymphotropic viruses-I/II, cytomegalovirus, parvovirus B19, WNV, CHIKV, dengue virus, and hepatitis E virus (Allain et al. 2009, Laperche et al. 2015). In this study, we determined the seropositivity of two mosquito-borne viruses CHIKV and WNV in Rwandan blood donors from known locations to obtain information on the geographical distribution of these infections.
CHIKV was first isolated in Tanzania in 1952 since when infections have been reported from all over the world (Caglioti et al. 2013, Hrnjakovic Cvjetkovic et al. 2015). The spread of the Aedes albopictus mosquito vector into new locations, especially in Europe and the Americas, and the expansion of the CHIKV 06.21 strain associated with this vector are important factors linked to the current outbreaks of CHIKV (Vazeille et al. 2007, Roche et al. 2015, Rossati et al. 2015).
CHIKV is an Alphavirus within the Togaviridae family; it is an enveloped RNA virus with a negative-sense single-stranded nonsegmented genome. Transmission of this virus mainly occurs through the bite of the infected Aedes mosquito, which is commonly found in tropical and temperate regions. Major vertebrate hosts include monkeys and humans, in whom this virus may replicate efficiently (Diallo et al. 1999). Outbreaks among humans may have resulted from significant human migrations, viral mutations with adaptation to new mosquito vectors, and from climate change, which could have created a favorable environment for expansion of these vectors to new territories (Tsetsarkin et al. 2011).
Clinical severity of CHIKV infections in humans can vary profoundly. About 3–25% of CHIKV-infected patients remain asymptomatic (Burt et al. 2012). When symptoms do occur, they vary from moderate to severe. As indicated by its name, chikungunya, which derives from the Tanzanian Makonde word “kun qunwala,” meaning “to become contorted” or “that which bends up,” is mainly characterized by debilitating arthralgia and fever. Symptomatic CHIKV infection typically begins after an incubation period of 1–12 days with fever, symmetrical polyarthralgia, skin rash with itching, headache, muscle pains, and gastrointestinal symptoms (Borgherini et al. 2007, Burt et al. 2012). The disease may also affect the eyes and central nervous system (CNS); patients may demonstrate iridocyclitis and retinitis, meningoencephalitis with convulsions and confusion, and in rare instances Guillain–Barré syndrome. Other rare complications include myocarditis, hepatitis, and mild hemorrhage. Chronic, relapsing, and incapacitating arthralgia mainly affecting small joints may persist for up to 3 years (Lemant et al. 2008, Caglioti et al. 2013).
Congenital CHIKV often presents with severe clinical manifestations, such as respiratory distress, necrotizing enterocolitis, meningoencephalitis, myocarditis, edema, bullous dermatitis, and pericarditis, and is associated with a high mortality rate (Villamil-Gomez et al. 2015). Consequently, diagnosis and follow-up of CHIKV infections in pregnant women are imperative.
WNV was discovered in Uganda in 1937, and during the past decades outbreaks have been reported in North America and southern Europe, especially around the Mediterranean Basin between 2004 and 2013 (Hernandez-Triana et al. 2014, Chancey et al. 2015). WNV is a positive sense single-stranded RNA virus belonging to the Flavivirus genus within the Flaviviridae family. The main reservoir is birds, in which WNV replicates efficiently, and the virus is transmitted by the bite of the Culex mosquito. Humans and horses are incidental hosts. Affected migratory birds spread the infection to new areas along their flyways (Campbell et al. 2002, Hayes et al. 2005).
In humans, 80% of WNV infections are asymptomatic, and symptoms may vary from minor self-limiting signs to severe neurological complications (Chancey et al. 2015). Clinical manifestations include fever, headache, fatigue, and myalgia, sometimes with nausea and vomiting. Skin manifestations include a nonpruritic maculopapular morbilliform rash, predominantly over the torso and extremities. Serious CNS complications may occur, including meningoencephalitis, myelitis, and acute flaccid paralysis (Sejvar 2014).
Currently, no antiviral treatment or vaccine is available for CHIKV or WNV, and medical care relies solely on supportive measures. Thus, mosquito bite prevention is the most important public health strategy to fight these infections. However, novel therapeutic and prophylactic strategies including monoclonal antibodies, antiviral drugs, and vaccines are currently under trial (Abdelnabi et al. 2015, Ahola et al. 2015, Roques et al. 2017, Runowska et al. 2018). Diagnosis is usually confirmed by use of virus-specific RT-PCR analyses and/or serological testing to identify immunoglobulin M and/or immunoglobulin G (IgG) antibodies.
To estimate the presence of CHIKV and WNV infections in Rwanda, and the spread of their vectors, we investigated CHIKV and WNV seropositivity among blood donors in Rwanda, and analyzed the distribution of their respective mosquito vectors. We found high IgG seropositivity for CHIKV, but low IgG seropositivity for WNV, among Rwandan blood donors. Both infections were unevenly distributed within the country, with the Eastern Province reporting the highest seropositivity rates for both viruses. Culex mosquitoes were more common than Aedes mosquitoes and both genera were more abundant in the Eastern Province than in other regions of Rwanda. However, probable serological cross-reactivity to related viruses within their respective families was detected for both viruses.
Materials and Methods
Study population and data collection
Serum or plasma samples were drawn consecutively from 874 Rwandan and 199 Swedish blood donors to determine the seropositivity to CHIKV and WNV by enzyme-linked immunosorbent assay (ELISA). All samples from Rwanda and Sweden were collected in 2015. Characteristics (age and gender) of the Rwandan and Swedish blood donors were described previously (Seruyange et al. 2018). The Rwandan donors were 34.6% female and 65.4% male, whereas the Swedish donors were 50.3% female and 49.7% male. Mean age of Rwandan donors was 27.5 years (range: 17–62 years) and that of Swedish donors was 43.7 years (range: 18–75 years).
Rwandan blood donors were registered in compliance with the national blood transfusion policy (MOH-Rwanda 2006), and grouped according to province of residence: Eastern, Western, Northern, Southern, and Kigali City. In Sweden, blood donors were anonymously recruited from the Gothenburg area, and identified only by gender and age.
Data on mosquitoes were derived from an assessment report on the circulation of yellow fever virus in Rwanda during November and December 2012, and reproduced with permission from the Rwandan Biomedical Center. The number and the genus of collected mosquitoes were reported, and the geographical distribution of adult mosquito vectors of CHIKV and WNV was defined.
Methods
Plasma/serum sample collection
Samples of Rwandan plasma and Swedish serum were collected, frozen, and stored at −20°C, as previously described (Seruyange et al. 2018). All tests, except the neutralization tests that were performed at the Department of Clinical Microbiology of Umeå University, were performed at the Clinical Virology Laboratory, Sahlgrenska University Hospital in Gothenburg, Sweden.
Serological analyses for CHIKV
ELISA plates (Nunc, Maxisorp) were coated with an in-house CHIKV antigen that was expressed and produced as follows: the CHIKV Env p62-E1 protein antigen contains the ectodomains CHIKV-115 p62 and E1 joined by a glycine serine linker to form a soluble protein. The p62-E1 antigen was expressed and secreted from stably transfected Drosophila melanogaster Schneider 2 (S2) cells as a streptavidin-tagged soluble recombinant protein (Voss et al. 2010) at Karolinska Institute and stored at a concentration of 1 mg/mL.
First, we investigated the IgG reactivity of the CHIKV Env p62-E1 protein antigen with a high IgG titer serum obtained from a cynomolgus macaque immunized with a DNA-launched CHIKV replicon encoding the CHIKV E protein, and boosted with recombinant modified vaccinia Ankara strain expressing the same antigen (Roques et al. 2017). To define the optimal antigen dilution, we tested IgG reactivity to different concentrations ranging from 4 to 0.125 μg/mL. After incubation, we added different dilutions ranging from 1:500 to 1:64,000 of this positive serum and of human plasma samples at dilution ranging from 1:100 to 1:12,800. Thereafter, plates were washed and horseradish peroxidase (HRP) conjugated mouse antimonkey IgG (cat. no. 4700-05; Southern Biotechnology) was added at a dilution of 1:5000, followed by rinsing and addition of substrate. We found that a concentration of 0.25 μg/mL for the CHIKV antigen, diluted in carbonate buffer at pH 9.6, gave the best signal/background ratio when the monkey positive serum and the human samples (n = 11) were tested at a dilution of 1:200.
Then we performed the CHIKV IgG ELISA on plasma and serum samples from all the Rwandan and Swedish blood donors. Each plate included a positive human plasma control (obtained from a Mozambique patient who was RT-PCR positive to CHIKV in plasma, samples were generously provided by Dr. Kerstin Falk, Public Health Agency of Sweden, Stockholm) in duplicate, pooled negative controls in quadruplicate, and donor samples in duplicate. All serum/plasma samples and controls were diluted 1:200 in 1% dry milk in PBS +0.05% Tween 20 before being added to the antigen-coated ELISA plates; reactions were then incubated at 37°C for 90 min.
After washes, 100 μL of conjugate consisting of antihuman goat IgG conjugated with HRP (Jackson ImmunoResearch Europe Ltd., UK) diluted to 1:2000 was added to the plates, which were then incubated for 60 min at 37°C. After washing, 100 μL of substrate (10 mg O-phenylenediamine diluted in 10 mL citrate buffer +4 μL of oxygen peroxide) was added and wells were checked for reactivity (yellow). Finally, after 10 min of incubation in the dark, the reaction was stopped by adding 100 μL of sulfuric acid. The optical density (OD) values were read on a multiscan ELISA reader at 492 nm, minus background OD values read at 620 nm. The cutoff value was determined by the first point of OD that had the lowest number of samples (nadir, close to zero), observed on a drawn chart based on the OD values of all samples (Supplementary Fig. S1). Thus, OD values greater than the cutoff set at 0.45 were considered to be positive.
Plaque reduction neutralization testing for alphaviruses
A subset of samples (n = 40, 10 CHIKV ELISA IgG negative and 10 CHIKV ELISA IgG positive samples from both Rwandan and Swedish blood donors) were subjected to plaque reduction neutralization test (PRNT) confirmatory testing to study cross-reactivity between CHIKV and o'nyong'nyong virus (ONNV), a closely related alphavirus that is also endemic in sub-Saharan Africa.
PRNT titers were determined by using the method as previously described (LaBeaud et al. 2015). In brief, Vero B4 (African green monkey kidney) cells were seeded in 96-well plates (15,000 cells/well). Fifty microliters of threefold serial dilutions of the sera samples (diluted from 1/40 to 1/1080) was mixed with 50 μL of viruses (50 pfu), incubated for 1 h at room temperature, thereafter the mixture was transferred to Vero B4 cells. After additional incubation for 1 h at 37°C in a carbon dioxide incubator, the cells were overlaid with 100 μL of Dulbecco's modified Eagle medium containing 2% carboxymethylcellulose (CMC; Sigma Life Science). After 3 days, the CMC overlay was removed, and the cells were fixed with 4% paraformaldehyde solution before being stained with crystal violet. Visible plaques were counted and the titers are expressed as the reciprocal of the serum dilution that showed 80% of plaque reduction (PRNT80). Samples were considered as CHIKV PRNT positive or ONNV PRNT positive if the PRNT titer was >40.
Serologic analyses for WNV
The WNV IgG ELISA was performed using a commercial serological kit supplied by FOCUS Diagnostics (Cypress, CA) according to the manufacturer's instructions. The OD values were read at 450 nm in a spectrophotometer. WNV IgG ELISA results were classified by index values (calculated using plasma sample OD divided by calibrator solution OD) as positive (≥1.5), equivocal (<1.5 and ≥1.3), and negative (<1.3). Equivocal results were considered to be negative in the final analysis.
Serologic analyses for tick-borne encephalitis virus
All Swedish blood donors, but only those Rwandan donors who tested positive for WNV IgG, were tested for tick-borne encephalitis virus (TBEV) IgG antibodies to check for cross-reactivity between the two Flaviviruses. For that purpose, the Enzygnost Anti-TBEV IgG test (Siemens Healthcare AB, Upplands Väsby, Sweden) was used according to the manufacturer's instructions. Data on TBEV IgG seroprevalence in a subset (n = 140) of the Swedish blood donors were recently published separately (Veje et al. 2018).
The immunochemical reactivity of the TBE virus-specific IgG antibodies in the sample (absorbance value “A”) was measured by spectrophotometer. Results were expressed in U/mL and were classified based on the cutoff, which was lot dependent, as either positive (Asample > cutoff +0.1), equivocal (cutoff ≤ Asample ≤ cutoff +0.1), or negative (Asample < cutoff), according to the manufacturer's instructions.
Mosquito collection
Results from a previous study (MOH-Rwanda 2012) were deduced with permission. Eight collection sites from four different ecological zones in Rwanda were chosen based on rainfall, vegetation, and altitude, and further defined based on differences in humidity, temperature, and land cover use.
Trained community health workers collected adult mosquitoes by human landing catches between 4 PM and 8 PM on two consecutive days at all sites. All mosquitoes caught were morphologically identified in the field at the genus level and thereafter kept frozen for transportation to the National Reference Laboratory in Kigali, where they were frozen at −70°C before being shipped to the Pasteur Institute, Dakar, Senegal, for confirmation of species.
Statistical analysis
Differences between Rwandan and Swedish blood donors in seropositivity, expressed as number (n) and percentage (%) of seropositive individuals, were evaluated using the Pearson's chi-squared test for independence. Comparisons of mean IgG OD values were calculated using the Student's t-test for two independent samples. p Values <0.05 were considered significant. The IBM SPSS Statistics version 20 software was used for all statistical analyses.
Ethics
The study was cleared by the Rwanda Military Hospital Ethics Committee (ref. no. 002/010/2014). Study procedures were explained and informed consent forms were distributed and signed by all participants. The Swedish blood donors were analyzed anonymously and results could not be traced back to individuals, so ethical permission was not required according to Swedish regulations.
Results
Seroprevalence of CHIKV and WNV
IgG reactivity indicative of seroprevalence of CHIKV was 63.0% in Rwandan donors, which was significantly higher than the seroprevalence in donors from Sweden (8.5%, p < 0.00001). A substantial difference in geographical distribution was noted among the provinces of Rwanda; the lowest seroprevalence was 51.2% in the Southern Province and the highest was 86.7% (p < 0.00001) in the Eastern Province (Table 1; Fig. 1).

Distribution of CHIKV IgG seropositivity and adult Aedes specimens in Rwanda. Each color represents the seropositivity to CHIKV IgG within each province of Rwanda, whereas the size of the mosquito is proportional to the number of specimens of Aedes mosquitoes collected. The seropositivity to CHIKV IgG was the highest within the Eastern Province where Aedes mosquitoes were also more abundant. CHIKV, chikungunya virus; IgG, immunoglobulin G. Color images are available online.
Chikungunya Virus and West Nile Virus Immunoglobulin G Seropositivity in Rwandan Provinces and in Sweden [n (%)]
Chikungunya IgG seropositivity was significantly higher in Rwanda than in Sweden (p < 0.0001), and this was valid for all provinces of Rwanda. No significant difference was observed considering WNV IgG seropositivity between both countries (p = 0.14). The Eastern Province accounts for the highest IgG seropositivity with p value <0.0001 for both CHIKV and WNV, as compared with other provinces.
CHIKV, chikungunya virus; IgG, immunoglobulin G; WNV, West Nile virus.
The seropositivity to CHIKV increased with age among Rwandan donors (Fig. 2). A comparison between two age groups, donors ≤25 years old and ≥26 years old, revealed significantly higher CHIKV seropositivity among donors in the older group (77% vs. 51%, p < 0.0001). Compared with Swedish donors, CHIKV seropositive Rwandan blood donors demonstrated higher mean OD values (1.5 for Rwandan donors vs. 0.75 for Swedish donors). In Rwanda, IgG seropositivity to CHIKV was higher among males than among females (69% vs. 51%, p = 0.0001).
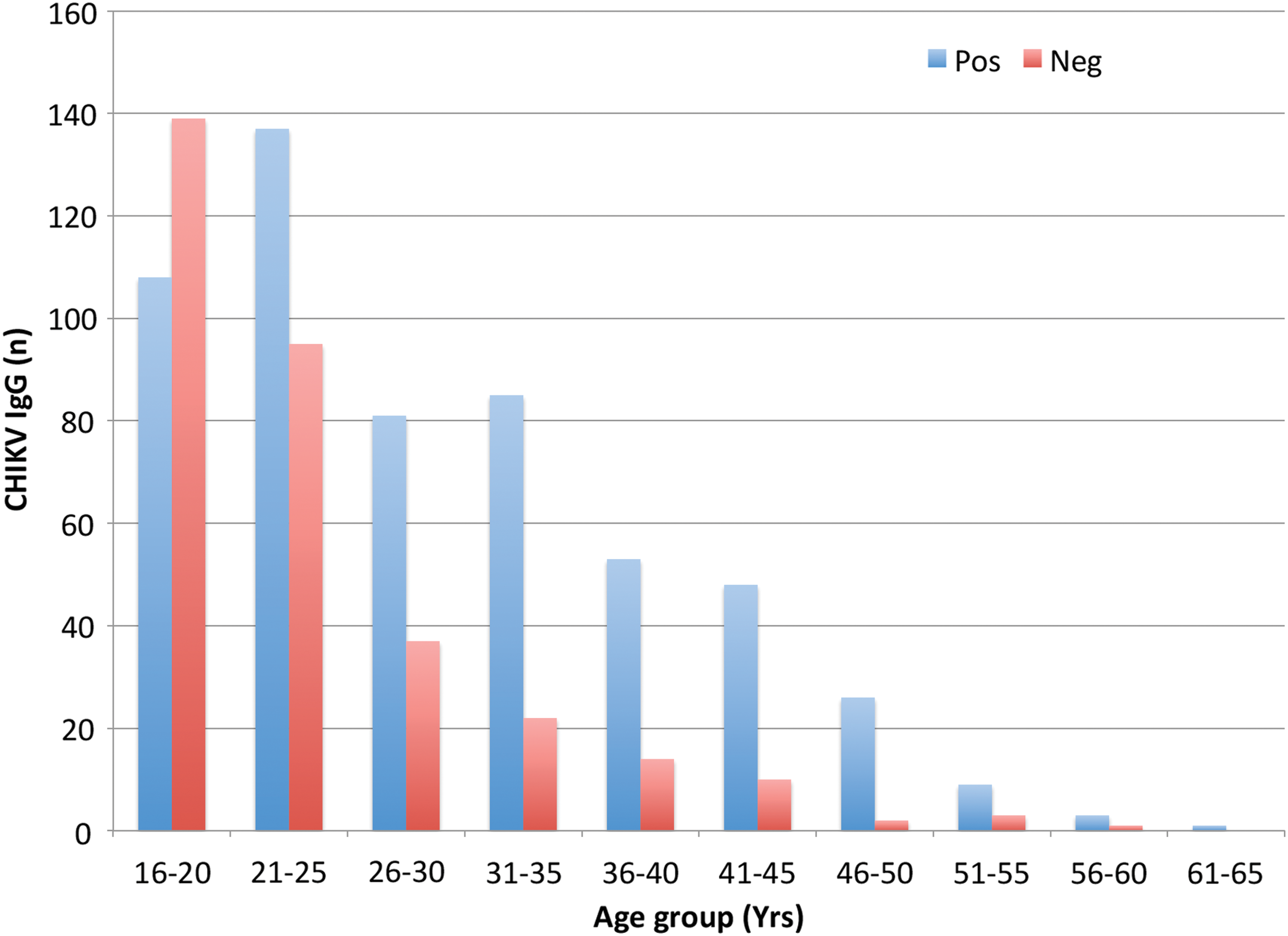
CHIKV IgG results and age group of Rwandan donors. The blue bars represent the number of CHIKV IgG seropositive donors whereas the red bars represent the number of seronegative donors. The proportion of CHIKV IgG seropositive donors increased with age. We compared CHIKV IgG results between donors ≤25 years old and those ≥26 years old using chi-squared test. Donors ≥26 years old were significantly the most seropositive; p < 0.0001. Color images are available online.
In contrast, WNV IgG seropositivity among Rwandan and Swedish donors showed no statistically significant difference: 10.4% were seropositive in Rwanda versus 14.1% in Sweden (p = 0.14). The Eastern Province in Rwanda also reported the highest seropositivity to WNV IgG at 33.3% (Table 1). WNV seropositivity was more common among male Rwandan donors than among females (12.8% vs. 5.9%, p = 0.0016), whereas no statistically significant difference in WNV seropositivity was found between male and female Swedish donors.
Only 7.3% (n = 78) of the 1073 blood donor samples from Rwanda and Sweden tested positive for both CHIKV and WNV IgG. Among Rwandan donors, 73 (8.3%) were seropositive for both viruses; 80.1% of WNV IgG seropositive donors also tested positive for CHIKV IgG. Only five (2.5%) Swedish donor samples were seropositive for both viruses.
Cross-reactivity between CHIKV and ONNV
From a subset of samples (n = 40, 20 Swedish sera and 20 Rwandan plasma samples) tested for cross-reactivity between the two alphaviruses CHIKV and ONNV using PRNT, all of the CHIKV ELISA positive and negative samples from Sweden and all of the CHIKV ELISA negative samples from Rwanda were found to be PRNT negative for both CHIKV and ONNV. Out of 10 CHIKV IgG positive Rwandan plasma samples, 2 were double negative, and 8 samples were double positive against both viruses in PRNT. In addition, four of these eight samples showed a higher PRNT titer against ONNV than CHIKV, two samples showed a higher PRNT titer against CHIKV than ONNV, whereas the other two samples were equally positive in PRNT against both viruses.
Cross-reactivity between WNV and TBEV
Of the 28 Swedish donors who were IgG seropositive for WNV, 22 (78.6%) were also IgG seropositive for TBEV. Somewhat surprisingly, among the 91 Rwandan donors who were IgG seropositive for WNV, 64 (70.3%) were also IgG seropositive for TBEV (Table 2). However, the Swedish donors who were IgG seropositive for TBEV demonstrated higher mean OD values than their Rwandan counterparts (1.34 vs. 0.65; p = 0.0001). Although values were retrieved from different ELISA tests, it is of interest to note that 13 of 28 Swedish donors demonstrated higher OD values for TBEV than for WNV, whereas none of their Rwandan counterparts did so (Fig. 3).
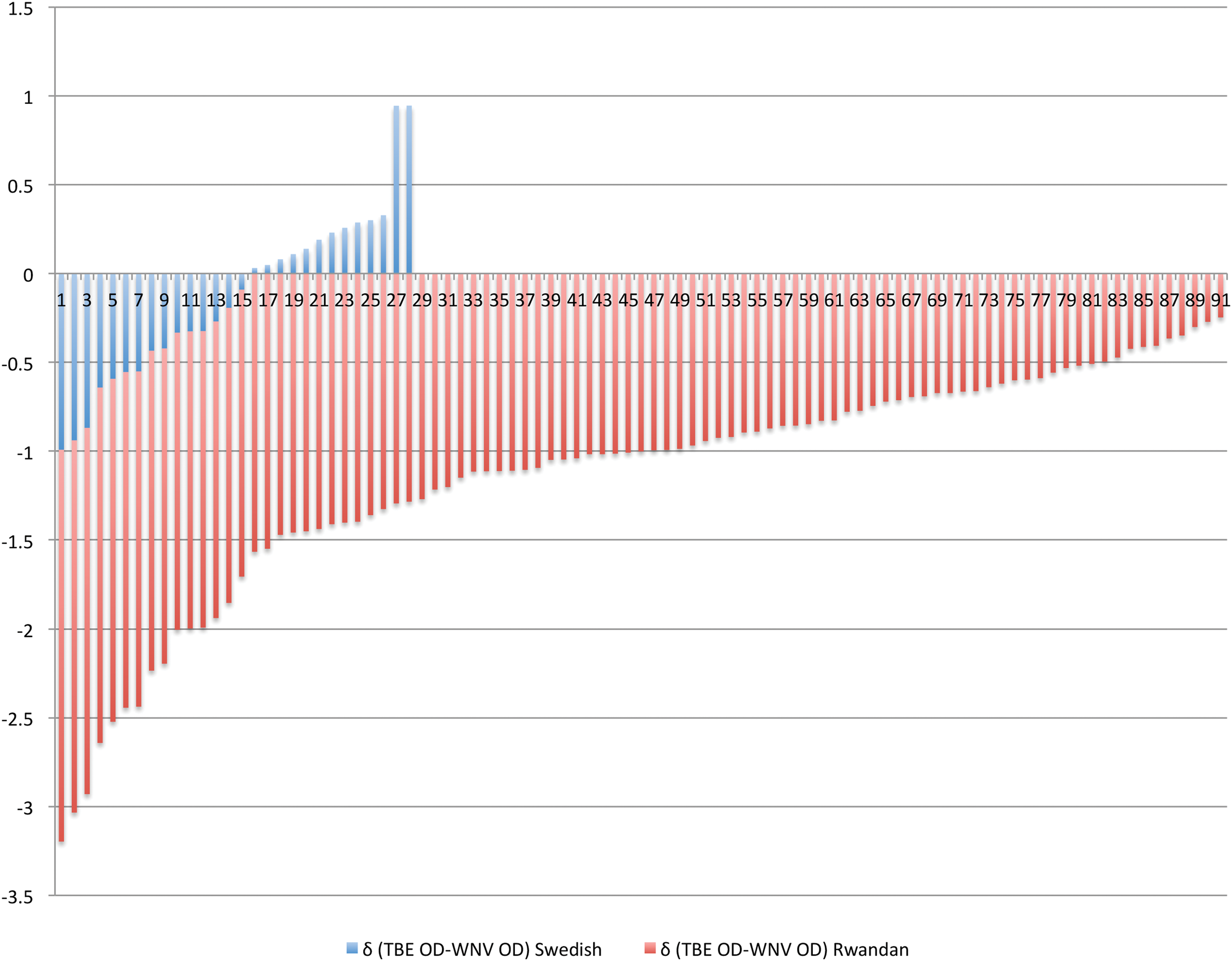
Difference (δ) in OD values between TBEV and WNV IgG from donors positive for WNV IgG. The difference of OD values (δ) between TBEV and WNV IgG of Swedish donors (blue vertical lines) shows 13 out of 28 donors with higher OD values against TBEV. None of Rwandan donors (in red vertical lines) showed higher OD values against TBEV. OD, optical density; TBE, tick-borne encephalitis; TBEV, tick-borne encephalitis virus; WNV, West Nile virus. Color images are available online.
Cross-Reactivity of West Nile Virus and Tick-Borne Encephalitis Virus Immunoglobulin G Tests in Rwandan and Swedish Blood Donors
Only Rwandan donors positive for WNV IgG (n = 91) and all Swedish donors (n = 199) were tested for TBEV IgG. Sixty-four (70.3%) of Rwandan donors and 22 (78.6%) of Swedish donors were double positive for TBE and WNV; there was no statistical difference (p = 0.4), but Swedish had higher mean OD values of TBEV IgG positive (1.34 vs. 0.647); p = 0.0001 and Rwandan had higher mean OD values of TBEV IgG negative (0.09 vs. 0.2); p < 0.0001.
OD, optical density; TBE, tick-borne encephalitis; TBEV, tick-borne encephalitis virus.
Collection of mosquitoes in Rwanda
A total of 1971 adult mosquitoes belonging to 5 genera were collected from eight collection sites in Rwanda during November and December 2012. Culex was the most abundant genus in all these regions with a total of 1626 (82.5%) of collected mosquitoes, followed by Aedes 190 (9.6%), Coquillettidia 95 (4.8%), Mansonia 41 (2.1%), and Anopheles 19 (1.0%) (Table 3). The Culex and Aedes mosquitoes were caught in all collection sites except in Musanze district. The highest number of collected specimens from each species was from the Eastern region (> 35% for Culex and >54% for Aedes), where the highest seropositivity was also found, as shown in Figure 1 for CHIKV.
Number of Mosquitoes Collected by Ecologic Zone in Rwanda, November–December 2012 (Reproduced by Permission from Rwanda Biomedical Center)
A total of 1971 adult mosquitoes belonging to 5 genera were collected: 190 (9.6%) Aedes, 19 (1.0%) Anopheles, 1626 (82.5%) Culex, 41 (2.1%) Mansonia, and 95 (4.8%) Coquillettidia. The genera Culex and Aedes were caught in all the ecologic zones with zone 4 (corresponding to the Eastern Province) having the highest number of specimens (>35% for Culex and >54% for Aedes), followed by zone 1 (32% for Aedes). Culex sp. was the most abundant genus in all the ecologic zones.
Discussion
Our data showed a high IgG reactivity to CHIKV envelope antigen, indicative of seroprevalence (>51%) in all Rwandan provinces among our study population, suggesting significant circulation of the virus in this country. The fact that no outbreaks have previously been reported may be due to herd immunity maintained through continuous infection, or, maybe more likely, to the absence of agent-specific viral diagnostics in the country. Cases of CHIKV infection in Rwanda may have been overlooked due to a syndromic approach and, therefore, misdiagnosed as malaria, rheumatoid arthritis, etc. Another explanation, based on the results of the PRNT showing reactivity to both CHIKV and ONNV, is that ONNV might also be circulating locally. However, the presence of the latter virus has not yet been investigated in patients or vectors in Rwanda.
Our data are comparable with those reported from blood donors in the Republic of Congo in 2011, where CHIKV seropositivity was 34.4% (Moyen et al. 2014). Four decades ago, CHIKV was reported to be the most common human arbovirus based on a seroepidemiological survey conducted from 1980 to 1982 in Burundi, a country bordering the Southern Province of Rwanda (Rodhain et al. 1987). Their study found no WNV seropositive samples; these findings may indicate that WNV is rare in this region. A more recent study in Kenya reported higher seroprevalence for CHIKV than for dengue virus (67% vs. 1.6%) (Grossi-Soyster et al. 2017).
The highest IgG reactivity for CHIKV and WNV, 86.7% and 33.3% respectively, was found in the Eastern Province of Rwanda as compared with the other Rwandan provinces (Western, North, South, and Kigali city), where mean seropositivity was 58.1% for CHIKV and 5.7% for WNV (Table 1). Similar findings were reported in Madagascar, where low seroprevalence for CHIKV was observed in higher altitude inland regions (Schwarz et al. 2012). Seropositivity for both viruses was higher among male Rwandan donors, possibly because men work in the forest industry and are more exposed to mosquitoes than women.
IgG seropositivity for CHIKV was significantly higher in Rwanda than in Sweden, but there was no significant difference between these two countries regarding seroreactivity to WNV. The relatively high positivity for WNV IgG among Swedish donors might be explained by cross-reactivity between this virus and TBEV, a flavivirus that is prevalent in Sweden. In contrast, seropositivity for WNV among Swedish donors may also be the result of tourism to southern Europe and or to United States, where WNV is endemic (Hernandez-Triana et al. 2014). Seropositivity to CHIKV in Sweden could also be explained by tourism, or by serological cross-reactivity to other alphaviruses. For instance, another mosquito-borne alphavirus, Sindbis virus, is endemic in parts of Central and Northern Sweden (Lundstrom et al. 1991, Gylfe et al. 2018) and it is important to address alphavirus cross-reactivity in future studies.
Furthermore, the high seroreactivity to CHIKV in Rwanda might be explained by cross-reactivity to ONNV, one of the endemic alphaviruses in sub-Saharan Africa and a close relative to CHIKV (LaBeaud et al. 2015), as demonstrated by our PRNT results for CHIKV and ONNV. Taken together, although PRNT titers from the two alphavirus assays are not directly comparable, these results may indicate seroprevalence of either one or both of these viruses in Rwanda. This question will be addressed in future studies.
As demonstrated in this study, the high degree of cross-reactivity between IgG antibodies against viruses belonging to closely related families, such as WNV and TBEV among flaviviruses and CHIKV and ONNV among alphaviruses, poses major problems for serological testing (Sanchini et al. 2013). Chabierski et al. (2014) produced a recombinant envelope protein with mutations in the conserved fusion loop to discriminate WNV serologically from other flaviviruses. In addition, Kam et al. (2015) described tests for anti-E2EP3 antibodies with the aim to differentiate CHIKV from other alphaviruses (and also from coinfection with flaviviruses) with similar clinical manifestations.
In contrast to our findings of probable cross-reactivity within the two virus families of alphaviruses and flaviviruses, the potential cross-reactivity between CHIKV and WNV was only 7.3% among all samples combined from Rwanda and Sweden. When considering each country separately, Rwanda accounted for 8.3% and Sweden for 2.3% of samples simultaneously seropositive for both CHIKV and WNV. The higher dual reactivity seen in Rwandan samples may reflect dual exposure to both CHIKV and WNV, as was also suggested by the highest prevalence of antibodies to both these viruses, and of their mosquito vectors in the Eastern Province (Tables 1 and 3). Since some Rwandan samples that tested positive for WNV also demonstrated low–or-moderate OD values for TBEV IgG, the cross-reactivity between these two flaviviruses will be further explored in a separate study.
To date, no actual clinical cases of CHIKV and/or WNV, nor any serological data concerning the epidemiology of these two viruses, has been reported from Rwanda. However, we assume that its inhabitants have been infected by these two viruses, especially since there are similarities in climate and ecological environment between Rwanda and its neighboring countries where these two viruses have previously been reported (CDC 2015). Furthermore, Kading et al. (2013) described WNV infection in gorillas sampled close to the Democratic Republic of the Congo border and suggested that these animals might have been infected in Rwanda.
In conclusion, we provide evidence that CHIKV and WNV are both circulating in Rwanda, as shown by the seroreactivity to both viruses, especially in the Eastern Province where their mosquito vectors were most common. Consequently, further studies on these two viruses in Rwanda are needed with regard to both vector competence and the clinical burden for health care institutions. Moreover, the serological cross-reactivity among alphaviruses and flaviviruses in Rwanda and in Sweden demonstrated by our study should be further elucidated.
Footnotes
Acknowledgments
The authors thank Maria Johansson working at the Virology Laboratory, Sahlgrenska University Hospital, for her technical assistance. The authors extend their gratitude to Janvier Sendarasi, who made the map of Rwanda showing the geographical distribution of IgG results for CHIKV and of adult Aedes mosquitoes. The LUA-ALF foundation of Sahlgrenska University Hospital, Gothenburg, Sweden (grant no. ALF-Gbg 145-841), the Swedish International Development Cooperation Agency: University of Rwanda–Sweden Program for Research, Higher Education and Institutional Advancement, and the Swedish Research Council (grant no. 2017-05607) provided financial support for this project.
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.