
Abstract
Salmonella enterica is a well-adapted zoonotic bacterium associated to cases of gastroenteritis and bacteremia with increased morbidity and mortality. In this study, three isolates of Salmonella Typhimurium obtained from human clinical samples, showing colistin resistance and low-level resistance to quinolones, have been genetically characterized. We detected the co-occurrence of mcr-1 and qnrS1 on a single IncHI2 plasmid in isolates of Salmonella Typhimurium obtained from Spanish children without a travel history. The multiresistant region contained numerous resistance genes. Isolates were clonally related, which suggests the presence of these clones in the community and the potential to cause outbreaks affecting the most susceptible population. It is necessary to monitor the presence of these plasmid-mediated resistance genes in human European strains of Salmonella spp. because of the risk of producing outbreaks of community-acquired infections.
Introduction
S
In this study, three isolates of S. enterica obtained from human clinical samples, showing colistin resistance and low-level resistance to quinolones, have been genetically characterized, as well as the plasmids carrying determining genes detected.
Materials and Methods
During the period between August and October 2015, three isolates of S. enterica showing co-resistance to fluoroquinolones and colistin were recovered from three patients with gastroenteritis. Patients were attended in two different primary health care centers in San Fernando (Cadiz, Spain). The identification and antimicrobial susceptibility testing were performed using Micro Scan ID/AST conventional panel in a MicroScan WalkAway System (Beckman Coulter, Brea. CA) using European Committe on Antimicrobial Susceptibility Testing (EUCAST) break points. In addition, quinolones susceptibility test was carried out by diffusion method using pefloxacin disk of 5 μg (Becton Dickinson, Franklin Lakes, NJ), ciprofloxacin minimum inhibitory concentration (MIC) Test Strips (Liofilchem, Roseto degli Abruzzi, Italy), and nalidixic acid disk of 30 μg (Becton Dickinson) according to EUCAST guidelines.
Pulsed-field gel electrophoresis (PFGE) analysis of XbaI-digested DNA (
Specific PCRs were performed to detect PMQR genes [qnrA, qnrB, qnrC, qnrD, qnrS, qepA, oqxAB, and aac(6′)-Ib-cr] and mcr-1 and mcr-2 genes as previously described (Liu et al. 2016, Rodríguez-Martínez et al. 2016). Positive and negative controls were included.
Plasmid DNA was extracted using the Kieser (1984) method and transformed by electroporation into E. coli DH10B (streptomycin-resistant). Plasmids, 154, 66, 48, and 7 kb from E. coli (EC50192) were used as molecular size markers. Transformants were selected on Luria-Bertani (LB) agar plates (Becton Dickinson France SA, Le Pont de Claix, France) supplemented with 100 mg/L ampicillin, 0.03 mg/L ciprofloxacin, 0.06 mg/L ciprofloxacin, 0.5 mg/L colistin, 1 mg/L colistin, or 2 mg/L colistin. Only transformants obtained from agar plates supplemented with 0.03 mg/L ciprofloxacin were selected for next assays.
The presence of PMQR and mcr genes in transformants was confirmed by genotypic methods, as described above. Plasmid DNA from the transformants was analyzed using restriction of S1 nuclease to establish the size of the plasmid (López-Cerero et al. 2011). Antimicrobial susceptibility testing for the isolates and their transformants was tested by broth microdilution. E. coli ATCC25922 was used as control for the microdilution assays.
Whole-genome sequencing (WGS) was used for analysis of genomes of transformants 93.1 and 106.1 for isolates 93 and 106, respectively. Genomic DNA was extracted from transformants and sequenced on the MiSeq platform (Illumina, San Diego, CA), generating 300-bp paired-end reads. The library was prepared using the Nextera XT DNA library preparation kit (Illumina), involving DNA fragmentation, adaptor ligation, and amplification. Raw reads were quality filtered and assembled into contigs on the CLC genomics workbench 10.0. (CLC bio; QIAGEN, Madrid, Spain), using default software settings. An average coverage of 50 × was observed. Plasmid replicons were identified with the PlasmidFinder tool (
Results
The three isolates (named as 93, 95, and 106) were serotyped as Salmonella Typhimurium, and showed the same antimicrobial susceptibility pattern. Isolates were resistant to ampicillin (MIC >16 mg/L), trimethoprim-sulfamethoxazole (MIC >4/76 mg/L), tetracycline (MIC >8 mg/L), colistin (MIC >4 mg/L), gentamicin (MIC = 8 mg/L), and tobramycin (MIC >8 mg/L). Susceptibility to nalidixic acid (<16 mg/L) was confirmed by disk diffusion. Regarding fluoroquinolones, MICs were 1 mg/L for norfloxacin and 0.38 mg/L for ciprofloxacin. Screening using pefloxacin was reported as resistant.
PFGE showed that the isolates 93 and 95 were identical (>99% of similarity, no band of difference) and possibly related with isolate 106 (86% similarity, six bands of difference) (Fig. 1).
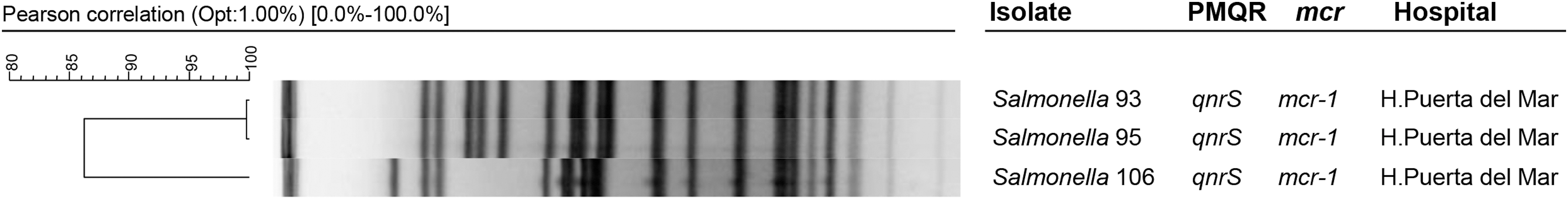
Pulsed-field gel electrophoresis and dendrogram analysis of mcr-1- and qnrS1-positive Salmonella Typhimurium isolates.
We detected qnrS1 and mcr-1 genes in all isolates. Transformants harbored single plasmids with both genes (Fig. 2). The size of plasmids was estimated as being between 175 and 208 kb. Positive results were confirmed by sequencing; MIC values are shown in the Table 1.
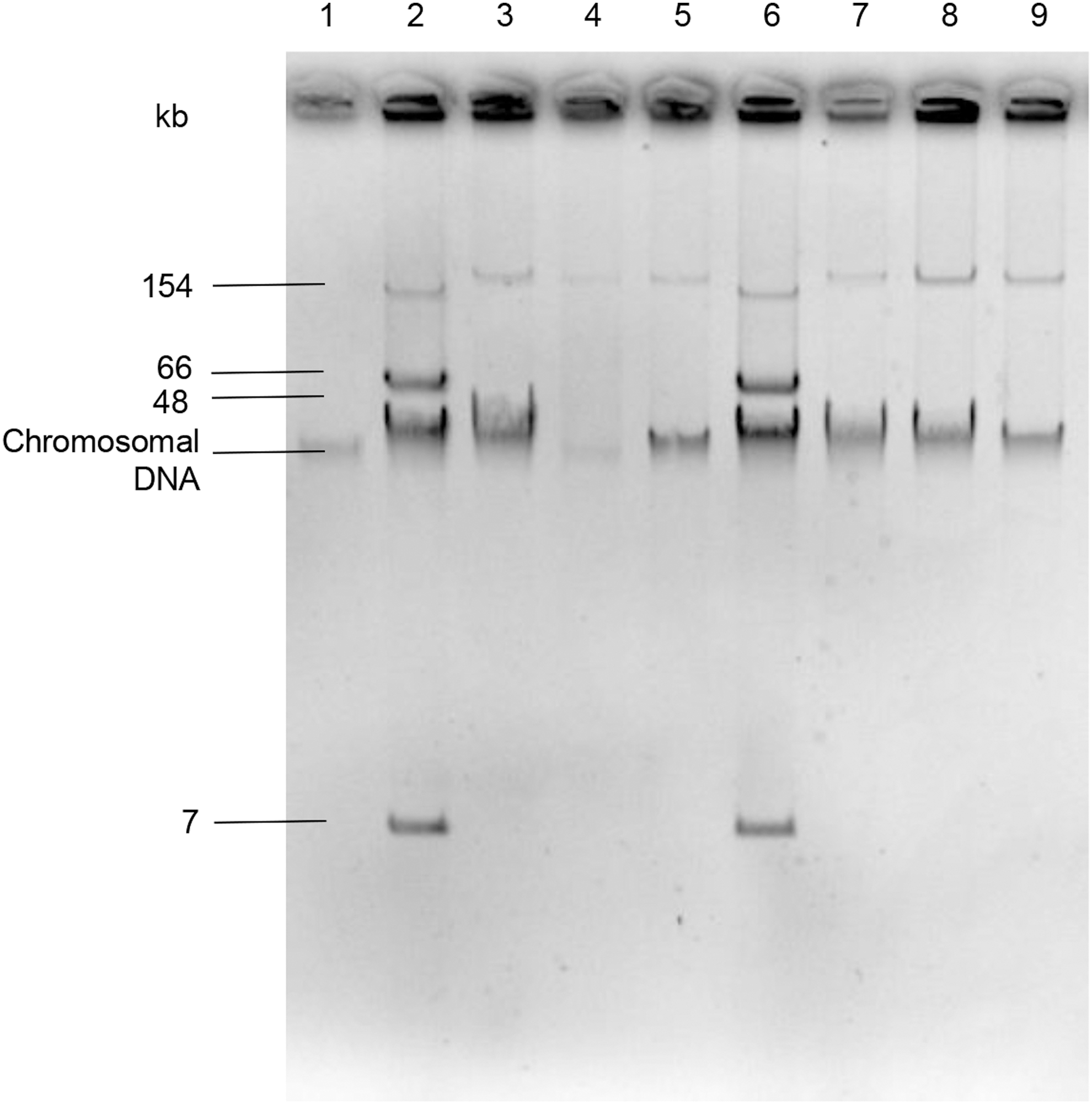
Plasmid profile of Salmonella Typhimurium isolates and transformants. 1. Escherichia coli DH10B; 2. E. coli 50192 (154-, 66-, 48- and 7 kb); 3. Salmonella Typhimurium 93; 4. Transformant 93.1; 5. Transformant 93.2; 6. E. coli 50192; 7. Salmonella Typhimurium 106; 8. Transformant 106.1; 9. Transformant 106.2.
Results of Susceptibility Testing of Escherichia coli DH10B, Salmonella Typhimurium 93, Salmonella Typhimurium 106, and Their Transformants
All MIC determinations were performed in broth microdilution assays according to CLSI standards.
MIC, minimum inhibitory concentration; CLSI, Clinical and Laboratory Standards Institute.
WGS assigned the plasmids to an IncHI2 group, and both plasmids were identical. The multiresistance region contained seven resistance genes, such as aph(4)-Ia, aac(3)-IVa, aadA2, bla-TEM-1b, sul3, dfrA12, and tetA. A schematic figure representing the genetic environment surrounding the mcr-1 gene is available as Supplementary Fig. S1.
Discussion
Several studies have shown the presence of PMQR and mcr-1 genes in Salmonella spp. isolates, mainly in food producing animals, but its co-occurrence is infrequent, even more the fact that both genes are located in the same plasmid. Wang et al. (2018) reported the detection of mcr-1 gene among ready-to-eat pork samples in China, where Salmonella Typhimurium plays an important role as a reservoir to disseminate multidrug resistance genes. In Spain, mcr-1 gene was detected on a plasmid highly mobilizable in Salmonella Typhimurium and other Enterobacteria isolated from poultry and swine (Quesada et al. 2016). In Italy, the prevalence of Salmonella isolates harboring mcr-1 from food-producing animals and food was higher than those from humans (Carnevali et al. 2016). A study carried out in Taiwan showed that mcr-1 was found in diverse S. enterica isolates causing infections in humans and food-producing animals (Chiou et al. 2017). A novel mcr-1 gene variant was reported in Salmonella Typhimurium isolate that also harbored PMQR genes such as oqxA and oqxB. Nevertheless, these PQMR genes were not carried in the same plasmid as mcr-1 gene variant (Lu et al. 2017). Plasmids belonging to IncHI2 group are predominant in Salmonella species and they are mainly responsible for the dissemination of antibiotic resistance in these bacteria (Chen et al. 2016). A study carried out in China revealed the presence of mcr-1 gene in 14.8% of Salmonella isolated from pigs at slaughter, most were serovar Typhimurium and the gene was located on IncHI2 plasmid (Yi et al. 2017).
In previous studies performed in E. coli isolates, mcr-1 gene was co-transmitted with other resistance determinants through IncHI2 plasmids, such as blaCTX-M, blaCMY, blaTEM, fosA, qnrS, floR, and oqxAB (Li et al. 2017). In Taiwan, mcr-1 gene found in several S. enterica isolates was carried on different plasmids and all the isolates, except for one, were multidrug resistant (Chiou et al. 2017).
We have only found two reports that describe the co-transmission of mcr-1 and qnrS1 in the same plasmid in six Salmonella isolates of human origin, although most patients had a travel history. These reports, from reference laboratories in United Kingdom and Denmark, analyzed colistin resistance in thousands of strains and only showed co-occurrence of mcr-1 and qnrS1 in Salmonella Typhimurium isolated from six patients. Five patients were adults, one patient was 8 years of age, and all patients, except one, had a recent history of travel to East Asia. Plasmids belonged to IncHI2 and IncI2 incompatibility groups (Doumith et al. 2016, Torpdahl et al. 2017).
Nevertheless, in our study, none of the three patients had history of travel, all of them were children, and two isolates were clonally related, which suggests that these clones are already circulating in our community, being able, in the same way as other Salmonella clones, to cause community outbreaks, affecting the most susceptible population. In our study, two patients have a family relationship, and isolation of Salmonella from fecal specimens was very close in time, which probably indicates exposure to a common source.
Conclusions
To our knowledge, this is the first report of co-occurrence of mcr1 and qnrS1 located in a single plasmid in Salmonella Typhimurium isolated from humans in Spain. It would be necessary to monitor the presence of resistance determinant genes to quinolones and colistin in Salmonella spp. of human origin as these clones are not only detected in food-producing animals but also could cause community-acquired infections and outbreaks in humans.
Sequence Accession Number
The Fastq files corresponding to transformant 93.1 and 106.1 with mcr-1 and qnrS1 bearing IncHI2 plasmids have been deposited in the GenBank database under accession number PRJNA510541.
Footnotes
Author Disclosure Statement
No conflicting financial interests exist.
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.