
Abstract
Our knowledge on the susceptibility of humans to different genotypes of the zoonotic tapeworm Echinococcus granulosus and the genetic make-up of the cysts in multi-organ involvements is limited. This study aimed to identify the genotype profile of E. granulosus in patients undergoing hydatid surgery in an endemic area for cystic echinococcosis (CE) in southeastern Iran. Individuals who underwent hydatid cyst surgery were included in this study. Protoscoleces/germinal layers from each individual isolate were washed and kept in −20°C until use. Genotyping was carried out using PCR-sequencing of two mitochondrial CO1 and ND1 genes. Molecular phylogeny and haplotype network analysis of the human isolates were carried out using sequence data obtained from this study and National Center for Biotechnology Information (NCBI) databases. Forty-two patients (23 women and 19 men) participated in the study; the mean age was 43 years. Eighteen (42.9%) and 24 (57.1%) patients were infected by E. granulosus sensu stricto (G1–G3) and Echinococcus intermedius (G6 genotype), respectively. Molecular study showed mixed infection of G1 (in the liver and right lung) and G6 (in left lung) in a patient. The study showed a significantly high proportion of CE patients infected with the G6 genotype particularly in the southern parts of the province. In the present study a human CE patient infected by two species/genotypes of E. granulosus sensu lato is documented.
Introduction
Cystic echinococcosis (CE) caused by the cestode Echinococcus granulosus sensu lato is one of the most medically and economically important of the zoonotic diseases prevalent in different parts of the world. Global monetary burden of CE due to E. granulosus sensu lato is estimated at US$ 2.1 billion annually, and it is believed to cost more than US$ 230 million per year in Iran (Fasihi Harandi et al. 2012, Deplazes et al. 2017). Therefore, control programs for zoonotic cestode infections are recommended by the World Health Organization (WHO 2015).
Significant intraspecific variations have been described within this species (Thompson, 2017). At least 10 different host-adapted genotypes have been identified within E. granulosus sensu lato. These genotypes include G1–G3 (E. granulosus sensu stricto, Kinkar et al. 2018), G4 (Echinococcus equinus), G5 (Echinococcus ortleppi, Thompson et al. 2002), Thompson and McManus, 2002 Toward a taxonomic revision), G6-G7 (Echinococcus intermedius), and G8/G10 (Echinococcus Canadensis, Laurimae et al. 2018a). Substantial genetic diversity within the parasite has considerable implications for the vaccine development, pathogenesis, diagnosis, and drug sensitivity (Eckert et al. 1988, McManus 2002, Thompson et al. 2002).
Among different genotypes/species of Echinococcus, humans are most susceptible to E. granulosus sensu stricto and most of the human CE cases are caused by this species (Rojas et al. 2014). However, it is believed that human being is refractory to the infection with E. equinus (G4 genotype) and less susceptibility of human has been expected for other genotypes/species (Thompson, 2017). It is estimated that E. granulosus sensu stricto (G1–G3) and E. intermedius (G6-G7) are responsible for 88% and 11% of global human CE infections, respectively (Rojas et al. 2014). Human infection by E. ortleppi (G5 genotype, cattle strain) has been reported from several geographical regions including The Netherlands, Argentina, Mexico, Brazil, France, South Africa, and India (Bowles et al. 1992, Kamenetzky et al. 2002, Deplazes et al. 2017).
Multi-organ hydatid disease has frequently occurred in humans, with the highest frequency of liver-lung infection. Involvement of the liver with one or more of other organs for example, spleen, kidney, and brain has been reported in the literature (Pawlowski 2001). It is estimated that in 20% of human CE, two or more organs are involved (Kern et al. 2017). Our knowledge on the genetic identity of the parasites involved in multi-organ/multi-cyst CE is limited. A couple of studies have been published on this issue in recent years (Oudni-M'rad et al. 2016, Debiaggi et al. 2017). A recent study in Argentina reported mixed G1/G7 genotype infection in a hepatic CE patient with three cysts (Debiaggi et al. 2017). Mixed cystic-alveolar echinococcoses and G1/G6 infection in dogs have been previously documented in Xinjiang, China (Bart et al. 2006, Zhang et al. 2006).
Our knowledge of the genetic make-up of hydatid cysts in multi-organ involvements is limited and the molecular epidemiological significance of mixed genotype infections in humans is not fully understood. Information on the species/genotype distribution of the parasites causing multiple CE infections is crucial in understanding disease transmission dynamics in different endemic regions of the world. To investigate Echinococcus species/genotypes involved in human CE, 50 cyst isolates from 42 human hydatid patients were studied in a localized geographical area using PCR-sequencing.
Materials and Methods
Study location and sample collection
Kerman province is situated in southeastern Iran, between latitude 26°29′ and 31°58′ N and longitude 54°20′ and 59°34′ E, with an arid and semi-arid climate, and a low annual rainfall that decreases toward the southeast. During 2-years period, from March 2011 to September 2013, a total of 50 samples of hydatid cysts from 42 patients were collected from Afzalipour Medical Center, the major referral hospital in the province. The patients agreed to give cyst samples with an informed consent. The cyst materials were transferred to the research laboratory of the Department of Parasitology, Kerman University of Medical Sciences.
All the cysts were examined to determine their fertility by microscopic observation of protoscoleces. For each fertile cyst, protoscoleces were washed several times with sterile solution of 0.9% sodium chloride followed by sedimentation in conical glassware at room temperature.
Molecular analysis
For DNA extraction ∼200 μL of the sediment or a piece of laminated/germinal layer was homogenized with an equal volume of distilled water. DNA extraction was carried out using High Pure PCR Template Preparation Kit (Roche, Mannheim, Germany) according to the manufacturer's instructions. Briefly, samples were incubated overnight in 200 μL tissue lysis buffer and 35–40 μg/mL proteinase K at 56°C. DNA extracted from protoscoleces was stored at −20°C until used in PCR amplification. Two primer pairs, JB3, 5′-TTTTTTGGGCATCCTGAGGTTTAT-3′ (forward), and JB4.5, 5′-TAAAGAAAGAACATAATGAAAATG-3′ (reverse) for CO1 gene (366 bp), and JB11, 5′-AGATTCGTAAGGGGCCTAATA-3′ (forward), and JB12, 5′-ACCACTAACTAATTCA CTTTC-3′ (reverse) for ND1 (444 bp) were used for PCR amplifications (Bowles et al. 1993).
The PCRs were carried out in 25 μL reaction volumes using Master Mix Red (Amplicon, Denmark) in Flexcycler thermal cycler (AnalyticJena, Jena, Germany). The thermal profile involved 10 min initial denaturation at 94°C, followed by 40 cycles, each of 30 s at 94°C, 60 s at 53°C, and 60 s at 72°C, and a final extension of 7 min at 72°C. The amplification products were subjected to electrophoresis on 1.5% agarose gel in TAE buffer.
Phylogenetic analysis
All the amplicons were sequenced on an ABI 3730XL capillary machine (Macrogen, Inc., South Korea). Sequence data were trimmed and adjusted manually and complete alignment was carried out using the softwares BioEdit (Hall 1999) and ClustalW (Thompson et al. 1997). Then, nucleotide sequences of CO1 and ND1 genes obtained from the present study were deposited in GenBank, National Center for Biotechnology Information (NCBI).
The evolutionary history was inferred by using the Maximum Likelihood method based on the kimura2 parameter model using MEGA5 software. Haplotypes defined by combination of single nucleotide polymorphisms within CO1 were assigned using DnaSP (Librado and Rozas 2009) and drawing of haplotype networks were performed by PopART software using statistical parsimony (Templeton et al. 1992, Leigh and Bryant 2015).
Results
A total of 50 hydatid samples were analyzed. Table 1 shows the frequency distribution of hydatid patients infected according to gender, age, and organs affected. The patients' age ranges between 4 and 74 years. Most of the patients (35.7%) were residents of Kerman, the capital of Kerman province. The lungs were the most frequently infected organ. Both right and left lungs were equally infected among pulmonary hydatid patients. Multi-organ involvements according to their genotypic identity have been summarized in Table 2. As shown in the table, one patient suffered from hydatid cysts in the liver and both lungs were diagnosed as having E. granulosus G6 genotype (E. intermedius) in the left lung and the G1 genotype (E. granulosus sensu stricto) in the liver and the right lung.
Frequency Distribution of Patients with Cystic Echinococcosis According to Gender, Age, Genotype, and the Involved Organ
Genotype Distribution of the Six Hydatid Patients with Multi-Organ Involvements from Kerman Province, Southeastern Iran
For all of E. granulosus isolates, fragments were successfully PCR-amplified within CO1 and ND1 genes. Alignments of the sequences determined herein with those of reference genotypes of E. granulosus sensu lato revealed the existence of genotype G1 and G6 in 18 (42.9%) and 24 (57.1%) respectively. Dendrograms generated by phylogeny are demonstrated in Fig. 1. Phylogenetic analysis revealed four haplotypes (Accession No. KY703621–KY703624) in the CO1 (three haplotypes for G1 and one haplotype for G6) and three haplotypes (Accession No. MF170142–MF170144) in ND1 gene (two haplotypes for G1 and one haplotype for G6). Figure 2 shows a haplotype network of CO1 diversity in representative human isolates from CE patients. Obviously, human isolates identified as E. granulosus sensu stricto exhibit higher nucleotide variations than E. intermedius.
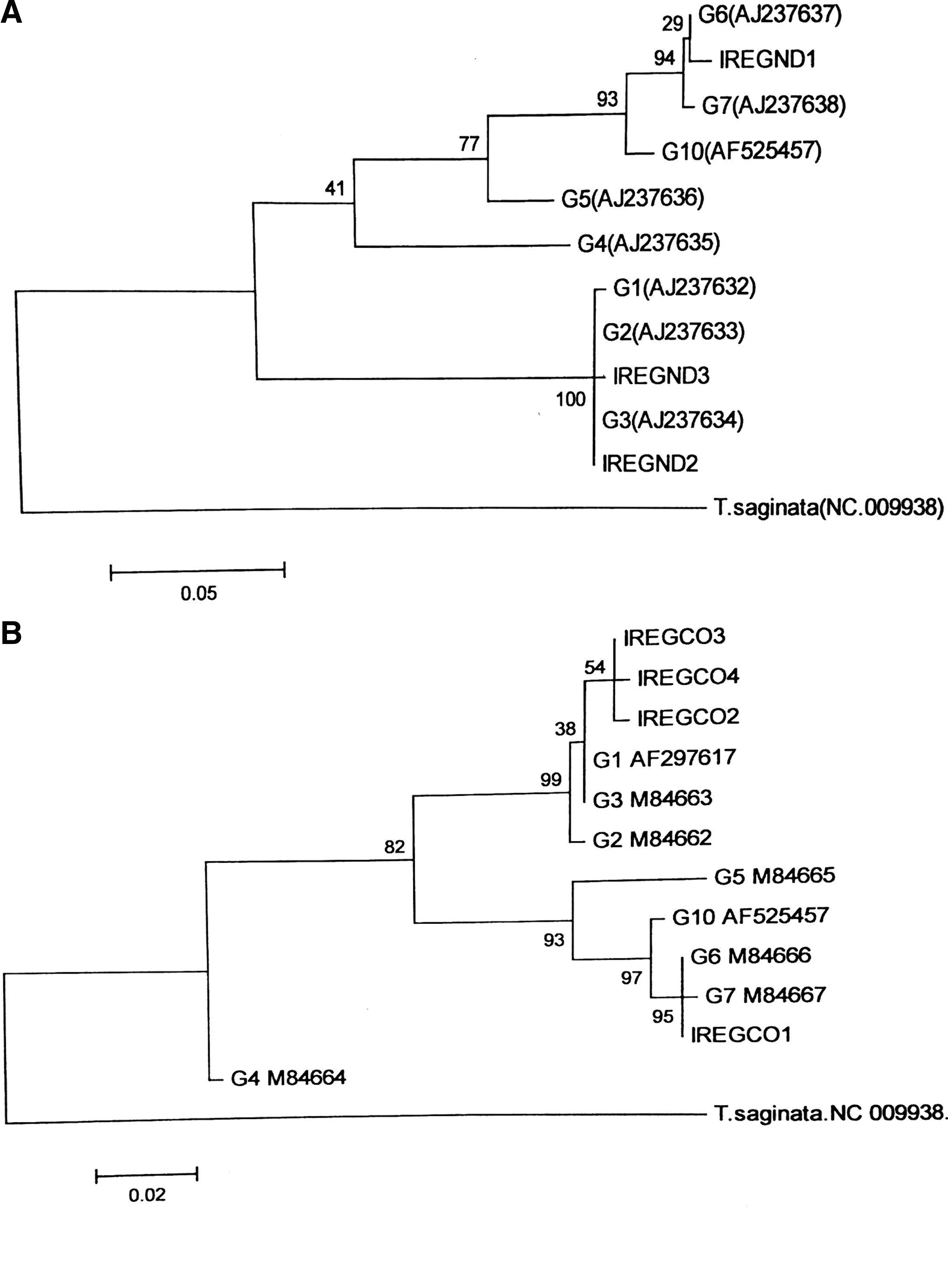
Molecular phylogenetic tree of Echinococcus granulosus isolates from human CE in 11 counties of Kerman province based on partial ND1

Network analysis of mitochondrial CO1 haplotypes found in human isolates of E. granulosus sensu lato based on statistical parsimony. The size of circles indicates the frequency of the haplotypes. Empty circles represent haplotypes detected in the current study (right panel: E. granulosus sensu stricto, left panel: E. intermedius). Solid circles show representative haplotypes of human origin found in NCBI GenBank. The circles are identified by the corresponding accession numbers (KY703622 and KY703623 are identical to DQ356877 and AF408686 respectively).
Discussion
This study aimed to identify the genotype composition and the significance of mixed infections with different species/genotypes within E. granulosus sensu lato as the main etiological agents of human CE in Iran. Table 3 summarizes the current information reported on the species/genotypes causing mixed Echinococcus infections in the world. In China and Kazakhstan simultaneous infection of dogs with E. granulosus and Echinococcus multilocularis has been previously documented (Štefanić et al. 2004, Zhang et al. 2006). Mixed-genotype infections have been also detected in the canine definitive hosts. Kamenetzky et al. described two different genotypes (G1and G6) in a single dog (Kamenetzky et al. 2002). Bart et al. documented a mixed G1 and G6 infection in a dog in Xinjiang, China (Bart et al. 2006).
Summary of the Current Knowledge on the Species/Genotypes Causing Mixed Echinococcus Infections in the World
In humans simultaneous alveolar and cystic echinococcoses have been reported from southern Ningxia Hui Autonomous Region in China where both AE and CE are endemic (Yang et al. 2006). Recent studies from Tunisia and Argentina described patients with mixed infections of G1/G7 and G1/G6 origins, respectively (Oudni-M'rad et al. 2016, Debiaggi et al. 2017). The present study describes the first documented human CE in the Middle East caused by two species/genotypes of E. granulosus sensu lato, that is, E. granulosus sensu stricto and E. intermedius. One out of six patients with multi-organ CE was diagnosed with a hepatic G1 and two pulmonary G1/G6 infections (Table 2). The both species/genotypes occur sympatrically in the province. As dogs have been shown to be infected by both G1 and G6 genotypes, G1 and G6 eggs could be readily dispersed in the environment and it is theoretically possible for humans to be infected with both genotypes. Different species/genotypes of E. granulosus sensu lato are believed to present different biological and epidemiological behaviors (Thompson, 2017). As the prepatent period in both genotypes is different in the definitive host, this issue will affect planning control programs. It has been shown that the prepatent period of G1 genotype is longer than G6 (Soriano et al. 2016). Therefore, this finding has important implications in the epidemiology and control of human CE.
The observed differences in the frequency of human infection with G1–G3 and G6 genotypes in the province may be explained by the following suppositions relevant to the parasite, host, and environment: (1) probable differences in infectivity to humans among different genotypes of E. granulosus sensu lato (Eckert et al. 1997), (2) potential differences in host genetics and immune system in response to Echinococcus infection (Lymbery et al. 1990), (3) probable differential fecundity and/or resistance of Echinococcus species/genotypes to the environmental conditions, and (4) pressure of certain genotypes/species of the parasite and the numerical abundance of some suitable hosts, for example, camels, well-adapted to the parasites perpetuating in the environment (Thompson et al. 1988, Harandi et al. 2002).
According to Alvarez-Rojas more than 11% of E. granulosus sensu lato isolates from human are due to G6/G7 genotype. However, in the present study more than 57% of the isolates were found as G6 genotype. This is especially peculiar because the proportion of G1 to G6 genotype increased southward from high altitude temperate regions of north to the low altitude hot areas of south. According to the latest national statistics out of 108,500 camels in the country, 12,000 (11%) camels are living in Kerman province. More than 60% of the camels are living in the arid/semi-arid areas in the southern parts of the province. According to a recent study from Iran on 280 isolates, G6 genotype was found in more than 71% of camel hydatid isolates. In total 192, 15, and 73 isolates were identified as G1, G3, and G6 genotypes. Sheep, cattle, and camel isolates were genotypically categorized as G1 (93%, 94%, and 25%), G3 (7%, 4%, and 4%), and G6 (0%, 2%, and 71%), respectively (Rostami et al. 2013).
Our knowledge on the probable differential fecundity and/or resistance of Echinococcus species/genotypes to environmental conditions is poor and more investigations on these issues are needed. It is not known whether the eggs of E. intermedius (G6 genotype) are less susceptible than G1 to unfavorable environments of the endemic areas in south Kerman. This study showed that G6 genotype is more prevalent than G1 in southern areas of the province. It has already shown that the phenotypes of the parasite could be influenced by host genetics (Lymbery et al. 1990). More in-depth studies on the mechanisms of the parasite pathogenesis and interactions between host and parasite are required to explain probable differences in infectivity to humans among different genotypes of E. granulosus sensu lato.
Aaty et al. (2012) found a significantly higher rate of human infection in Egypt caused by G6 genotype. Another study in Iran on 125 formalin-fixed paraffin-embedded blocks of human CE patients showed more than 40% of the isolates as G6 genotype (Rostami et al. 2015b). More investigations are required to clarify determinants of higher frequency of E. intermedius (G6-G7 genotype) in humans from certain geographical regions.
As shown in Fig. 1, CO1 gene presents a higher degree of variation than ND1. This is in agreement with the findings in other cestode parasites, for example, Taenia hydatigena, Taenia multiceps, and Taenia saginata (Campbell et al. 2006, Rostami et al. 2015a). In the present study G6 isolates were found as one haplotype (Fig. 2), however, large-scale mitogenome studies showed higher variations in the mitochondrial regions (Laurimae et al. 2018b). It should be noted that the isolates investigated in the present study were collected from a relatively localized geographical area with similar socioeconomic/climate conditions while in other studies the parasites were isolated from vast geographical localities across the country (Rostami et al. 2015b).
As revealed in Fig. 2, in human CE the intraspecific variations within E. granulosus sensu stricto are higher than that of E. intermedius (G6-G7 genotype). Wider range of intermediate host species, more intense transmission, and higher turnover of the parasite in sheep and longer lifespan of camels are among the main reasons of greater genetic diversity in E. granulosus sensu stricto. Further in-depth molecular epidemiological studies on human isolates, using several mitochondrial and nuclear loci, are required to improve our understanding of the nature of human CE agents in the world.
Conclusions
The study demonstrated concomitant hydatid cysts in human caused by two different species/genotype of E. granulosus sensu lato. This may lead us closer to the understanding of pathogenesis and host–parasite relationships in CE. Haplotype analysis, subgenotyping, and molecular investigations on the sources of contamination and the dynamics of transmission are of paramount importance and the literature is poor on this issue. More molecular studies are required to be carried out in this interesting field of CE research. The significantly higher proportion of patients with E. granulosus G6 genotype than the world average in certain regions of the Middle East should be considered in planning CE prevention and control programs.
Footnotes
Acknowledgment
The authors wish to thank Afzalipour Medical Center personnel for their assistance in the collection of parasite materials.
Author Disclosure Statement
No conflicting financial interests exist.