
Abstract
Vector-borne diseases are emerging at an increasing rate and comprise a disproportionate share of all emerging infectious diseases. Yet, the key ecological and evolutionary dimensions of vector-borne disease that facilitate their emergence have not been thoroughly explored. This study reviews and synthesizes the existing literature to explore global patterns of emerging vector-borne zoonotic diseases (VBZDs) under changing global conditions. We find that the vast majority of emerging VBZDs are transmitted by ticks (Ixodidae) and mosquitoes (Culicidae) and the pathogens transmitted are dominated by Rickettsiaceae bacteria and RNA viruses (Flaviviridae, Bunyaviridae, and Togaviridae). The most common potential driver of these emerging zoonoses is land use change, but for many diseases, the driver is unknown, revealing a critical research gap. While most reported VBZDs are emerging in the northern latitudes, after correcting for sampling bias, Africa is clearly a region with the greatest share of emerging VBZD. We highlight critical gaps in our understanding of VBZD emergence and emphasize the importance of interdisciplinary research and consideration of deeper evolutionary processes to improve our capacity for anticipating where and how such diseases have and will continue to emerge.
Introduction
The threat of emerging infectious diseases to humans and wildlife is a pressing issue that is expected to worsen in response to human population growth and global change (Gratz 1999, Morens et al. 2004, Patz et al. 2008, Keesing et al. 2010, Kilpatrick and Randolph 2012). In 2015, mosquito-borne Zika virus emerged in South and Central America followed by mosquito-borne transmission in Southern Florida and at the Texas/Mexico border in the United States (Belluck and Alvarez 2016). Widespread Zika virus transmission, along with the new recognition of congenital Zika syndrome that causes fetal death or neurologic defects in babies born to women infected during pregnancy, has brought vector-borne pathogens to the forefront of public attention. Over 60% of emerging human diseases are zoonotic, or caused by microorganisms that are transmitted from wildlife to humans (Jones et al. 2008). Vector-borne zoonotic diseases (VBZDs), transmitted primarily by blood-feeding, or hematophagous, and arthropods, are an important subset of zoonotic diseases that are emerging at an increasing rate relative to directly transmitted infectious diseases that lack an arthropod intermediary. The most recent compilation of data documenting that a high number of emerging infectious diseases are vector borne is over 15 years old (Jones et al. 2008). Specifically, although 14% of human infectious diseases are vector borne, VBZDs comprise 22% of all emerging infectious diseases of humans and are thus disproportionately represented among emerging infectious diseases (Cleaveland et al. 2001, Taylor et al. 2001, Woolhouse and Gowtage-Sequeria 2005, Vorou et al. 2007, Jones et al. 2008). Another recent report by the Centers for Disease Control (CDC) found that the number of reported cases of tick-borne diseases has doubled in recent years in the United States, mostly due to increased cases of Lyme disease, but also due to other diseases increasing or because new pathogens have been described (Rosenberg et al. 2018). The other major pathogens causing human infectious diseases consist of directly transmitted viruses, prions, bacteria, and parasites (Cleaveland et al. 2001, Jones et al. 2008, Keesing et al. 2010).
Despite recognition of VBZDs as a major group of emerging diseases, there has not been a systematic characterization of the most important vectors, pathogens, environmental contexts, or ecological and evolutionary features that promote their emergence. This study synthesizes reported VBZD incidences for multiple pathogens and vectors to identify patterns of emergence to ask when and where vectored pathogens have emerged and what common factors facilitate their emergence? The emergence of new or expanding vector-borne pathogens in humans likely reflects common ecological and evolutionary processes rather than isolated phenomena. We conducted a literature review to assess VBZD emergence and discuss key ecological and evolutionary characteristics of vector-borne diseases in contrast to nonvectored human pathogens to develop a conceptual framework for understanding why vector-borne diseases are disproportionately represented among global emerging infectious disease events.
Methods
We followed the CDC definition of emerging infectious disease: a disease that has increased in incidence because it is newly described, recently expanded in distribution, or increased in prevalence. The definition of a vector can differ between fields and contexts (Wilson et al. 2018), but here, because we seek to understand evolutionary drivers and associations, we defined a vector as a blood-feeding arthropod in which a pathogen, once ingested with a vertebrate blood meal, replicates and is ultimately transmitted in a subsequent blood meal. This is in contrast to pathogens that might be mechanically transferred from one host to another on the exterior surfaces of the arthropod's body (Wilson et al. 2018). This review focuses on arthropods because they are the most common and well-characterized vectors of pathogens. Major groups of arthropod vectors include insects such as mosquitoes, fleas, and kissing bugs, as well as arachnids such as ticks. There are nonarthropod animals that engage in hematophagy, such as leeches, vampire bats, and even vampire finches (Schluter and Grant 1984), but these examples are unusual and their importance in pathogen transmission is limited, so they are not included in this analysis.
Literature review of emerging VBZDs
Standardized database searches were conducted on BIOSIS Citation Index and Web of Science on July 10, 2018, and included the search terms “emerging AND infectious AND disease AND vector” in the topic field and the year range was 1940–2018. A disease that emerged more than once or in multiple locations was only counted once, but separate emergence locations were recorded. We explored spatial trends in vector-borne disease emergence by comparing emergence events by continent and by latitude. For the continent-level analysis, all 131 pathogens obtained from the BIOSIS and Web of Science searches were included, and the continent of emergence for each pathogen was determined from information provided in the relevant articles. Pathogens emerging simultaneously on multiple continents (e.g., Borrelia miyamotoi) were included in the count for each of those continents (e.g., North America and Europe).
We then sought to control for differences in surveillance effort by estimating each continents' capacity for detecting emerging vector-borne pathogens. We used the number of field entomological articles published in each continent as a proxy for relative surveillance effort. Specifically, we conducted a Web of Science search on the topic “field entomology” between 1940 and 2018, which generated a list of 348 articles. For each article, we then determined the continent that the research occurred, rather than the home continent of the researchers (as reported by Web of Science), and tallied the number of studies per continent. We then divided the number of emerging vector-borne pathogens per continent by the number of field entomological articles per continent to obtain scaled emergence event values.
For the emergence-by-latitude analysis, we included all vector-borne pathogens with known emergence locations at the country level or finer, as determined by the available literature. This included 76 of the original 131 emerging vector-borne pathogens identified. For these pathogens, the mid-point latitude value of the reported emergence location was used in the analysis. For example, Anaplasma phagocytophilum, which was reported to emerge in Duluth, Minnesota (Bakken et al. 1994), would be assigned the middle latitude value for Duluth (roughly 46°N).
Drivers of emergence
To quantitatively assess factors that may have promoted a particular emerging VBZD, we recorded drivers that were identified by the authors in the primary studies we reviewed. In the numerous cases where no driver was indicated or proposed, we recorded these diseases as “unspecified.”
Results
Major patterns of emerging vector-borne diseases
Our analysis identified 131 emerging vector-borne diseases in the peer-reviewed scientific literature from the years 1940 to 2018 (Supplementary Table S1). The BIOSIS query resulted in 1020 results and Web of Science produced 836 results. All peer-reviewed journal articles were reviewed to determine if the article was about an emerging vector-borne disease (Supplementary Table S1). We identified 53 emerging vector-borne diseases since 2005, in addition to the 78 that were originally described by Jones et al. (2008). New genetic variants of pathogens that evolved to infect a new vector or host were considered separate pathogen emergence events in our analysis.
Our analysis found that acari (mites and ticks) and diptera (flies) comprise nearly 90% of all vectors involved in emerging VBZD (Fig. 1). Within these groups, the vectors most commonly involved in transmitting VBZD are dominated by hard ticks (family Ixodidae) and mosquitoes (family Culicidae), respectively, and are responsible for vectoring approximately three-quarters of all documented emerging VBZD. Ixodidae ticks transmit 40% of documented emerging VBZD and mosquito vector 36% (Supplementary Table S1 and Fig. 1). Examples of Ixodidae are Ixodes, Dermacentor, or Amblyomma spp. ticks that are responsible for transmitting some of the most prevalent and widespread bacterial infections to humans such as Lyme disease, Rocky Mountain spotted fever, and ehrlichiosis. Mosquito species involved in the majority of mosquito-borne diseases are primarily in the genera Aedes, Anopheles, or Culex and are well-known vectors of West Nile virus, malaria, and Zika virus. The remaining emerging VBZDs are principally vectored by fleas, sand flies, and phlebotomine flies (Fig. 1).
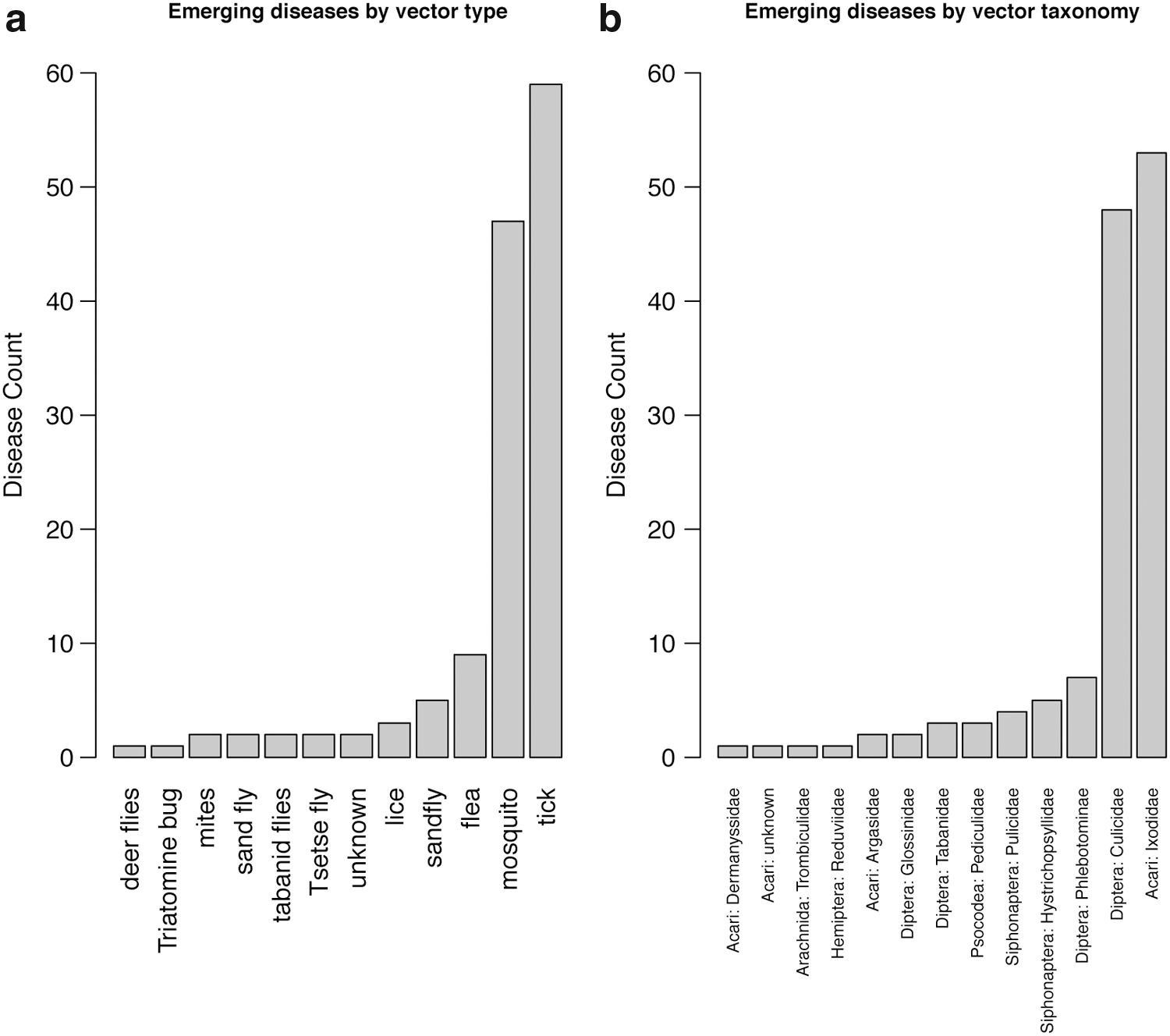
Histogram showing the
The pathogens transmitted by the two most prominent vector families, Ixodidae ticks and Culicidae mosquitoes, are dominated by Rickettsiaceae (N = 37) and a variety of viruses, including Flaviviridae, Bunyaviridae, Togaviridae, and rarely Reoviridae (N = 49). The remaining vector-borne pathogens are bacterial (N = 23), protozoan (N = 9), and a variety of other pathogens such as nematode worms and flukes (Supplementary Table S1 and Fig. 2b). Of the emerging pathogens transmitted by arthropod vectors, bacteria are responsible for 59% of all VBZDs, followed by viruses (27%), and then protozoa (15%) (Fig. 2). When examining the types of emerging pathogens at the family level, the results showed that Rickettsiaceae bacteria are the most common family of pathogens by a large margin and account for over a quarter of all emerging pathogens at the family level.
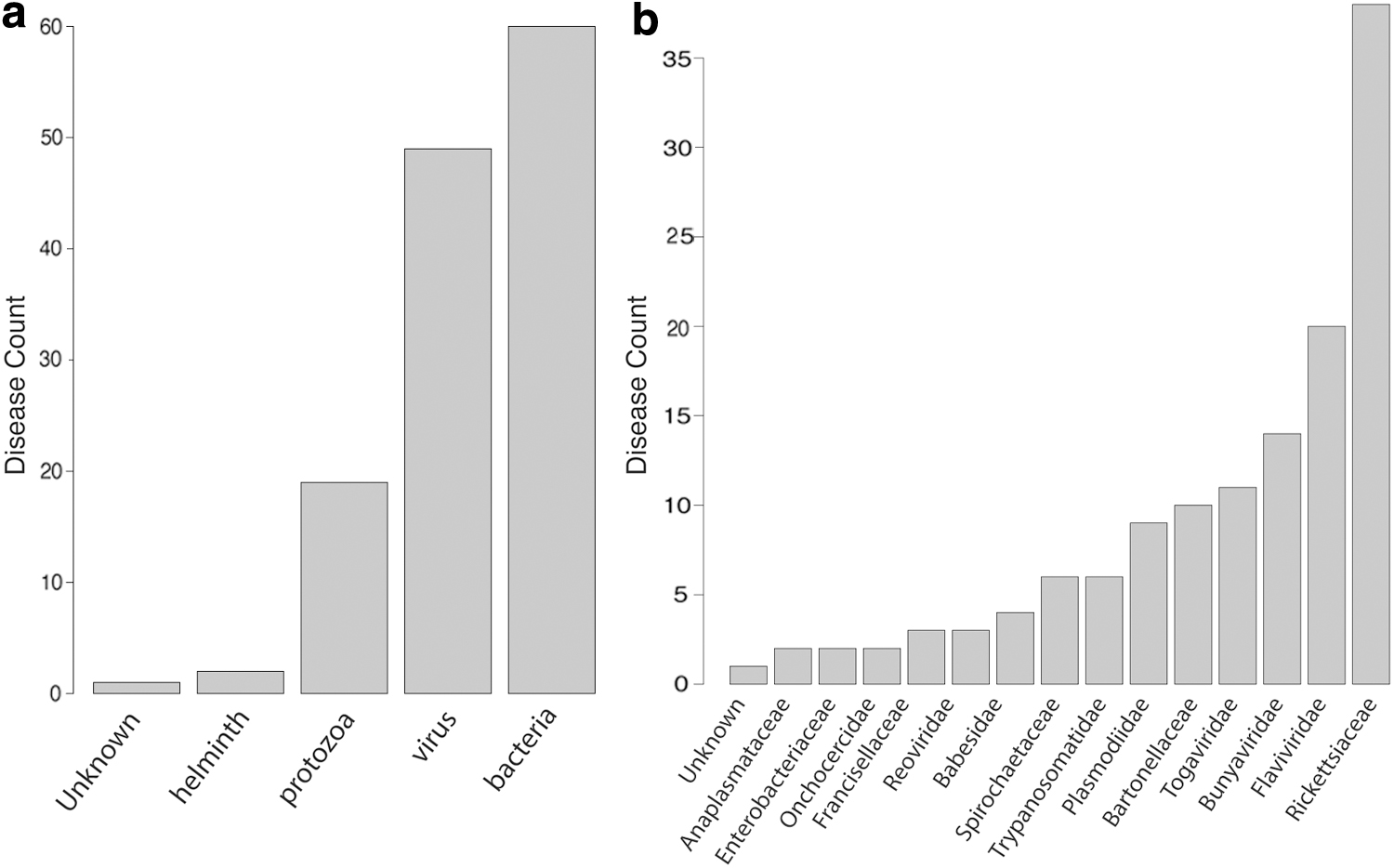
Count of disease types categorized by
Spatial patterns of emergence
We found the highest number of vector-borne pathogen emergence events in North America (27%) followed by Europe (21%) and Asia (20%) (Fig. 3). However, as unequal surveillance efforts across continents may influence the detection of emergence events, we repeated this analysis after scaling the number of emerging pathogens using a proxy measure of surveillance effort for each continent by accounting for the relative number of field entomology publications from each region. Using this correction, we found Africa hosted substantially more emerging vector-borne pathogens (43%) than any other continent, and Australia also hosts a relatively larger share, while North America and Europe's contribution becomes smaller, perhaps due to sampling bias (Fig. 3). Analyzing vector-borne disease emergence patterns by latitude, in another effort to minimize surveillance bias, revealed that emergence events were highest at both the mid-latitudes (45°N) and the tropics (15°S), supporting both the scaled and unscaled results from above (Fig. 4).

Histogram showing locations of emerging vector-borne pathogens by continent

Emerging vector-borne disease events shown by latitude.
Potential drivers of emergence
Our analyses found that, while climatic and weather-related factors are explicitly implicated in the emergence of some VBZDs, these are not the most commonly noted reasons for emergence (Fig. 5). Instead, our analysis found that land use changes were the most commonly cited driver for VBZD emergence, accounting for 26% of all VBZD diseases (Fig. 5). The second largest category was unspecified or unknown drivers of emergence (14%). Following as the third most important category was international trade and commerce (11%), reflecting the way that increasing globalization has moved people, vectors, and pathogens into ever closer contact (Fig. 5). Climate and weather-related factors are ranked fourth as a driver of emerging VBZDs and account for 10% of identified VBZDs in this study.
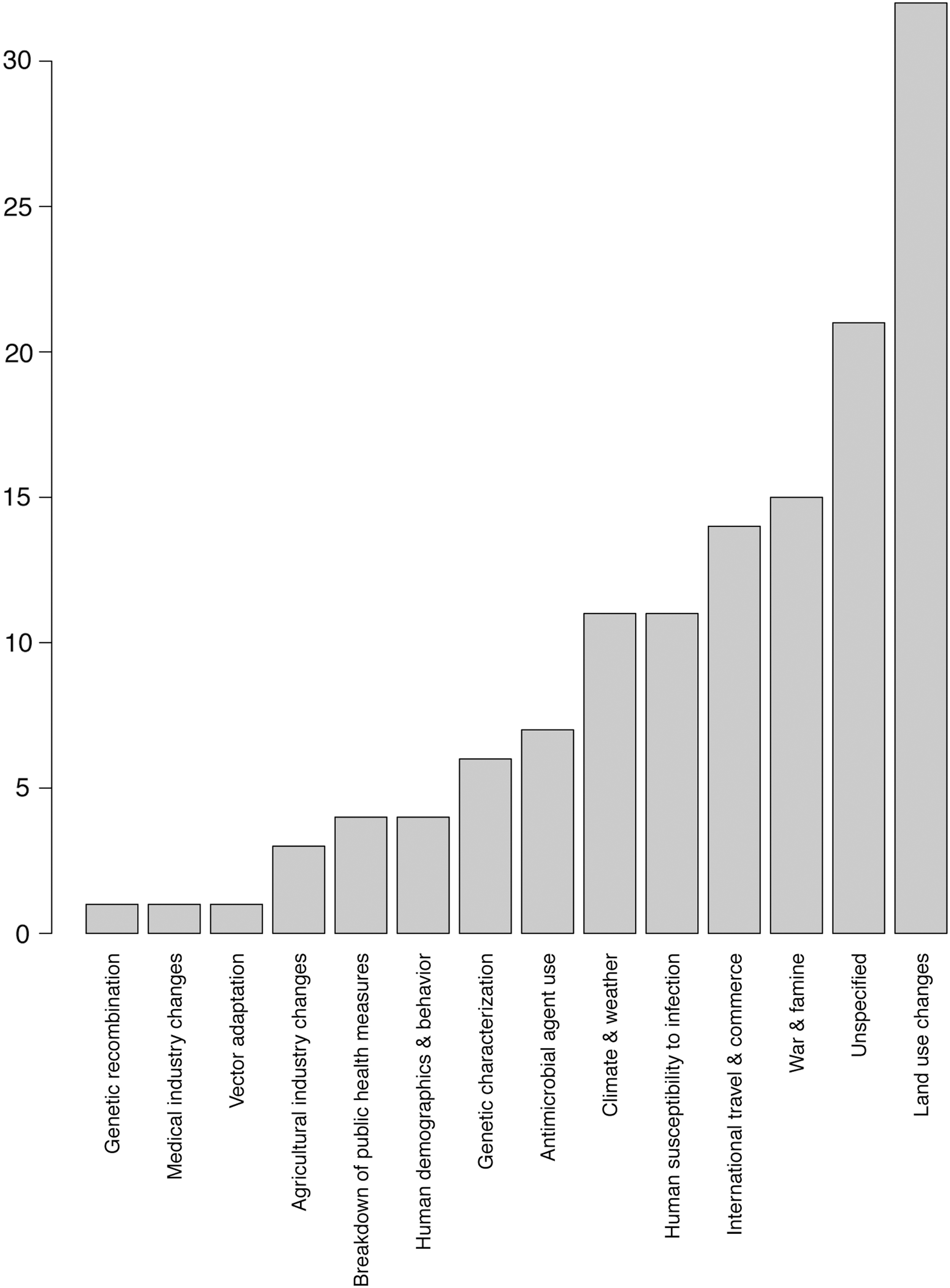
Putative drivers of vector-borne disease emergence as specified by primary literature authors.
Discussion
Our literature review revealed spatial and pathogen-related patterns and trends in vector-borne disease emergence and provides much needed quantitative analysis of characteristics and drivers of emerging vector-borne diseases. Ticks (family: Ixodidae) and mosquitoes (family: Culicidae) are the leading vectors of emerging VBZD, accounting for over 90% of documented instances of emerging VBZDs. Emerging VBZDs were largely found to be bacteria in the family Rickettsiaceae and viruses in the family Flaviviridae. Furthermore, Rickettsiaceae are more than two times more likely to be agents of an emerging VBZD than the next two most common pathogens, the viral families Bunyaviridae and Flaviviridae, each comprising 11% of all emerging VBZDs. The Rickettsiaceae are always transmitted by ticks (Tomassone et al. 2018), whereas Bunyaviridae and Flaviviridae are predominantly transmitted by Aedes or Culex mosquitoes, although some members are transmitted by hard ticks (van der Weijden et al. 2007, Yu et al. 2011, Wormser and Pritt 2015). Nearly half of all the emerging VBZDs (47%) are attributed to these agents, and suggest important aspects of their biology, evolutionary history, and transmission ecologies that facilitate their spread and emergence.
The role of vector type in the emergence potential of pathogens
Ticks and mosquitoes are the most common zoonotic disease vectors (Fig. 1). Their extremely high reproductive output under suitable conditions can contribute to high levels of transmission when pathogens are present. Certain mosquito vectors within the Aedes and Culex genera are invasive and highly successful in disturbed environments, thriving particularly well in peridomestic landscapes where human and domesticated animal blood meals are readily available (Juliano and Lounibos 2005, Medlock et al. 2012). Aedes species that vector dengue, Zika, and chikungunya viruses have adapted to breed in artificial water containers abundant in human communities. These ecological features contribute to effective and potentially large-scale transmission among humans.
Mosquitoes and ticks can support pathogens at high levels within their bodies and transmit them throughout the course of their lives. However, different feeding modes, continuous in the case of mosquitoes, and discrete and limited in the case of hard ticks, can impact transmission dynamics. Continuous feeding vectors take numerous blood meals and can transmit pathogens at any of these events. Limited-feeding hard ticks take only a single blood meal per life stage, of which they have three: larvae, nymph, and adult. In the latter case, vector competence and pathogen transmission depend on the persistence of the pathogen through the transstadial molt from one cohort to the next (e.g., from fed larvae to molted nymph). Co-feeding (Hudson et al. 2001, Beugnet and Marie 2009), long-lived vertebrate reservoir hosts (Kilpatrick et al. 2017), or asynchronous activity of life stages (Kurtenbach et al. 2006) are mechanisms for overcoming these life stage barriers at the population and community level in tick-borne diseases. A pattern that emerges from this analysis is that ticks primarily transmit emerging bacterial diseases (N = 44/54) and viral pathogens to a lesser degree, whereas mosquitoes only transmit viral pathogens (Supplementary Table S1). The ability of ticks to transmit multiple types of emerging pathogens may be partially responsible for their prominence as a vector of emerging zoonotic diseases.
Spatial patterns of emerging VBZD
Our analysis of spatial patterns of VBZD emergence found that North America and Europe are associated with the highest numbers of emerged VBZD over the last several decades. These patterns have been reported by others (Woolhouse and Gowtage-Sequeria, 2005, Jones et al. 2008), but minimized because of the high likelihood of surveillance bias. However, we conducted an analysis of emergence events based on latitude, an approach that is less prone to sampling bias, and found that temperate latitudes around 45°N have the highest number of emerging VBZD. Furthermore, we attempted to account for sampling bias by scaling our results by the number of field entomology publications in the databases we used (BIOSIS and Web of Science). After correcting for spatial reporting biases, we find that Africa has likely experienced more than double the number of emergence events than any other continent. In the case of dengue, Africa was found to be particularly at risk for transmission due to high levels of precipitation and temperature suitability for dengue, as well as abundant and highly connected low-income urban and peri-urban centers to facilitate human movement (Bhatt et al. 2013). In addition, weaker surveillance systems, combined with the presence of ecological drivers of emergence such as rapid urbanization and relatively high wildlife species richness (Jones et al. 2008), position Africa to be particularly vulnerable to new and resurging vector-borne disease epidemics. In contrast, while the ecological drivers associated with vector-borne disease emergence are present in North America and Europe, these continents are better equipped to contain large-scale outbreaks due to greater surveillance, infectious disease research efforts, and health infrastructure (Jones et al. 2008).
Potential drivers of emergence
At the root of all global change is the unparalleled influence of human population growth on ecological processes. While the topic of vector-borne disease emergence, especially malaria, has been well covered elsewhere with respect to climate change (Rogers and Randolph 2006, Richert-Poeggeler et al. 2008, Paaijmans et al. 2009, Proestos et al. 2015, Lafferty and Mordecai 2016), less attention has focused on other aspects of anthropogenic-driven global change. For instance, globalization and human movement far outpace the rate of climate change and is directly responsible for the circumglobal spread of several emerging VBZDs, including chikungunya and dengue viruses (Mackenzie et al. 2004, Wilder-Smith and Gubler 2008, Bennett 2014, Musso et al. 2015).
Dengue virus is a prime example of how the interaction between rapid pathogen evolution, human movement, and changing vector ecology has driven emergence. Dengue virus, which comprises four genetic and serologically distinct lineages, originated from zoonotic transmission amongst non-human primate hosts in tropical forest canopies, and emerged as the serotypes adapted to human hosts and vectors in the genus Aedes (Bennett 2010, Wang et al 2000). Following the domestication and spread of its main vector Ae. aegypti, and later an increasingly important secondary vector Ae. albopictus, a global transmission arena was born, characterized by ongoing emergence in humans (Wilder-Smith and Gubler 2008, Lambrechts, Scott and Gubler 2010).
The movement of humans on all scales remains one of the main drivers of dengue resurgence. For example, across the Pacific, following the establishment of Ae. aegypti, travel of infected humans spread dengue virus, leading to widespread epidemics in the 1970s (Effler et al. 2005; Steel et al. 2010). Dengue continues to spark outbreaks in Hawaii (Effler et al. 2005) where both Ae. aegypti and Ae. albopictus persist after they became established in the 1890s (Winchester & Kapan, 2013). At the regional scale, in Thailand seasonal movement brings dengue into Bangkok from outlying provinces (Cummings et al. 2004). At a city-wide scale, Singapore's 2006 epidemic was fueled by human movement where patches of mosquitoes in public spaces became infected and subsequently spread disease to uninfected hosts (Adams & Kapan 2009, Ooi et al. 2006).
Land use change as a prominent driver of VBZD emergence may explain the high prevalence of emerged VBZD in North America and Europe. Extensive land use change in these continents, most notably anthropogenic fragmentation in Europe (Wade et al. 2003), reforestation in the northeastern United States and deforestation in Africa (Cronon 1983, Stanek et al. 2002, Vijay et al. 2016), and urbanization across North America and Europe (Chen et al. 2014) provide increasing opportunities for contact between people and potential disease vectors and vector-borne disease reservoirs (Patz et al. 2008). Furthermore, warming in these mid-latitude regions appears to be increasing the geographic range and suitable season length for many arthropod species (Greer et al. 2008, Butterworth et al. 2017), whereas warming in tropical regions is resulting in shifts in arthropod range rather than overall increases (Lafferty and Mordecai 2016).
The emergence of Lyme disease, the most prevalent vector-borne disease in the Northern Hemisphere with an estimated 300,000 cases annually, exemplifies the way anthropogenic land use change can lead to emergence. Genetic studies estimate that the Lyme disease causing bacterium, B. burgdorferi, has been in North America for hundreds of thousands to millions of years (Hoen et al. 2009, Margos et al. 2011), but only emerged as a zoonotic pathogen in the early 1980s due to deforestation and secondary growth in the northeastern United States, which led to increased abundances of ticks and their hosts, particularly white-footed mice and deer (Barbour and Fish 1993). In this case, landscape change altered host-vector contact opportunities and land use patterns put humans in more contact with the vectors as well.
Due to the importance of abiotic limits to arthropod vectors, the emergence of VBZDs is often attributed to changes in climate and weather (Gubler et al. 2001, Ogden et al. 2006, Nakazawa et al. 2007, Medlock et al. 2013). Changing climatic conditions have been associated with changes in vector range (Ogden et al. 2006, Medlock et al. 2013), phenology (Ogden et al. 2008), and behavior (Tomassone et al. 2018). However, our analysis found that land use change and international trade and commerce are more frequently cited as important to VBZD emergence than climate and weather. Ecological impacts due to anthropogenic landscape changes, local contact opportunities, and humans and vectors moving between or to new climatically suitable regions due to globalization outrank climate change alone as drivers of emerging VBZDs.
Our review indicates that arboviral mutations are not the exclusive drivers of outbreaks or emergence events. Several other factors have been identified as shown in Fig. 5, but for arboviruses specifically include the following: (1) viral introductions into immunologically naive populations where most hosts are susceptible to infection, (2) intense urban encroachment where humans serve as amplifying hosts in an exclusive mosquito-human-mosquito cycle, (3) expansion into new areas by travelers with subclinical disease, but are transmissible to mosquitoes, (4) movement of mosquito vectors in human vehicles from local to regional scales (Adams and Kapan 2009, Stoddard et al. 2009), (5) serologic cross reactivity of sympatric viruses that results in antibody-driven enhancement of disease severity (Dejnirattisai et al. 2016, Paul et al. 2016), or (6) genetic differences in susceptibility across human populations. Lapses in mosquito control or incomplete insecticide coverage also contributes to arbovirus re-emergence (Maciel-de-Freitas and Valle 2014).
Ecological and evolutionary attributes of VBZD
The increasing emergence and prominence of VBZDs relative to other emerging infectious diseases suggest vector-pathogen relationships are especially sensitive to global change and other drivers of disease emergence. While it is generally understood that VBZD are emerging (Jones et al. 2008, Keesing et al. 2010, Woolhouse and Gowtage-Sequeria 2005), the ecological and evolutionary reasons for this phenomenon have not been thoroughly considered. We evaluate the unique ecological and evolutionary profiles of VBZD to better understand why they are disproportionately represented among all emerging infectious diseases.
Clearly, a common feature among arthropod vectors of pathogens is blood feeding or hematophagy. Hematophagy in arthropods has arisen independently 21 times (Marquardt and Kondratieff 2005) and represents a successful evolutionary strategy, presumably because blood is highly nutritious and accessible wherever vertebrates are present. Although blood is a rich source of protein as well as fluid in arid environments, it lacks some key nutrients, particularly B-vitamins such as biotin and folic acid (Buchner 1953, Noda et al. 1997, Russell and Moran 2006, Feldhaar 2011, Nikoh et al. 2014, Hunter et al. 2015). Thus, many hematophagous arthropods such as ticks and mosquitoes rely on obligate bacterial endosymbionts to synthesize important nutrients; for instance, Wolbachia in mosquitoes or Rickettsia, Francisella, or Coxiella in ticks (Buchner 1953, Noda et al. 1997, Hunter et al. 2015). Notably, many of these endosymbionts are closely related to pathogens.
Rickettsia species are common endosymbionts of arthropods, especially ticks, and some have evolved into vertebrate pathogens (Perlman et al. 2006). Many Rickettsia species once thought to be nonpathogenic, such as R. helvetica, R. slovaca, and R. parkeri (La Scola and Raoult 1997, Shpynov et al. 2003, Paddock et al. 2004), are now known to be pathogenic. Similarly, the bacterium B. miyamotoi was presumed to be a tick symbiont for many years until it was recognized as an emerging tick-borne pathogen throughout the northern hemisphere (Fukunaga et al. 1995, Barbour et al. 2009, Padgett et al. 2014, Krause et al. 2015). Likewise, Coxiella burnetii, the etiological agent that causes Q fever, has evolutionary origins in an endosymbiotic Coxiella group (Duron et al. 2015). The close phylogenetic relationships between tick symbionts and pathogens suggest that the importance of ticks as vectors of emerging infectious diseases may stem partially from their dependence on symbiotic microorganisms, which furnish potential evolutionary sources of pathogens (Noda et al. 1997, Ahantarig et al. 2013, Duron et al. 2015). Furthermore, vector-borne viral pathogens in the families of Flaviviridae and Bunyaviridae share ancestry with mosquito-restricted viruses that are presumably nonpathogenic (Cook et al. 2006, Marklewitz et al. 2011, Bennett 2014, Chandler et al. 2014). Close symbiotic relationships between vectors and microorganisms may thus comprise an important source for emerging pathogens.
Homologous origins of vector endosymbionts and pathogens may also promote interactions within the vector between these groups (Ahantarig et al. 2013). Indeed, the vector endosymbiont community can affect the transmission potential of a variety of pathogens such as dengue (Bian et al. 2010) and Zika viruses (Dutra et al. 2016), Plasmodium parasites (Dong et al. 2009), A. marginale (Gall et al. 2016), and Borrelia burgdorferi (Narasimhan et al. 2014) through a variety of mechanisms, including induced vector immunological responses (Dong et al. 2009, Wang et al. 2009), morphological changes (Narasimhan et al. 2014), or direct microbial competition (Cook and McGraw 2010). Efforts to manipulate vector symbionts as a means of vector and disease control, such as those currently underway to curb Aedes abundances and vector competence (O'Neill 2018), are an active field of study and hold promise as a way to alter future vector-borne disease emergence rates and incidence.
Although many tick endosymbionts are vertically transmitted from female to offspring at high levels (Ponnusamy et al. 2014, Williams-Newkirk et al. 2014), some Rickettsia species have evolved a horizontal transmission pathway through a vertebrate host that allows a pathogenic relationship to evolve. Gene exchange among microbes within the vector may be important for generating new pathogens, and evidence suggests that gene transfer may contribute to some level of genetic diversity required for successful infection of vertebrates (Ogden et al. 2015). Of the recent mosquito-restricted viral pathogens within the Flaviviridae and the Bunyaviridae families, (Chandler et al. 2014, 2015), in at least one case, pathogen and presumed symbiont differ by a complete gene (Chandler et al. 2014), demonstrated experimentally to be necessary for vertebrate infection (Yamao et al. 2009).
RNA viruses as emerging pathogens
Evolutionary relationships and strategies for maintaining alternating arthropod-vertebrate-arthropod transmission of arboviruses (arthropod-borne viruses) are surprisingly conserved. Nearly all arboviruses belong to one of four RNA viral families (Togaviridae, Flaviviridae, Bunyaviridae, and Rhabdoviridae) and share common traits as pathogens. In particular, all arboviruses, except for the DNA virus African swine fever virus, possess RNA genomes that have high mutation rates due to the absence of proofreading enzymes for error correcting during replication (Hanley and Weaver 2008). In addition, Bunyaviruses have segmented genomes that often demonstrate incongruent evolutionary histories suggestive of reassortment and larger evolutionary change (Briese et al. 2013, Bennett et al. 2014). Our analyses found that Flaviviruses and Bunyaviruses were some of the most frequently reported emerging VBZDs likely due to the conservation of these traits.
Although factors promoting RNA arbovirus emergence are difficult to define, some mechanisms promoting emergence can be identified from patterns across outbreaks. The more recent emergence of chikungunya virus (Togaviridae), which initially began in Kenya in 2005, spread to Reunion Island off the coast of Madagascar, and then spread throughout Southeast Asia, was promoted by the following: (1) international travel where infected hosts (probably humans since they are the more frequent airline passengers) spread the virus long distances, (2) the unique entomological environment in the 2005 Reunion Island outbreak where the primary urban vector, Aedes aegypti, was absent, forcing a host-switch to a secondary vector, Ae. albopictus, and (3) viral mutations that augmented transmissibility for Ae. albopictus in Oceania and Southeast Asian outbreaks (Tsetsarkin et al. 2007, 2014, 2016). Arbovirus adaptation to vertebrate hosts can also promote emergence. Venezuelan equine encephalitis virus (Togaviridae) developed mutations that generated higher viremias (virus levels in blood) in horses, leading to equine epizootics and then human epidemics in Venezuela (Anishchenko et al. 2006). Thus, the ability to switch vectors and hosts under new contexts makes Flaviviruses and Bunyaviruses some of the most adaptable and nimble arboviruses.
Viral mutations are also an important emergence event, but are difficult to characterize. Dengue virus (Flaviviridae) infects 400 million people globally each year and Zika virus has spread rapidly since 2015 into more than 60 new countries and territories worldwide. In both of these pathogens, genetic mutations are linked to higher transmissibility and pathogenicity. A few mutations in dengue virus originating in Asia have been verified in the laboratory to lead to higher viremia in vertebrates and greater transmissibility in vectors and vertebrates (Rico-Hesse 2009). For Zika virus, a single mutation in the viral polyprotein has been linked to increased microcephaly and mortality in fetal mice (Yuan et al. 2017) and another mutation of a viral protein has been shown to increase infection and transmission by Ae. aegypti vectors (Liu et al. 2017). When West Nile virus (Flaviviridae) emerged in the United States beginning in 1999, a mutation was identified that increases viremia and fatality in avian hosts, causing massive mortality in American crows and promoting spread of the virus across the United States in the 2000s (Brault et al. 2007). A lineage distinguished by three mutations also emerged in 2001, which was more effective at being transmitted by United States mosquito Culex pipiens, particularly under warmer conditions (Kilpatrick et al. 2008). While these types of genetic mutations are difficult to detect and link definitively to disease emergence, more surveillance and laboratory studies can increase our ability to determine the role of genetic mutations in arbovirus emergence.
Conclusion and synthesis
The emergence of VBZDs is an illustration of how humans are inextricably connected to global ecology. The unchecked growth of the human population over the last two centuries has created a large and ever-growing ecological niche for vectors that are exceptionally good at exploiting such opportunities and the evolutionary nimble microorganisms they carry. Increased ecological opportunities brought on by human population growth and landscape transformation coupled with the enormous evolutionary capacity of these microbes, facilitated by a vector delivery system, has set the stage for the emergence of vector-borne infectious diseases.
Pathogen transmission entails complex ecological interactions and often flexible or diverse genetic or gene expression profiles. Ecological complexity combined with inherent genetic variability, especially in RNA viruses, affords opportunities for spillover and adaptation to novel vectors and vertebrate hosts and sets vector-borne pathogens apart from directly transmitted pathogens to provide one possible explanation for the former's prominence among emerging infectious diseases.
Synthesis of the emergent ecological and evolutionary properties of vectors and their pathogens is critical to develop a more complete and nuanced understanding of vector-borne disease emergence, especially in the current age of global change. We highlight several major, nonmutually exclusive drivers of VBZD emergence (e.g., land use change, international travel and commerce, and genetic mutations; Fig. 5). While many of the patterns we document here are not necessarily surprising, they do point to important paths in research and gaps in surveillance that stress the need for increasing capacity for international and interdisciplinary approaches. These types of efforts will be important in the quest to anticipate where and how VBZD have and will continue to emerge so that they can be managed.
Footnotes
Acknowledgments
This work was the culmination of an American Association for the Advancement of Sciences (AAAS) Pacific Division conference session titled, ‘Ecological and evolutionary factors driving the emergence of vector-borne diseases.’
Author Contributions
A.S. conceived of the study and conducted data collection. All authors were involved in discussion and design of analyses. A.S., L.I.C., and D.K. conducted statistical analyses. The article was drafted and revised by all authors.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This work was supported by a San Francisco State University Presidential Award to A.S. and funding was provided by the National Science Foundation to A.S.: #1750037 and 1745411.
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.