
Abstract
Trypanosoma cruzi is a parasitic protozoan that infects a diversity of hosts constituting the cycle of enzootic transmission in wild environments and causing disease in humans (Chagas disease) and domestic animals. Wild mammals constitute natural reservoirs of this parasite, which is transmitted by hematophagous kissing bugs of the family Reduviidae. T. cruzi is genetically subdivided into six discrete typing units (DTUs), T. cruzi (Tc)I to TcVI. In Brazil, especially in the state of Paraná, TcI and TcII are widely distributed. However, TcII is less frequently found in wild reservoirs and triatomine, and more frequently found in patients. The goal of this study was to investigate the natural occurrence of T. cruzi in wild synanthropic mammals captured in urban forest fragments of the Atlantic Forest of Paraná, southern Brazil. In this way, 12 opossums and 35 bats belonging to five species were captured in urban forest parks of the city of Maringá, Paraná, an area considered endemic for Chagas disease. PCR-kinetoplast DNA molecular diagnostic reveals Trypanosoma sp. infection in 12 (100%) Didelphis albiventris and 10 (40%) Artibeus lituratus. In addition to demonstrating the presence of Trypanosoma in the two groups of mammals studied, we obtained an isolate of the parasite genotyped as TcII by amplification of the cytochrome oxidase II gene by PCR, followed by restriction fragment length polymorphism with AluI, and confirmed by PCR of rDNA 24Sα. This is the first record of the encounter in wild mammals of Trypanosoma DNA (in A. lituratus) and T. cruzi DTU TcII (in D. albiventris) in the state of Paraná.
Introduction
Trypanosoma cruzi (Kinetoplastida, Trypanosomatidae), the etiologic agent of Chagas disease or American trypanosomiasis, is a parasitic protozoan capable of infecting a diversity of hosts, constituting a wild enzootic transmission cycle through vectors in humans and domestic animals. The species is subdivided into discrete typing units (DTUs), from T. cruzi (Tc)I to TcVI, as well as Tcbat (Marcili et al. 2009, Zingales et al. 2009, 2012, Lima et al. 2015). Another recently proposed classification system separates T. cruzi into just three groups, from mtTcI to mtTcIII, based on mitochondrial genes (Barnabé et al. 2016). All DTUs are found in a diverse array of mammalian hosts, representing seven different orders, and in triatomine insects (Reduviidae, Triatominae) throughout all states and biomes in Brazil (Jansen et al. 2015). However, in the state of Paraná, which is 97.8% Atlantic Forest biome, TcII is the most frequently isolated (95%) DTU in human patients with chronic infection (Gasparim et al. 2018), as well as in the triatomine species Panstrongylus megistus and Triatoma sordida, but not in wild mammals (Abolis et al. 2011). In this state, TcI has been isolated, in these same species of triatomines, both in pure and mixed infections with TcII, as well as the marsupial Didelphis albiventris.
Mammals of the orders Chiroptera and Didelphimorphia are well-known reservoirs of T. cruzi and other trypanosomatids. With ample population abundance, accentuated synanthropism, and high mobility, these animals play an important role in the distribution of diverse species of pathogens (De Oliveira et al. 2010, Hamilton et al. 2012, Brook and Dobson 2015, da Costa et al. 2015). These mammals' species demonstrate the highly complex ecological interactions found in the T. cruzi transmission cycle, being necessary the conduction of studies that evaluate its dynamics in the context of the One Health concept, triad that encompasses humans, domestic animals, wildlife, and the changing ecosystems in which they live (Thompson 2013, Pinazo and Gascon 2015).
In this study, a preliminary investigation into the presence of Trypanosoma in free-range wild mammals was accomplished in Atlantic Forest remnants situated in urban areas, an environment not still explored in the state of Paraná.
Materials and Methods
Study area
Wild mammals were captured in two urban forests fragments, in 23°25′39′ of latitude south and 51°55′51″ of longitude west, and in 23°23′27′ of latitude south and 51°56′34″ of longitude west. Both, remnants of the Atlantic Forest biome of the city of Maringá, located in the northeast of Paraná, southern Brazil, and an area considered endemic for Chagas disease (Fig. 1).

Map highlighting the two capture points located in the urban area of the Municipality of Maringá, state of Paraná, Southern Brazil.
Wild mammal capture
The campaigns were held once a month in the following periods: from September 2014 to December 2015. Bats were captured with mist nets, conditioned in cloth sacs, and identified at the species level. Opossums of the species D. albiventris were captured using Tomahawk Live Traps baited with fruits and peanut butter. These methods were approved by the Instituto Chico Mendes de Conservação da Biodiversidade—Ministério do Meio Ambiente, Brazil (no. 42881) and by the Ethics Committee in Animal Experimentation (CEUA) at Universidade Estadual de Maringá (UEM) (no. 023/2014).
Blood collection and hemocultures
Opossums and bats were anesthetized with 4.0 mg/kg chloridrate of xylazine and 20.0 mg/kg body weight chloridrate of ketamine for blood collection. Approximately 0.5 mL of whole blood obtained by cardiac puncture in the bats and from the tail medium vein in the opossums was placed in Novy, McNeal, and Nicolle medium, and covered with an overlay of liver infusion tryptose (LIT) medium containing 10% fetal bovine serum. Another 0.2 mL was stored in microtubes containing 400 μL of 0.2 M ethylenediaminetetraacetic acid (EDTA) and 6.0 M guanidine solution for the extraction and amplification of blood DNA according to Miyamoto et al. (2006).
Molecular analysis for detection of Trypanosoma DNA
DNA was extracted by the phenol/chloroform method and precipitated with addition of ethanol and sodium acetate (Macedo et al. 1992). PCR amplification was performed with oligonucleotides number 121 (5′-AAATAATGTACGGG(T/G)GAGATGCATGA-3′) and number 122 (5′-GGTT CGATTGGGGTTGGTGTAATATA-3′) according to the protocol of Gomes et al. (1998) modified by Miyamoto et al. (2006). These sequences amplify a specific fragment of ∼330 base pairs (bp) of kinetoplast DNA (kDNA) from Trypanosoma sp. The amplification products were analyzed using 4.5% polyacrylamide gel electrophoresis, stained with silver, and digitally recorded.
Isolation and genotyping of T. cruzi
Parasites from a unique positive hemoculture, with low manipulation to avoid clonal selection, were amplified in LIT medium until the exponential phase. This material was centrifuged and washed with Krebs-Ringer-Tris/pH 7.2 solution buffer three times. The obtained mass was then resuspended in 500 μL of lysis solution (0.5 M EDTA, 5 M sodium chloride and 1% sodium dodecyl sulfate) supplemented with 10 mg/mL of proteinase K (Invitrogen) at 37°C overnight. DNA extraction was performed by the phenol/chloroform method after Sá et al. (2016).
PCR/restriction fragment length polymorphism cytochrome oxidase II
Differentiation of T. cruzi isolates (TcI to TcVI) was done by amplification of the cytochrome oxidase II (COII) gene by PCR, followed by restriction fragment length polymorphism (RFLP) with AluI (de Freitas et al. 2006, Abolis et al. 2011). The primers used were the Tcmit-10 (5′-CCATATATTGTTGCATTATT-3′) and Tcmit-21 (5′-TTGTAATAGGAGTCATGTTT-3′) primers with Taq DNA polymerase (Platinum, Invitrogen) and the restriction enzyme AluI (New England BioLabs) in NEB 4 buffer. The PCR amplicons were resolved on 6% polyacrylamide gel and products sizes analyzed as described by Sá et al. (2016).
PCR of rDNA 24Sα
PCR amplification of rDNA 24Sα using D71 (5′-AAGGTGCGTCGACAGTGTGG-3′) and D72 (5′-TTTTCAGAATGGCCGAACAGT-3′) primers as previously described by Souto et al. (1996). Amplified fragments were resolved on 6% polyacrylamide gel, stained with silver, and analyzed according to Sá et al. (2016).
Results
Detection of the kDNA minicircle in Trypanosoma
PCR-kDNA revealed that all samples of captured D. albiventris (n = 12, 100%) exhibited a fragment of 330 bp kDNA minicircle from Trypanosoma. Of the 35 examined bat species, PCR-kDNA also detected this same fragment in 10/25 (40%) Artibeus lituratus caught. In other species of captured bats, Sturnira lilium (n = 4), Carollia perspicillata (n = 3), Pygoderma bilabiatum (n = 2), and Platyrrhinus lineatus (n = 1), this fragment was not detected (Table 1). This high rate of positivity in PCR was not observed in hemocultures, and in only 1/12 (8.3%) opossums was positive in this technique, allowing to obtain a T. cruzi isolate (Table 1).
Wild Mammals Captured and Examined for the Presence of Trypanosoma cruzi (By Detection of Kinetoplast DNA PCR and Hemoculture) in Two Atlantic Forest Urban Parks of Maringá City, Paraná, Southern Brazil
Genotyping of DTU of T. cruzi isolate
The isolate was genotyped by analysis of mitochondrial COII gene polymorphisms. The PCR/RFLP-COII showed two bands, the first at ∼80 bp referring to the species T. cruzi and the second at 250 bp, which consistently classifies the isolate as TcII (Fig. 2A).
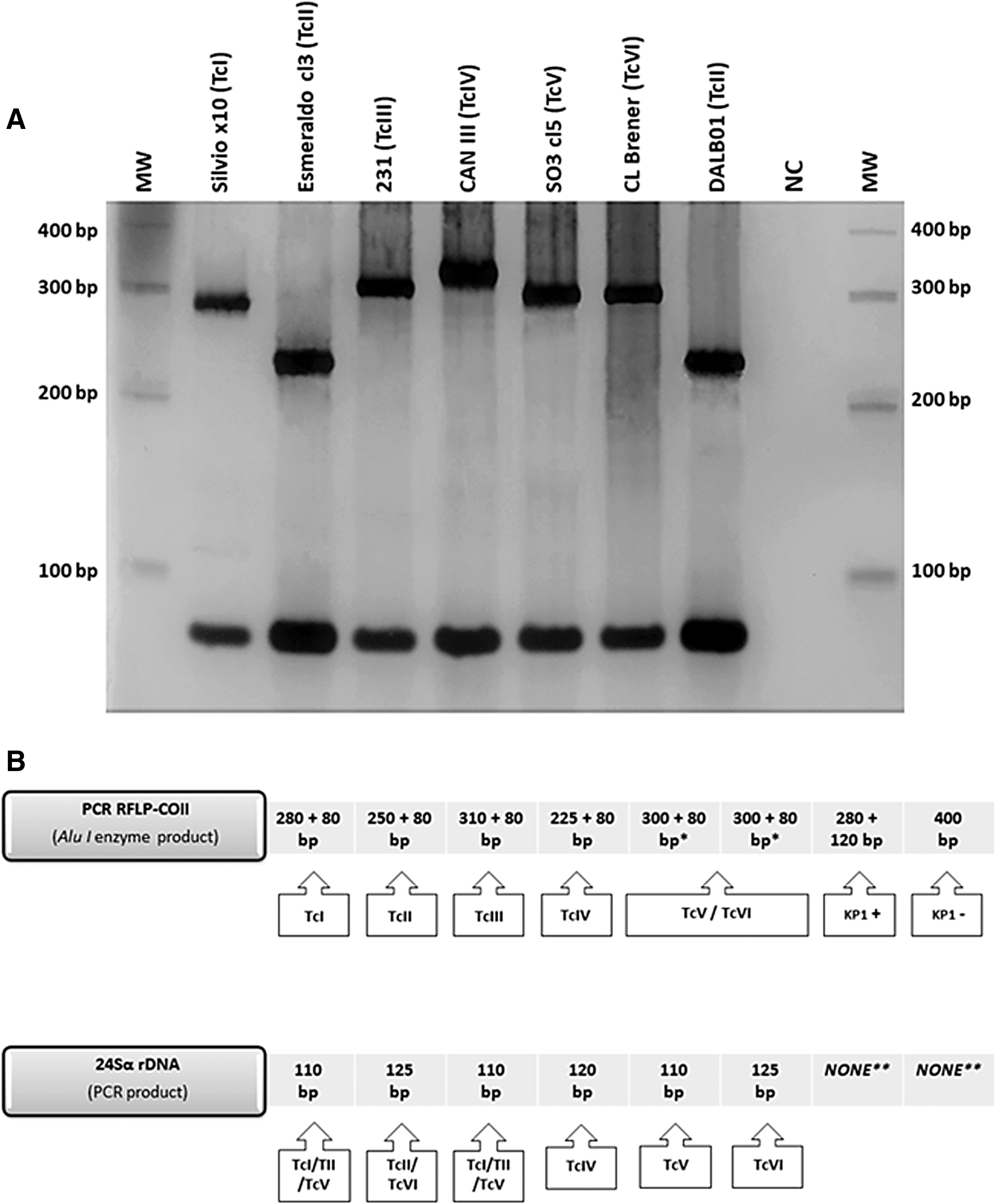
Amplification and restriction fragment profiles of PCR/RFLP-COII in silver-stained 6% polyacrylamide gel used for genotyping Trypanosoma cruzi isolate (MDID/BR/2015/DALB01) obtained from Didelphis albiventris
The result of PCR amplification of the rDNA 24Sα showed a band of ∼125 bp compatible with the TcII and TcVI genotypes. Although this marker is not able to distinguish these two genotypes, it was used to confirm the DTU of the isolate (Zingales et al. 2009, 2012). The profiles generated by the genetic analyzes of COII and rDNA 24Sα together, allowed to classify the T. cruzi isolate obtained as TcII (Fig. 2B).
Discussion
Studies of the dynamic population of T. cruzi in Paraná, located in the Southern Region of Brazil, an endemic area for Chagas disease (Gasparim et al. 2018), reveal TcII to be the principal DTU isolated from chronic phase patients residents in this state (Zalloum et al. 2005, Abolis et al. 2011). In contrast, a wild host of this DTU has yet to be registered in the state. TcII was surprisingly found in this central urban forest (Atlantic Forest biome) of the city of Maringá in the northwest of the state, in D. albiventris, a wild reservoir generally considered uncommon for harboring this DTU.
All previous attempts to associate T. cruzi populations with specific mammalian host species have led to controversial results (Jansen et al. 2015). Even without an association between DTUs and mammalian reservoirs, a majority of isolates from D. albiventris are genotyped as TcI, an encounter with TcII being a rarity. When encountered, TcII is generally associated with mixed infections of TcI, TcV, or TcVI (Abolis et al. 2011, Jansen et al. 2015, 2018). However, the ability of Didelphis spp. to rapidly control TcII infections and reduce parasitemia to undetectable levels (Legey et al. 2003, Jansen et al. 2015) is a likely variable affecting isolation efficiency of TcII from these animals.
D. albiventris is characterized as a highly synanthropic species, adaptable, opportunistic, and omnivorous, as well as potential disseminators of disease and one of the main reservoirs of T. cruzi and potentially other Trypanosoma spp. (De Oliveira et al. 2010, Jansen et al. 2015, Dario et al. 2017), which justifies our single encounter of TcII. Jansen et al. (2015) reported the finding of TcII infecting wild animals in all Brazilian biomes, including the Caatinga in the Northeast Region of Brazil, where that species of marsupial has been found harboring TcII, in both pure and mixed infections with TcV and TcVI. However, its discovery was never recorded in Paraná, a state that has 97.8% of its area inserted in the Atlantic Forest biome (Muylaert et al. 2018). This is the first registration of T. cruzi genotyped as TcII isolated from D. albiventris in the state of Paraná.
The high positivity of PCR-kDNA for Trypanosoma spp. in D. albiventris and the nonencounter of triatomines in the area of study could be related to the complexity of parasitic cycles in the wild environment. Nouvellet et al. (2013) states that the probability of transmission after contact with an infected triatomine is 6.0 × 104, confirming that transmission is still more difficult in free-range wild mammals were thick, dense skin impedes contact with the metacyclic forms of the parasite (Jansen et al. 2015). The same holds true for infected A. lituratus specimens, in addition to the large diversity of Trypanosoma spp. with which they are subject to becoming infected (Dario et al. 2017, Jansen et al. 2018, Lourenço et al. 2018).
Although no triatomine was found in the study area, the TcII encounter with the high positivity of D. albiventris for Trypanosoma sp. suggest an active wild cycle. Opossums and bats are reservoirs of T. cruzi that are frequently found in the peridomicile (Abolis et al. 2011, Jansen et al. 2015). Therefore, surveillance work is required that encompasses synanthropic animals in urban and periurban forest areas to avoid reintroduction or a spillover of T. cruzi to the domestic cycle in controlled areas for Chagas disease (Pinazo and Gascon 2015, Gasparim et al. 2018). The encounter of TcII in D. albiventris, the main isolate obtained in chronic patients in Paraná, suggests that surveillance actions in these areas with a focus on synanthropic wild animals should be considered.
PCR-kDNA analysis of T. cruzi is used in many experimental studies, both for proving the infection and for monitoring the cure of treated animals due to its high sensitivity and specificity to T. cruzi (Teston et al. 2013, Margioto et al. 2016, Zanusso Junior et al. 2018). This is why it has also been used in the laboratory diagnosis of American trypanosomiasis in chronic human patients (Ramírez et al. 2015). Despite the high sensitivity, its use is limited to the genus Trypanosoma, when applied to the parasite's detection in wild animals. This is because wild animals are exposed to a wide range of Trypanosoma spp., some of which have not yet been cataloged or even recently discovered (Lopes et al. 2018), requiring the use of several molecular markers to identify these species unequivocally. However, the use of more than one molecular marker for blood samples obtained from wild animals becomes infeasible, since the variety of Trypanosoma spp. that can infect didelphids and chiroptera is immense (Dos Santos et al. 2018, Jansen et al. 2018, Lourenço et al. 2018).
In bats, the presence of Trypanosoma infection detected through PCR-kDNA accompanied by the difficulty in obtaining parasite isolates could be directly related to these mammals' capacity to incubate pathogens (Brook and Dobson 2015), or still to the presence of Trypanosoma spp. infection not grown in axenic medium (Dario et al. 2017, Jansen et al. 2018). In the same way, D. albiventris has already been found to be opportunistically predating on A. lituratus (Gazarini et al. 2008), which may be the tip of the iceberg for a semiopen cycle or spillover between niches in urban and periurban areas. Whereas bats weakened by concomitant infections, together with poor dietary supply, may be the source of infection for this didelphid. The encounter of A. lituratus infected by Trypanosoma sp., a species that presents high population densities and high synanthropism, reinforces the importance of studies that focus on synanthropic species in the wild cycle of T. cruzi. This register contrasts with the finding of a migratory bat (Tadarida brasiliensis) found infected with TcV in the southern United States (Nichols et al. 2019). The authors report that even with a low prevalence, T. brasiliensis may contribute to future enzootic expansion of T. cruzi, playing a unique role in the epidemiology of T. cruzi through its annual migrations (Nichols et al. 2019).
PCR-RFLP that targets the COII gene restricted by the enzyme AluI is able to accurately separate the T. cruzi genotypes into five groups: TcI, TcII, TcIII, TcIV, and TcV/TcVI (de Sá et al. 2013, Sá et al. 2016). In its turn, the TcV/TcVI pattern can be differentiated using a second genetic marker based on the 24Sα gene sequence of Trypanosoma ribosomal DNA (de Sá et al. 2013). These markers together still allow identification of T. rangeli (a very common species and frequently isolated from the wild environment), and the separation of its major genotypes: KP1+ and KP1− (Sá et al. 2016). This pair of techniques, therefore, allows to classify stocks of T. cruzi within the six genetic lineages previously proposed with a high resolution, hence its use being of great value in population studies with obtaining isolates. This pair of techniques used allowed the isolate obtained from D. albiventris to be genotyped as DTU TcII (Fig. 2).
In previous studies, T. cruzi isolates obtained from humans living in Paraná, all in the chronic phase of infection, were genotyped as TcI, TcII, and TcIII, with TcII being the most prevalent DTU (Abolis et al. 2011). In contrast, TcII had previously been found in triatomines in genetically pure or mixed infections with TcI, whereas the isolates obtained from marsupial D. albiventris presented only TcI profiles (Zalloum et al. 2005, Abolis et al. 2011). These data allow the first record of TcII isolate from a wild D. albiventris from an urban forest fragment of the state of Paraná.
Conclusion
PCR-kDNA, despite detecting the presence of kDNA restricted to the Trypanosoma genus, is shown as an alternative for the evaluation of groups of animals that act as reservoirs in areas with potential of transmission. Because it is a low-cost technique and requires a small amount of blood, PCR-kDNA has proved to be of great value in the monitoring of infection, allowing more efficient isolation strategies to be used in places where active cycles of Trypanosoma spp. are detected. This strategy may potentiate the number of isolates obtained in forest areas yet to be explored as to the genetic diversity of the parasites. The results show a relation of the synanthropic mammals sampled with persistent infections by one T. cruzi DTU (TcII), or even by Trypanosoma spp., which may be circulating in the wild environment of the region, but are more difficult to detect or isolate by more conventional techniques, such as hemoculture. Infection by Trypanosoma spp. was recorded in A. lituratus that inhabits and forages urban forest areas and their surrounds in the city of Maringá. The TcII genotype was also observed in D. albiventris, an unusual finding that had not yet been recorded in a wild mammal in the state of Paraná.
Footnotes
Acknowledgments
The authors thank Prefeitura Municipal de Maringá and Secretaria de Meio Ambiente e Bem Estar Animal (SEMA) for the support.
Author Disclosure Statement
No competing financial interests exist.