
Abstract
Hantaviruses are worldwide pathogens, which often cause serious or even fatal diseases in humans. Hosts are predominantly in the form of rodents and soricomorphs; however, bats are also described as an important reservoir. In Hungary, representatives of two human pathogenic species of the genus Orthohantavirus are present: the Dobrava-Belgrade orthohantavirus and Puumala orthohantavirus. In Hungarian forests, the dominant rodent species are Apodemus flavicollis, Apodemus agrarius, Apodemus sylvaticus, and Myodes glareolus, all of which are natural reservoirs comprising different hantaviruses. The aim of the study was to survey the prevalence of hantaviruses among rodent populations and examine the potential relationship regarding population densities, years, sex, and seroprevalence. Rodents were trapped at 13 sampling plots in a forest reserve located in the Mecsek Mountain range, Hungary, from March to October between 2011 and 2014. Rodent serum samples were tested for IgG antibodies against Dobrava-Belgrade virus and Puumala virus by enzyme-linked immunosorbent assay (ELISA) using a recombinant nucleocapsid protein. During the 4-year sampling period, 2491 specimens were tested and 254 (10.2%) proved seropositive for orthohantaviruses. In 2011, the seroprevalence among Apodemus spp. and M. glareolus was 17.2% (114/661) and 3.9% (3/77), respectively, although this rate had reversed itself in 2014. Seropositivity was substantiated in 18.4% (12/65) of Myodes voles, while only 3.6% (13/359) of the tested Apodemus rodents were found to be IgG positive. Seroconversion was observed in 58 cases, while seroreversion was only detected in 3 individual cases. A significant difference among the number of infected males and females was identified in the first 2 years of our study. Winter survival with respect to rodents was not negatively affected due to the hantavirus infection. Hantavirus seroprevalence was not directly influenced by host abundance. Consequently, we assume that high rodent density alone does not lead to an increased risk of hantavirus infection among the rodent host population.
Introduction
Orthohantaviruses are members of the Hantaviridae family. All have a negative-sense, single-stranded, tripartite RNA genome. They are known to cause serious and often fatal human diseases, including hemorrhagic fever with renal syndrome (HFRS) in Eurasia and hantavirus cardiopulmonary syndrome in America. The hosts of these viruses are rodents, soricomorphs, and bats. Viruses are transmitted to humans through inhalation of dust particles covered with virus-contaminated rodent excreta or directly due to a rodent's bite (Vaheri et al. 2013).
In Europe, there are two major human pathogenic hantaviruses; Dobrava-Belgrade virus (DOBV) is carried by the yellow-necked mouse (Apodemus flavicollis), striped field mouse (Apodemus agrarius), and wood mouse (Apodemus sylvaticus), while the bank vole (Myodes glareolus) is the natural host reservoir of Puumala virus (PUUV). Among these rodents, A. flavicollis is the dominant species in Hungarian deciduous forests, including a number of similar European habitats (Schmaljohn and Dalrymple 1985, Plyusnin et al. 1996, Jonsson et al. 2010).
Nearly 150,000–200,000 human cases of hantavirus infection are reported annually worldwide (Schmaljohn et al. 1997). DOBV causes HFRS, while PUUV infection results in a milder form of HFRS, which is clinically defined as nephropathia epidemica (NE). NE is the most prevalent disease in Western and Central Europe. Nowadays, PUUV is responsible for more than 9000 cases per year in Europe with a mortality rate of 0.1–0.4%, while the most severe forms of HFRS are associated with DOBV, with a mortality rate ranging from 3% to 12%, respectively (Bi et al. 2008).
Numerous studies revealed that PUUV outbreaks in humans are associated with high densities of bank voles. PUUV epidemics throughout Western Europe showed multiannual cycles with peaks every 2–3 years, following vole population abundance cycles (Heyman et al. 2001, 2007, Sauvage et al. 2003, Kallio et al. 2009, Tersago et al. 2011). Rodent population fluctuations are thought to be driven by predators, climate, weather, and food availability (Cavanagh et al. 2004, Monchatre-Leroy et al. 2017). Human DOBV infections exhibit similar characteristics throughout the Balkans (Hukić et al. 2003, Rogozi et al. 2013).
There are five hantavirus species circulating in Hungary: DOBV, PUUV, Tula orthohantavirus, Seewis orthohantavirus, and Nova mobatvirus (Scharninghausen et al. 1999, Jakab et al. 2007a, 2008, Kang et al. 2009a, 2009b, Németh et al. 2013). The clinical importance of DOBV and PUUV among both hospitalized patients and risk populations was well established in several studies in Hungary (Jakab et al. 2007b, 2011, Oldal et al. 2014a, 2014b).
In this multiannual study, the objective was to determine the dynamics regarding PUUV and DOBV infections among rodents in a temperate mixed deciduous forest in Hungary. The specific aims of the present study were (1) to assess the abundance of captured rodents, (2) to determine the seroprevalence of DOBV and PUUV infections, (3) to examine the number of infected rodents and seroprevalence in relation to population abundance of Apodemus and Myodes, (4) to characterize the seasonal and annual changes of PUUV and DOBV throughout the 4-year study period, and finally (5) to evaluate the seasonal and annual differences of hantavirus seroconversion.
Materials and Methods
Sample collection
The study area was the Kőszegi-forrás Forest Reserve in Mecsek Mountain range (46°09′28.88″ N, 18°17′09.90″ E) in southern Hungary where DOBV was previously detected in Apodemus spp. (Németh et al. 2011). The forest reserve lies on both sides of a deep valley and is in a joining plateau, with an elevation of 320–400 meters above sea level. There were a total of 468 rodent live traps located in 13 sampling quadrats on a roughly 11-ha area. Rodents were trapped from 2011 through 2014 during an 8-month period (from March to October) each year. The capture–mark–recapture (CMR) method was applied using plastic box-type live traps placed in 6 × 6 grids with 5-m spacing between traps. Bacon and various forms of cereal anointed with aniseed extract and vegetable oil were mixed together and used as bait. Every month, a standard five-night sampling period was carried out, in the course of which traps were examined every morning.
Captured animals were marked by toe tattooing, followed by recording data on each individual specimen regarding sex, age, and body mass. The scrotal position of the testicles in males and gravidity, lactation, and perforated vagina for females were considered as signs of sexual activity. Body weight (±0.5 g) was measured using a 100-g Pesola spring balance. Age was determined based on external features and body weight and categorized as juvenile: immature pelage, or adult: fully matured adult molt (Gurnell and Flowerdew 2006).
Blood samples were extracted from all trapped animals each month using retro-orbital sinus bleeding, which was repeated upon recapture. With regard to individual animals, blood was taken only once per month. The hantavirus infection of marked individuals was monitored based on the antibody presence in these monthly blood samples. Based on the CMR method, the abundance of rodent (Apodemus and Myodes) populations was determined using the minimum number alive (MNA) as a population index suitable for assessment regarding changes in the abundance of rodents (Krebs 1966, Boonstra et al. 1978, Slade and Blair 2000, Brehme et al. 2011). Based on the analysis of monthly samples, the minimum number of infections (MNI) was calculated as an index of the number of individuals in which antibodies against hantavirus were present (Luis et al. 2010, Carver et al. 2011).
Serological screening by enzyme-linked immunosorbent assay
Rodent blood samples were screened for the presence of IgG antibodies against DOBV and PUUV by standard indirect enzyme-linked immunosorbent assay (ELISA). For ELISA analysis, we applied recombinant DOBV and PUUV antigens, both produced in an Escherichia coli bacterial expression system, as described previously (Németh et al. 2011). Microtiter plates (MaxiSorp Nunc-Immuno Plate, Nunc) were coated using a combination of DOBV and PUUV antigens, followed by incubation for 2 h at 37°C. Excess antigens were removed using a rinse cycle (1 × 30 s) with phosphate-buffered saline (PBS) (pH = 7.5), containing 0.05% Tween-20 (Sigma) (PBS-T). Plates were blocked for 2 h at 37°C, using PBS containing 5% nonfat dry milk (Blotting-Grade Blocker; BIO-RAD) and 5% sucrose (Merck).
Rodent serum samples were used in 1:100 dilutions in PBS containing 2.5% nonfat dry milk. Following incubation for 1 h at 37°C, wells were rinsed with PBS-T (5 × 5 min). Horseradish peroxidase-conjugated rabbit anti-mouse IgG (Dako) was used as the secondary antibody, diluted 1:1500 in PBS containing 2.5% nonfat dry milk. Plates were incubated at 37°C for 1 h, followed by a second rinse cycle with PBS-T (5 × 5 min). Color development was implemented using 100 μL of 3,3′,5,5′-tetramethylbenzidine substrate (BD OptEIA), and incubation was carried out at room temperature in absolute darkness for 15 min. The reaction was ceased by addition of 100 μL of 2 M sulfuric acid, and optical density was measured using a microplate spectrophotometer (Thermo Electron Corporation) at 450 nm. The cutoff value was calculated as previously described (Dobly et al. 2012a).
Statistical analyses
Pearson's chi-square tests were performed to evaluate the differences in the total number of tested and infected rodents (Apodemus and Myodes), the seroprevalence calculated from cumulative data, and the total number of males and females. Furthermore, seasonal distribution of these parameters is summarized in Table 2, and seasonal differences regarding cumulative values were assessed using Pearson's chi-square test. Seasonal characteristics were determined as follows: spring (March–May), summer (June–August), and autumn (September and October).
Summary of Captured Rodent Species by Years and Seasons and Corresponding Hantavirus Seroprevalence Rates
The total amount of examined rodents is not equal to the sum of the annual specimens (Table 1) as recaptures were taken into consideration (the same animal could be captured more times in different seasons).
Based on monthly data, boxplot (medians ± percentiles (25–75%) and nonoutlier range) was used to present the median values of tested and infected rodents and seroprevalence values in comparison of Apodemus and Myodes. Furthermore, Wald–Wolfowitz runs test (Sprent and Smeeton 2007) was performed to evaluate the difference regarding these parameters between the two tested animal groups. Based on monthly values of MNA, MNI, and hantavirus seroprevalence, which were calculated from CMR data, the linear regression method was performed to analyze the relationship between population abundance (MNA) and number of infected animals (MNI) and between MNA and seroprevalence in case of Apodemus and Myodes, respectively.
Additionally, a linear regression method was used to investigate the temporal changes of infected animals and hantavirus seroprevalence based on the monthly data of all tested animals, comprehensively, and the two genera separately. Temporal changes in seroprevalence of males and females of all species were analyzed using the same method.
The distribution regarding the average values of hantavirus seroconversion is presented in boxplots (mean ± standard error, lower and upper limits of 95% confidence interval), and the Kruskal–Wallis test (followed by Dunn's post hoc test for multiple comparisons) was used to compare the amount of seroconversion between different seasons and years, respectively (Zar 2010). All statistical analyses were conducted using STATISTICA 12 software (StatSoft, Inc. 2014). Statistical tests were considered significant at the level p ≤ 0.05, deemed as standard in all analyses (Sokal and Rohlf 2012).
Ethics statement
The South-Transdanubian Inspectorate of Environment Protection, Nature and Water Conservation (Hungary) provided an ethics statement authorizing the trapping and marking, including blood sampling, of the rodents encompassed within the area of the Kőszegi-forrás Forest Reserve.
Results
Comprehensively, 3598 blood samples were collected from 2491 rodents during a period of 140 trapping days. Rodents were categorized representing four species; 45 (1.8%) A. agrarius, 1749 (70.2%) A. flavicollis, 180 (7.2%) A. sylvaticus, and 517 (20.8%) M. glareolus were sampled. The sex ratio of each species was approximately equal since there was no significant difference (χ 2 = 3.038; df = 1; p = 0.081) between the number of male and female rodents (Table 1). Due to a large deviation among the number of different DOBV host Apodemus species, we did not analyze them separately and they were considered as one group (Apodemus spp.) in this study.
Total Number of Individuals Collected in the 4-Year Study
In the course of the 4-year-long screening, the total number of individuals of Apodemus spp. was significantly higher than the tested number of M. glareolus (χ 2 = 852.207; df = 1; p < 0.001). All serum samples were screened for hantavirus IgG using ELISA tests with mixed PUUV and DOBV antigens. Of the 2491 animals, 254 were seropositive for hantaviruses. Furthermore, 31 cases were also positive; however, these samples distinctly contained maternal antibodies. There was a significant difference between a total number of infected Apodemus spp. (217;10.99%) and Myodes (37;7.15%) (χ 2 = 127.559; df = 1; p < 0.001), while a significant difference regarding the total hantavirus seroprevalence was not detected among the rodent genera during the entire duration of the study (4 years) (χ 2 = 0.808; df = 1; p = 0.369). An intensified, robust, complex statistical analysis regarding these data confirmed previously acquired results: distribution of the total number of tested specimens (Wald–Wolfowitz runs test: z = 2.12, p < 0.05) and the number of infected animals (z = 3.18, p < 0.01) differed significantly, while there was no significant difference in distribution of seroprevalence values between Apodemus and Myodes (Wald–Wolfowitz runs test: z = 1.59, p = 0.112) (Fig. 1).
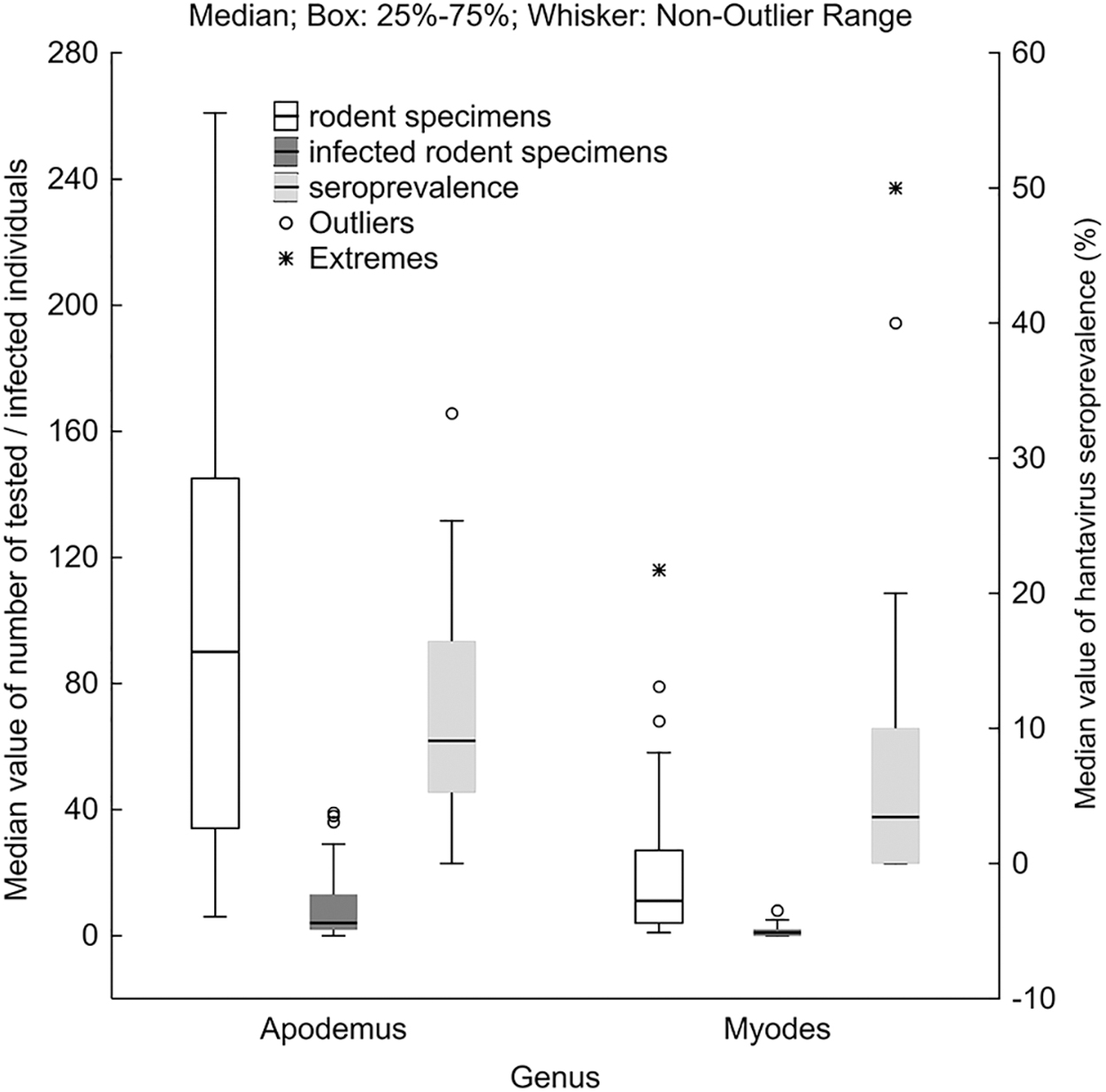
Median value of individuals (white), infected rodent specimens (dark gray), and hantavirus seroprevalence (light gray) in comparison of Apodemus and Myodes.
In 2011, the seroprevalence among Apodemus spp. was 17.25% (114/661), while it was only 3.9% (3/77) for M. glareolus. Although the total number of captured animals was significantly higher (χ 2 = 110.556; df = 1; p < 0.001) in the subsequent year (2012), the DOBV seroprevalence decreased to 10.2% (88/864). From another perspective, the number of seropositive Myodes voles increased to 5.3% (18/337) in the same year. We experienced a population decline in both genera in 2013. The seroprevalence of Apodemus exhibited a further decrease to 5.3% (χ 2 = 1.529; df = 1; p = 1.216), while in the case of Myodes, it was even higher (8%) when compared with the previous year (χ 2 = 0.530; df = 1; p = 0.466). Despite the significant increase in abundance of Apodemus in 2014 (χ 2 = 53.457; df = 1; p < 0.001), the seroprevalence steadily experienced a decline to 3.6% (13/359). Contrarily, there was no significant increase among the number of voles (χ 2 = 1.956; df = 1; p = 0.162), although seroprevalence was two times higher than previously recorded; 18.5% (12/65).
There was no significant difference regarding the positivity of Apodemus mice between the seasons within a given year (Table 2). The same seasonal change was observed in the case of Myodes, highlighting a difference in which significant deviation was detected in the spring of 2014 (χ 2 = 12; df = 1; p < 0.001). During the four monitored spring seasons, seroprevalence among Apodemus mice was nearly identical, with the exception occurring in 2014, in which it was lower when compared with previous years.
In the summer of 2011, the number of DOBV-infected rodents was significantly higher than in the same seasons of the following 3 years (χ 2 = 5.87; df = 1; p < 0.05). The same pattern was observed in the prevalence data for autumns of different years: in 2011, it was 23%, while in the period from 2012 to 2013, the seroprevalence was about 5–9% (χ 2 = 8.53; df = 1; p < 0.01). The number of Apodemus mice was considerably low during the autumn of 2014, therefore the data were not deemed suitable regarding statistical analysis. In the case of hantavirus-seropositive voles, there were noticeable differences.
The number of infected rodents among the four examined springs spiked in 2014. Regarding summer seasons, a significant growth in seroprevalence data was observed (χ 2 = 6.95; df = 1; p < 0.05), following the first year of low prevalence level, in 2012 and 2013. The number of Myodes voles was substantially lower during the summer of 2014, essentially ruling out the credibility regarding the acquired data and therefore deemed unsuitable for statistical analysis (Table 2).
Based on the monthly data of individual capture histories, our results demonstrate a significant linear regression between MNA and MNI in both genera, although the relationship was stronger in the case of Apodemus, shown by the larger regression coefficient (Fig. 2A, B). However, hantavirus seroprevalence was independent of the population size represented by the MNA values in the case of both genera (Fig. 2C, D).
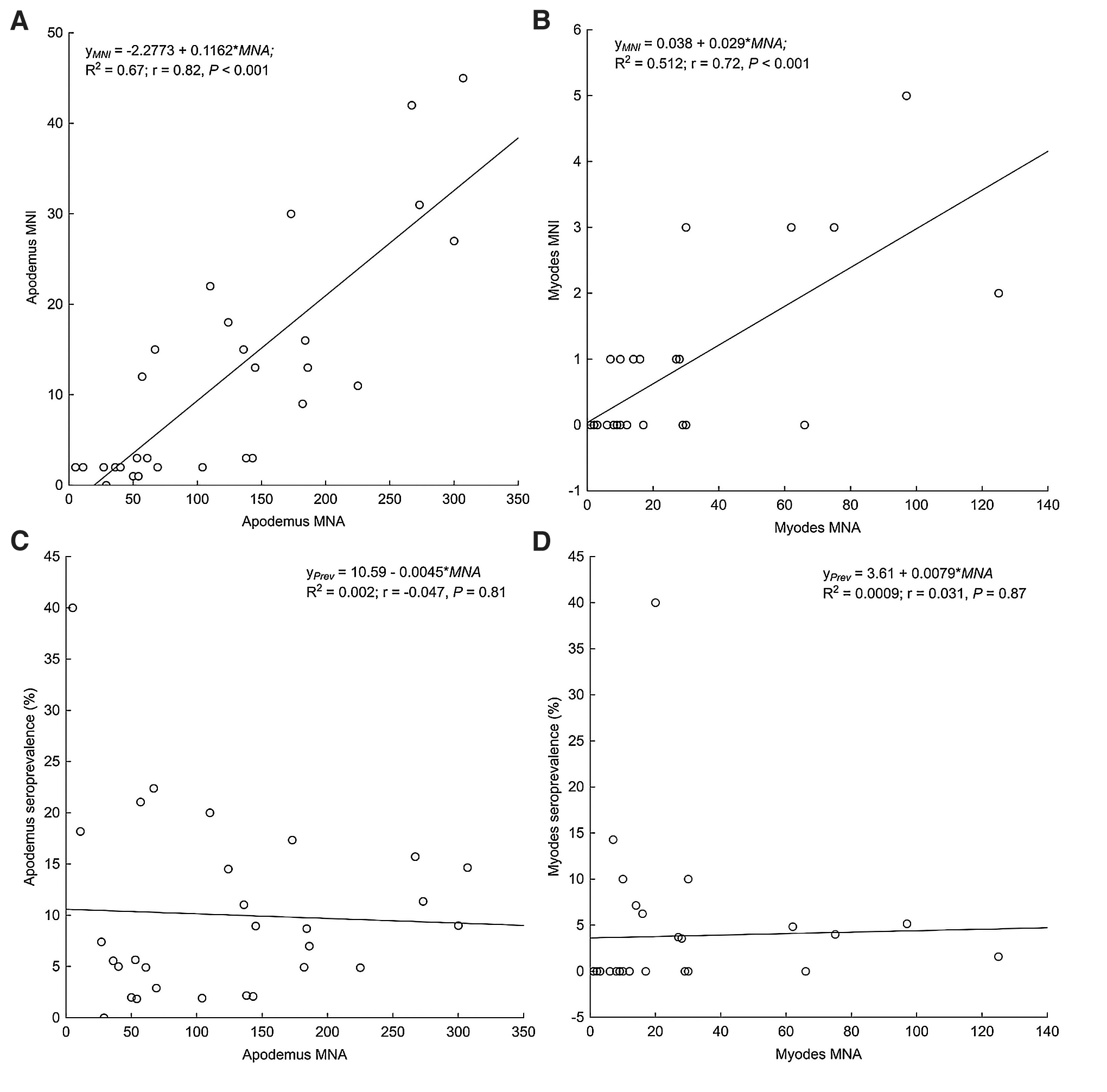
Minimum number of infected animals (MNI)
Based on linear regression, the annual changes in the number of infected rodents and hantavirus seroprevalence showed a significant decline during the 4-year-long study when all tested animals were taken into account, although the degree of the decrease was lower in the case of seroprevalence than the number of infected individuals, as shown by the difference of the slopes of linear functions (Fig. 3). When the two genera were analyzed separately, the annual changes of infected animals and hantavirus seroprevalence revealed differences in the comparison of Apodemus and Myodes. In the case of Apodemus, the number of infected individuals followed a significant linear decreasing trend, while the number of infected bank voles exhibited a slightly positive, yet not overly significant, trend during the 4 years (Fig. 4A). Hantavirus seroprevalence of the two genera also showed a contrary tendency, neither the decrease of Apodemus nor the increase of Myodes seroprevalence proved significant (Fig. 4B). In the assessment of the total number of tested animals, we found a remarkable difference in the annual change of hantavirus seroprevalence among the two sexes. While a significant decline of seroprevalence was observed for males, the regression analysis showed a negligible decrease in the case of females, reaching equal values in 2014 (Fig. 5).
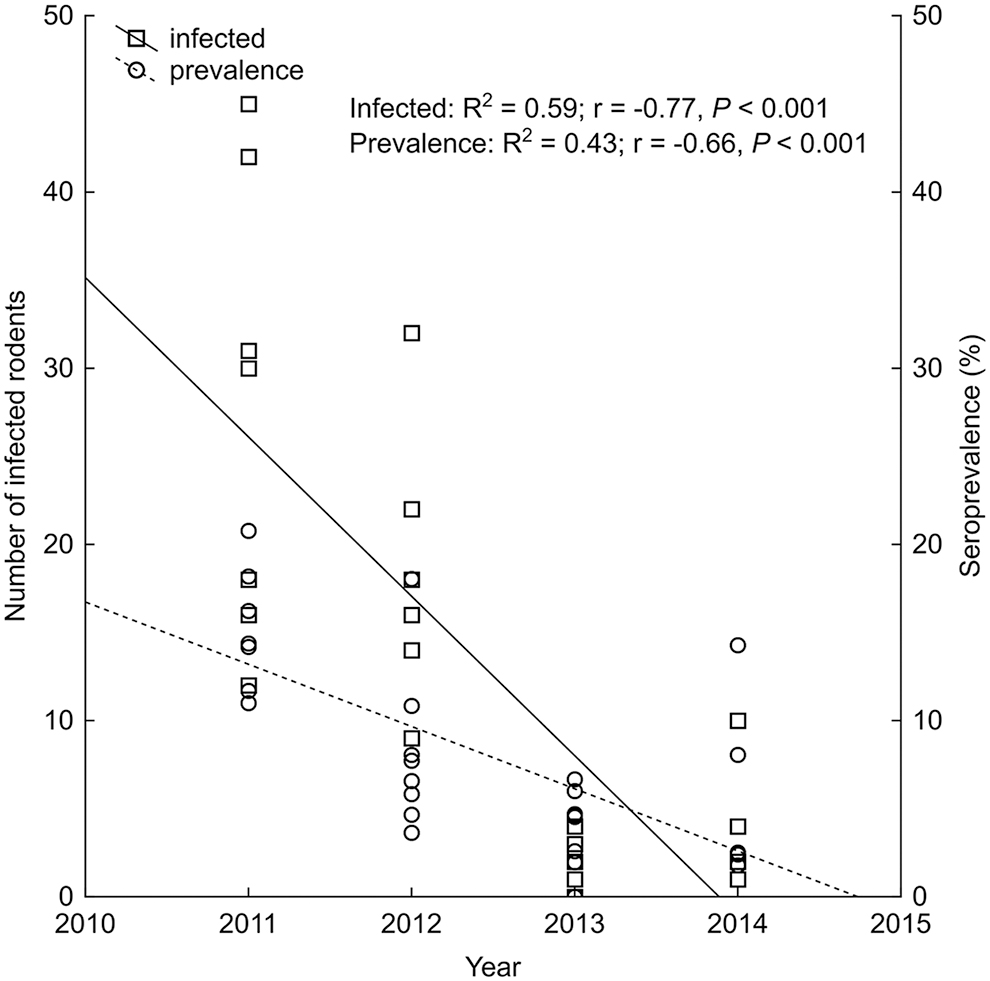
Annual changes of total hantavirus-infected rodents and seroprevalence during the 4-year study period.

Comparison of the annual changes of number of hantavirus-infected Apodemus and Myodes rodents
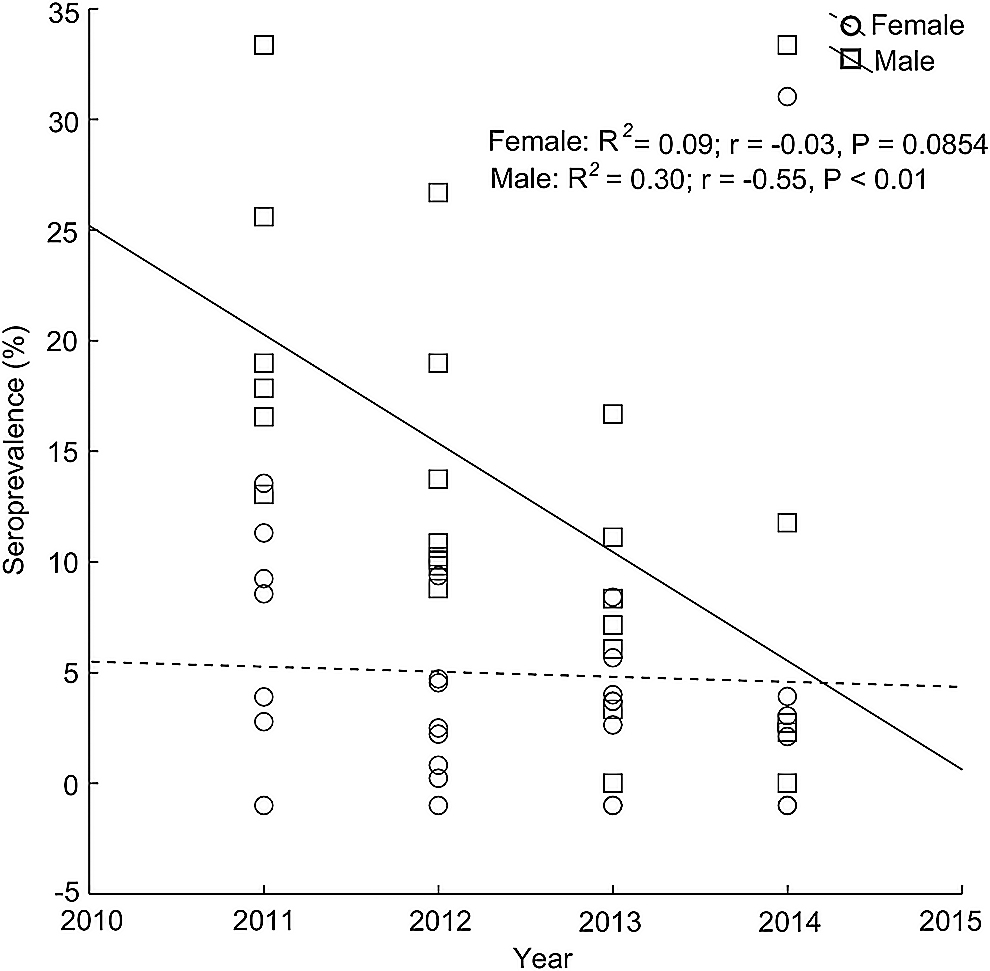
Annual changes of male and female hantavirus seroprevalence.
Based on the capture–recapture data, the seasonal frequency of seroconversion is presented in Table 3. Most rodents were trapped just once (1746/2491), the other 745 individuals were recaptured two or more times. Seroconversion was detected in 58 cases (Apodemus spp.: 56, M. glareolus: 2). The difference in distribution of seroconversion was not significant between seasons (Kruskal–Wallis test: H(2, 29) = 1.39; p = 0.497), although the amount of seroconversion was the largest in summer (Fig. 6A). In the comparison of years, a significant difference was observed [H(3, 29) = 15.17; p < 0.01]: the number of seroconversion events was significantly lower in 2013 when compared with 2011 (post hoc Dunn's test: z = 2.81, p < 0.05) and 2012 (z = 3.48, p < 0.01) (Fig. 6B). The significant difference of seroconversion in 2013 was due to the single detection in the case of the bank vole.
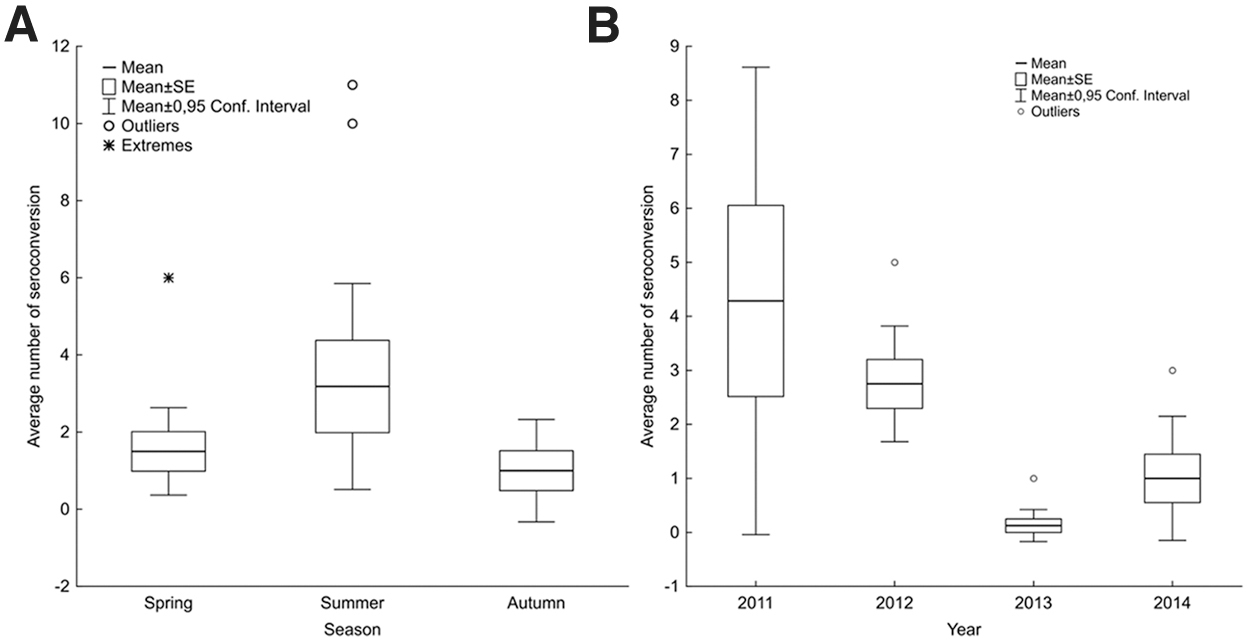
Seasonal
Number of Hantavirus Seroconversion Cases per Season Among Apodemus spp. and Myodes glareolus
There were no sex differences among seroconverted rodents (25 males and 33 females) (χ 2 = 1.1; df = 1; p = 0.29). Seroreversion was observed in case of three A. flavicollis rodents. Two of the specimens were already seropositive at the first capture and became seronegative 13–15 months later. The third rodent was identified as seronegative when first captured, and then 1 month later, it was shown to be seropositive, while 12 months later, the specimen was again seronegative.
Of the rodents captured in the autumn, 9.1% (41/451) were recaptured next spring (Table 4). The winter survival of Myodes was lower when compared with that of Apodemus; however, this observation may be due to a diminished number of captured specimens. Distinctly, there was a sufficient amount of data acquired in only 1 year of the study, in which none of the hantavirus-infected animals survived the winter, while 3.4% of the seronegative individuals were recaptured next spring. There was no significant difference in winter survival among male and female voles (χ 2 = 3.38; df = 1; p = 0.066). In the case of Apodemus, a significant difference between the hantavirus-seronegative and -seropositive individuals was observed in 2011 (χ 2 = 33.3; df = 1; p < 0.001), while in 2012 and 2013, the number of infected and noninfected mice was nearly equal.
Rodent Survival from Autumn (September and October) to Spring (March–May) in Relation to Apodemus and Myodes Hantavirus Status and Sex in the Winter of Every Year
Among seronegative rodents, there was no significant difference between males and females (χ 2 = 1.11; df = 1; p = 0.29), while the prevalence of seropositive males that survived the winter was significantly higher when compared with females (χ 2 = 7.1; df = 1; p < 0.01). The total prevalence values of animals that survived the winters of 2011 and 2012 were nearly equal, 5.9% (1/17) and 5.3% (20/378), while following the winter of 2013, it was significantly higher, reaching 35.7% (20/56) (χ 2 = 21.35; df = 1; p < 0.001) (Table 4).
Discussion
There are only a handful of long-term studies that focus on the simultaneous prevalence of DOBV and PUUV in rodents monitored continually in a well-defined sampling area. In contrast, several studies determined the dynamics of hantavirus infection in Northern and Western Europe (Kallio et al. 2009, 2010, Dobly et al. 2012b, Weber de Melo et al. 2015, Voutilainen et al. 2016, Reil et al. 2017). Although it is well known that hantaviruses are present in Central Europe, studies specifically targeting infection dynamics in this area are completely absent to date. For this reason, we performed a 4-year-long transmission dynamics study to investigate infection changes of hantaviruses among rodent populations and determine those influencing factors that may likely play an important role in these changes.
In a Polish study, hantavirus seroprevalence was also determined (Sadkowska-Todys et al. 2015). Fifty-five rodents of the 564 tested animals were found to be positive for anti-hantavirus antibodies. Similarly, much like our study, the predominant species was A. flavicollis (854/592). Admittedly, their sustained research and the data compiled are not ideally comparable with ours due to the different trapping period and techniques applied. In two Belgian studies, two rodent species (M. glareolus and A. sylvaticus) were examined (Escutenaire et al. 2000, Dobly et al. 2012b). In both cases, researchers implied that the bank vole was the dominant species. Although PUUV seroprevalence fluctuated, it was still significantly higher when compared with DOBV positivity during the period of investigation. Contrarily, in our study, Apodemus spp. were dominant throughout the entire period. From another perspective, we could not observe a positive relationship between prevalence and host density; moreover, the seroprevalence among mice decreased continuously, while among voles, it increased constantly, independently of the rodent density (Fig. 4).
In several other studies, only one rodent species was tested for anti-hantavirus antibodies and for a longer duration. Finnish and German researchers investigated PUUV infection among rodents in endemic areas. In the study by Weber de Melo et al. (2015), PUUV seroprevalence was not dependent on host abundance during the 8-year period. The Finnish 6-year study (Voutilainen et al. 2016) described a cyclic fluctuation in host abundance; therefore, the transmission dynamics followed the population dynamics. In a review (Khalil et al. 2014), authors clearly identified distinctive factors that may likely modify the prevalence of the virus: sex, age, and density were the most important characteristics, among others.
The hantavirus infection rate is usually higher among males than females, likely due to sex-specific behavior, such as aggressiveness and biting (Escutenaire et al. 2002). We observed the same distribution in the first 3 years, although infectivity between the two sexes was equal in the fourth year.
The presence of age-dependent maternal antibodies is crucial from the aspect of susceptibility regarding hantavirus infection seen among young rodents; therefore, age can be an important risk factor. Hence, determination of the host's age is an important issue for correct interpretation of seroprevalence results. Most virological and ecological studies determine the age of animals based on their body weight, which implies that seropositive rodents with a lower weight may likely contain maternal antibodies. The age determination method applied in our study aligned with the recapture data and provided a sufficient base to clearly distinguish maternal antibodies from the natural immune response. In some cases, we identified hantavirus-infected seropositive adults even if their body weight was lower. These results suggested that a method based on body weight measurement is not substantial to determine the presence of age-dependent maternal antibodies correctly.
Numerous studies described the seasonal changes of hantavirus seroprevalence in rodents. Studies from Belgium and Germany described higher hantavirus seroprevalence when bank vole abundance was high in the spring season (Dobly et al. 2012b, Reil et al. 2017). One explanation researchers provided was the presence of overwintered old bank voles in the spring, which were more likely to be infected. Reil et al. (2017) reported similar findings; hantavirus seroprevalence was significantly higher in spring when the abundance of bank voles was high.
In contrast, in a 3-year-long Belgian study, researchers found that hantavirus seroprevalence was high in the autumn of the first year (20.1%), followed by a reduction in the second year, and a rise in the spring of the third year (Escutenaire et al. 2000). Although there were no considerable differences between seasons within years, we identified significant differences between the same seasons of consecutive years (Table 2). Hantavirus seroprevalence among Apodemus mice was elevated in the autumn of 2011 and had plunged in the autumn of 2013. Among Myodes voles, the highest seroprevalence was observed in the spring of 2014.
The 4-year analysis of seroconversion based on monthly data showed that hantavirus infection was higher in summer, yet a significant difference was not observed (Fig. 6). Similar to our findings, in consideration of the study by Bernshtein et al. (1999), the most seroconversions were recorded during the breeding season; however, they found more seroconversion cases among males, which was not observed in our results.
Winter survival was detected among 41 cases during the period examined. Although data culminated from voles were limited, a lower surviving prevalence was observed among hantavirus-seropositive rodents that survived the winter. Kallio et al. reported similar findings in their study (Kallio et al. 2007). Among Apodemus mice, we could not detect a similar difference, likely due to the fact that the prevalence of winter survivors was higher or nearly the same among seropositive mice. Furthermore, winter survival regarding seropositive male mice was higher, while in case of seronegative mice, prevalence was higher among surviving females.
In our study, we ushered in new results regarding seroprevalence, temporal dynamics, and risk factors for two different hantavirus species in a given area, characteristics of which (as of yet) have not been pursued throughout Central Europe. In close connection with these results, we also determined the abundance of host rodents. Significant differences between the numbers of infected males and females were detected in the first 2 years of our study. However, infectivity between the two sexes was equal in the third and fourth years, which surfaces the question whether there is infection dominance among male individuals. Although in most studies the relevance of spring and autumn seasonality was indeed proven regarding viral infection, we have not been able to confirm this specific aspect in our present investigation. Finally, we have pointed out that hantavirus seroprevalence is not clearly dependent upon host abundance, thus we cannot state that high rodent density increases the risk of hantavirus infection in their animal hosts.
Footnotes
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This research was funded by the Hungarian Scientific Research Fund OTKA KH129599. The project was supported by the European Union and cofinanced by the European Social Fund: Comprehensive Development for Implementing Smart Specialization Strategies at the University of Pécs (EFOP-3.6.1.-16-2016-00004), and by the University of Pécs within the Viral Pathogenesis Talent Centre program. The research was financed by the Higher Education Institutional Excellence Program of the Ministry for Innovation and Technology in Hungary, within the framework of the “Innovation for a sustainable life and environment” thematic program of the University of Pécs (TUDFO/47138/2019-ITM). G.K. was supported by the János Bolyai Research Scholarship of the Hungarian Academy of Sciences.