
Abstract
The Japanese encephalitis virus (JEV), a mosquito-borne zoonotic pathogen, is the major cause of viral encephalitis worldwide. An investigation of mosquito species diversity, JEV infection rate, and seasonal population fluctuations of Culex tritaeniorhynchus in Huaihua County, Hunan Province, China, revealed the distribution of vector mosquito populations and genotypes and molecular characteristics of current, common JEV strains in this region. Research on mosquito species diversity in different habitats in Huaihua revealed that local community composition was relatively simple, including five species from four genera (two Culex spp., one Anopheles sp., one Aedes sp., and one Armigeres sp.). Cx. tritaeniorhynchus was clearly the dominant species comprising 94.2–98.6% of all specimens and was always the most common species captured in paddy fields, pigpens, and human dwellings. The seasonal abundance of Cx. tritaeniorhynchus was relatively even, with a single seasonal peak in late August. Two Huaihua JEV strains isolated from the mosquito and pig were highly congruent. The genetic affinities were determined by analyzing capsid/premembrane (C/PrM) and envelope (E) gene variation. The results showed that they were of genotype III and most closely related to the live, attenuated vaccine strains SAl4-14-2 and SA14 and JaGAr01. The Huaihua E protein shares high similarity (mosquito 98.8% and pig 97.6%) at the nucleotide level with the SA14-14-2 vaccine. Although we found that the E gene sequences of the Huaihua JEV mosquito strain and pig strain have 11 and 15 amino acid site substitutions compared with the SAl4-14-2 vaccine, key sites that associated with JEV's antigenic activity and neurovirulence were unchanged. The SA14-14-2 vaccine should therefore be effective in preventing JEV infection in the Huaihua region.
Introduction
Japanese encephalitis virus (JEV) is a major cause of viral encephalitis. JEV infection can also lead to acute encephalitis in humans, of which ∼25% to 30% cases are fatal and 50% result in permanent neuropsychiatric sequelae (Banerjee et al. 2017). JEV is mainly prevalent in the East, South Asia, and Western Pacific regions, including Australia (van den Hurk et al. 2012). JEV exists in a zoonotic cycle of transmission between mosquitoes and amplifying hosts such as pigs and wading birds (Le Flohic et al. 2013). Of these, pigs are considered the most important natural amplifying hosts.
JEV belongs to the genus Flavivirus of the family Flaviviridae and its genome consists of a single-stranded positive-sense RNA ∼11 kb in length. Based on the diversity of nucleotide sequences of the E protein gene or the gene encoding the complete polyprotein, JEV can be classified into five genotypes (I–V) (Solomon et al. 2003).
Although vaccination has now dramatically reduced case numbers, JEV remains a major public health problem (Hegde and Gore 2017). It is reported that JEV is prevalent throughout China, even including high altitude regions in Tibet (Zhang et al. 2017). Huaihua County (N27°48′09.57″, E109°95′39.61″) is the biggest county in Hunan Province that has been constantly suffering from JEV since the 1950s (Guo and Li 2002). Local subtropical monsoon climate provides warm sunny climate and abundant rainfall for Culex tritaeniorhynchus (Giles) with suitable breeding conditions. Detailed investigations of basic aspects of the ecology of Cx. tritaeniorhynchus such as population density and seasonal population fluctuations have yet to be done. Moreover, it remains unknown if Cx. tritaeniorhynchus is still the dominant species after ongoing urbanization and development in the Huaihua region, with gradual reduction in rice cultivation and standardization of pig husbandry. In addition, the lack of further research on common JEV strains in this region limits our current understanding of local JEV genotypes and their etiology. For example, China's JEV monitoring database shows a gradual replacement of the JEV Type III genotype by the Type I genotype over the past 30 years (Wang et al. 2007); whether the local JEV genotype in Huaihua region followed the national trend remains unclear.
We here present results of an investigation of mosquito species diversity—seasonal population fluctuations of Cx. tritaeniorhynchus in Huaihua County. In addition, the JEV genotypes and infection rates in vectors and pigs were investigated. Our results may have potential implications for prediction and prevention of JEV outbreaks in this region.
Materials and Methods
Study site
Fieldwork was conducted in Huaihua County (N27°48′09.57″, E109°95′39.61″), Hunan Province, China. Mosquitoes were collected from July to September in 2012 and 2013 at sites located in the east, southwest, and north of Huaihua County, namely Chihui Village (N27°48′09.57″, E109°95′39.61″), Lutianchong Village (N27°58′1.82″, E109°98′3.93″), Aitouping Village (N27°56′7.78″, E110°0′11.06″), and Shunxipu Village (N27°54′8.59″, E109°91′0.99″). Three habitat types representative of the most common village habitats were selected for mosquito sampling in each village: Habitat A, rural environment with paddy fields; Habitat B, rural environment with domestic animals, including pigs, ducks, hens, and cattle (except for cattle, other domestic animal species are kept inside at night, usually in semiclosed shelters); and Habitat C, human dwellings. The distance between sampling sites was about 2 km.
Mosquito and pig brain sample collection
Mosquitoes were trapped daily at every sampling site from late July to early September (local mosquito activity season) in 2012 and 2013. Light traps were set daily at a height of 1.0 meters above the ground from 6:00 pm to 8:30 pm (local time) with CO2 CDC light traps for Habitat A, animal-baited light traps for habitat B, and human-baited light traps for habitat C. Three light traps were used simultaneously at one sampling site. Pig brain samples were collected from local slaughterhouses. Samples were transferred into 1.5-mL tubes, which were labeled with the date and collection site before being frozen in liquid nitrogen and transported to the laboratory. Captured mosquitoes were anesthetized, identified to sex and species on the basis of morphology, and counted (Lu 1997). Females were pooled in 1.5-mL tubes with 30–50 individuals per pool and stored at −80°C.
Data analyses
The proportion of positive pools was calculated by the number of infected pools to the total number of pools tested. The minimum infection rate (MIR) is calculated as the ratio of the number of positive pools to the total number of mosquitoes tested.
The Berger–Parker dominance index, D = N max/N, where N max is the number of the dominant species and N the total number of all specimens, was used to provide an index of overall dominance. We used standard criteria to rank the relative dominance of various species; dominant species were those for which D ≥ 0. 1, abundant species were those where D was in the range 0.05 ≤ D < 0.1, common species were those for which D = 0.01 − 0.05, occasional species were those for which D = 0.001 − 0.01, and rare species were those for which D < 0.001 (Berger and Parker 1970).
The Shannon–Wiener diversity index, 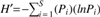 , where S is the number of species and Pi
the proportion of individuals of each species, was used to calculate an index of species diversity in different habitats (Spellerberg and Fedor 2003).
, where S is the number of species and Pi
the proportion of individuals of each species, was used to calculate an index of species diversity in different habitats (Spellerberg and Fedor 2003).
Pielou's evenness index formula, J = H′/lnS, where J is index of the degree of homogeneity and S is the number of species, was used to calculate an index of the numerical equality between species in different months and habitats (Pielou 1966).
Simpson's formula, 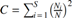 the number of individuals of each species, and N the total number of individuals of all species, was used to estimate the ecological dominance of mosquito species in different habitats at different times of the year (Simpson 1949).
the number of individuals of each species, and N the total number of individuals of all species, was used to estimate the ecological dominance of mosquito species in different habitats at different times of the year (Simpson 1949).
JEV detection and genome sequencing
Total RNA was extracted from original biological samples or cell culture suspensions using TRIzol reagent (Invitrogen) with slight modifications. RNA extraction was washed with double-distilled water, treated with diethyl pyrocarbonate (DEPC), transferred into a sterile glass tissue grinder, and triturated in 2 mL of TRIzol reagent. The mixture was allowed to stand for 10 min at room temperature before ∼1/5 of the volume of chloroform was added, after which the mixture was shaken. The resulting RNA pellet was washed with 75% ethanol. Each viral RNA extract was transcribed into complementary DNA (cDNA) using an RNA PCR kit (AMV ver. 2.1; Takara Biotechnology, Dalian, China Co. Ltd.). The resulting cDNA was used as the template for subsequent PCR reactions. Whole genomes were amplified using primer sets described in Table 1. PCR products were recovered using a gel purification kit (Qiagen, Valencia, CA). DNA sequencing was performed on an Applied Biosystem 377 automatic sequencer (Takara Biotechnology, Dalian, China Co. Ltd.).
Primers Used for RT-PCR Assay to Detect the Complete Genome of Japanese Encephalitis Virus in Mosquitoes
Sequence alignment and phylogenetic analysis
The evolutionary relationships of the capsid/premembrane (C/PrM) and envelope (E) genes were compared, respectively, with 2 Huaihua mosquito and pig isolated JEV strains and 31 references of JEV in GenBank from different decades, countries, and hosts (Table 2). Using Murray Valley encephalitis virus as an out-group, we created a neighbor-joining (NJ) tree with a bootstrapping resampling process (1000 replicates) by MEGA software, version 6.0. Differences in amino acids were analyzed by GENEDOC software, version 2.7. The genotype was decided by the method established by Woan-Ru Chen (Chen et al. 1990, 1992).
Background Information on 31 Japanese Encephalitis Virus Strains Compared with Huaihua Japanese Encephalitis Virus Isolates in This Study
CSF, cerebro-spinal fluid.
Results
Species dominance and diversity
A total of 33,516 mosquito specimens representing 5 species from 4 genera (2 Culex spp., 1 Anopheles sp., 1 Aedes sp., and 1 Armigeres sp.) were collected in the 2-year study. The dominance index reflects differences in the abundance of individual species, the higher the dominance index, the more uneven the distribution of individual species and the more obvious the dominant species are. As shown in Table 3, in the year of 2012, the local dominant species was Cx. tritaeniorhynchus (Berger–Parker dominance index, D = 0.9828). All other species were much less abundant; Anopheles sinensis (Wiedemann) was a common species with D = 0.0127 and Armigeres subalbatus (Coquillett) was an occasional species with D = 0.0034, while Culex pipiens quinquefasciatus (Say) and Aedes albopictus (Skuse) were locally rare with D < 0.001, respectively. The species constitution in 2013 was similar to the one in 2012 except for the absence of Ae. albopictus.
Relative Dominance of Mosquito Species Collected in Different Habitat Types in Huaihua, Hunan Province, China, in 2012 and 2013
The species distribution patterns were consistent in different habitat types, the abundance of Cx. tritaeniorhynchus was so high (94.2–98.6% of all specimens) that it was clearly the dominant species in all three habitat types. The diversity index, evenness index, and dominant concentration index (Table 4) were very similar in paddy fields, hog pens, and human dwellings in both 2012 and 2013, indicating that species diversity was relatively low.
Mosquito Species Diversity, Evenness, and Dominant Concentration Index at Huaihua, Hunan Province, China, in 2012 and 2013
Cx. tritaeniorhynchus density was high in paddy fields, moderate in human dwellings, and low in pigpens. Abundance was relatively constant from July to mid-August with a peak in late August, followed by a decline in early September with the gradual decrease in temperature (Fig. 1).

Seasonal trends in abundance of Culex tritaeniorhynchus in different habitat types in Huaihua, Hunan Province, China.
JEV detection and genome sequencing
A total of 18,957 Cx. tritaeniorhynchus mosquitoes were captured in 2012 and 13,492 were captured in 2013, which were pooled into 501 and 295 samples (30–50 individuals of each pool), respectively. Five pooled samples from 2012 and six from 2013 were PCR positive for JEV. MIRs for 2012 and 2013 were 0.26% and 0.44%, respectively. There was no significant difference between the three habitats. In addition, 22 of 134 (16.4%) pig brain samples were PCR positive.
Two isolations of the virus were obtained from the PCR-positive sample by inoculating mosquito and pig brain homogenate supernatants into monolayers of C6/36 cells. One complete virus genome from the mosquito of 10,381 nt was sequenced (GenBank acc. no. KU904395). Two C/PrM genes and two E gene sequences obtained from the mosquito and pig were 907 and 1500 nt long, respectively, and have been deposited in GenBank (acc. no. KM897136 and KM897135 for mosquito and MH359234 and MH359235 for pig).
Analysis of C/PrM and E gene nucleotide and amino acid homology
Phylogenetic tree topologies based on C/PrM and E gene variation are highly congruent (Figs. 2 and 3). The C/PrM and E gene sequence identities between two strains isolated from the mosquito and pig were 97.2% and 99.8%, respectively. Huaihua JEV strains from the mosquito and pig were most closely related to the live, attenuated JEV vaccine strains SAl4-14-2, virulent parental strain SA14, and Japan isolation JaGAr01, clustering within genotype III.

Phylogenetic tree of relationships between JEV strains based on C/PrM gene variation. Full nucleotide sequences of different JEV genomes were aligned and evolutionary analyses were conducted in MEGA 6.0 using 1000 bootstrap replicates of the sequence data. The MVE-51 virus was used as an out-group. The bar indicates the phylogenetic distance associated with 0.1 amino acid substitutions per site. C/PrM, capsid/premembrane; JEV, Japanese encephalitis virus.

Phylogenetic tree of relationships between JEV strains based on E gene variation. Complete nucleotide sequences of different JEV genomes were aligned and evolutionary analyses were conducted in MEGA 6.0 using 1000 bootstrap replicates of the sequence data. MVE-51 virus was used as an out-group. The bar indicates the phylogenetic distance associated with 0.1 amino acid substitutions per site. E, envelope.
For the JEV mosquito strain, levels of C/PrM gene nucleotide and amino acid homology with the SAl4-14-2 vaccine were 99.7% and 99.99% and levels of E homology with the vaccine strain were 98.8% and 99.98%, respectively. For the JEV pig strain, levels of C/PrM gene nucleotide and amino acid homology with the SAl4-14-2 vaccine were 99.1% and 97.2% and levels of E homology with the vaccine strain were 97.6% and 99.97%, respectively.
Sequencing results indicate that C/PrM nucleotide and amino acid homology between the Huaihua JEV mosquito strain and 31 reference strains ranged from 79.8% to 99.9% and 54.6% to 100%, respectively, while the Huaihua JEV pig strain and reference strains were 81.1–99.9% and 59.4–99.9%. There was no significant difference in the nucleotide sequence between the strains isolated in different years, such as there was only 5.2% difference between the Beijing-1 strain in 1949 and the Huaihua mosquito strain.
Levels of E gene nucleotide and amino acid homology between the mosquito strain and 31 references were 40.8–99.4% and 15.9–99.99%, while they were 54.2–100% and 14.2–99.0% for the pig strain and references.
Analysis of mutation at critical amino acid residues
Because the JEV E gene is the main gene involved in protective antigenic activity, any amino acid differences with current vaccines will directly affect a vaccine's ability to confer immunity against the JEV. We compared amino acid sequences of the Huaihua JEV mosquito strain (KM897135) and pig strain (MH359235) with that of the live vaccine strain SA14-14-2 and other similar strains (Table 5). We found 11 substitutions among the 500 E protein amino acid residues in the mosquito strain (Appendix Fig. A1). These included sites E138, E176, and E177 in E protein Domain I, E107, E244, E264, and E279 in Domain II, and E315 in Domain III. There were also substitutions at other loci outside the active region (E415, E439, and E447). Besides the sites mentioned above, there were 4 additional substitutions, including E123, E209, E340, and E459, in the Huaihua JEV pig strain. However, some key sites that affect virulence and antigenic activity were unchanged compared with other JEV Type III genotype strains.
Comparison of Key Japanese Encephalitis Virus E Protein Amino Acid Residues at Sites Associated with Virulence in Different Japanese Encephalitis Virus Strains
JEV, Japanese encephalitis virus.
Discussion
JEV is active in all of the Chinese provinces except Qinghai, and nearly 50% of the world's reported cases occur in China (Zheng et al. 2012). Numerous JEV strains have been isolated from mosquitoes, pigs, and human patients in different areas in China (Li et al. 2017, Xia et al. 2018), similar to Uttar Pradesh, India, which suffered the rapid dissemination of JEV within a population, facilitated by different species of Culex (Nyari et al. 2015). Huaihua County locates in central China with subtropical monsoon climate. Abundant sunlight, warmth, and rain make it an ideal place for crop growing and pig raising. Cx. tritaeniorhynchus and pigs played critical roles in JEV transmission. Therefore, research on the local seasonal population dynamics of Cx. tritaeniorhynchus, the JEV genotype, and the natural JEV transmission cycle is important for understanding the transmission and spread of JEV in the Huaihua region. Detection of JEV in mosquitoes and pigs around paddy fields and pigpens confirmed that there is a risk of JEV infection in this area.
Cx. tritaeniorhynchus predominantly rests on pig farms and human dwellings and was the main nocturnal mosquito in the Huaihua region. Its broad distribution, seasonal peak in abundance, and blood feeding mainly on animals and occasionally on humans (Philip Samuel et al. 2008, Tuno et al. 2017) make it an ideal combination for transmission of JEV to humans as the most important local vector. Adult Cx. tritaeniorhynchus presented over a relatively long period each year with a peak of abundance in late August, then began to decline in early September in relation to seasonal changes in rice crop irrigation, which is consistent with an entomological study conducted in neighboring country India (Kanojia et al. 2003).
The minimum JEV infection rates in mosquitoes detected in Huaihua were low (0.26% in 2012 and 0.45% in 2013), while the pig infection rate was 16.4%, which could be explained by the pigs' virus-amplifying work, the majority mosquitoes captured from paddy fields instead of pigpens, or the possible existence of direct transmission of JEV between pigs without mosquitoes (Ricklin et al. 2016).
A previous study partially sequenced the C/PrM and E gene regions of available JEV strains, both of which are important for induction of protective immunity (Ni and Barrett 1995). Viral E proteins have been demonstrated to be reliable phylogenetic markers and are one of the major structural proteins that play an important role in immunogenicity, tissue tropism, cell fusion and infection, and virus maturation (Monath et al. 2002, Gao et al. 2015). We found that both C/PrM and E gene variations indicate that the Huaihua JEV strain is of the Type III JEV genotype. The sequence identities between Huaihua JEV mosquito and pig strains of the C/PrM and E gene were 97.2% and 99.8%, respectively, confirming the local natural JEV transmission cycle within mosquitoes and pigs.
The three-dimensional structure of the JEV envelope glycoprotein comprises chiefly three domains; Domain I (E1–E51, E137–E196, and E293–E311), Domain II (E52–E137 and E197–E292), and Domain III (E310–E411). Domains fold independently onto the surfaces of viruses to form immunoglobulin-like structures, and because of this conformation, many antigen-neutralizing epitopes are concentrated in this domain (Kolaskar and Kulkarni-Kale 1999, Luca et al. 2012). The E protein's Domain III is the main epitope (also called the antigenic determinant), the key sites of which are concentrated in the three regions between E337–345, E377–382, and E397–403 (Wu and Lin 2001). Compared with the vaccine strain, there was only one change at E340 in the Huaihua pig strain, which was totally identical in the Huaihua mosquito strain, without changing the antigenic activity by influencing the binding reaction between antigens and neutralizing antibodies.
The disulfide bond formed by cysteine at E304 and E335 is the necessary structure for binding of antigens and antibodies (Banerjee et al. 2017). His at E264 strengthens a hydrogen-bonding network, leading to a more stable virus (Wang et al. 2017). E222 and E327 are potential genotype-related neutralization determinants and are critical in determining the protective efficacy of the live Japanese encephalitis vaccine SA14-14-2 (Ye et al. 2015). Mutations at E306, E331, E387 (Konishi et al. 1999), and E333 (Wu et al. 1997) sites may be related to the JEV neutralization activity and antigenic determinants. N103K might act as the initiator for transition of genotype III to genotype I (Banerjee et al. 2017). In summary, there was no mutation at the sites mentioned above in the Huaihua JEV strains.
Comparison of sequence data revealed that although the Huaihua JEV mosquito/pig strains and the SA-14-14-2 vaccine differed at 11/15 amino acid sites, key regions that determine antigenic activity were identical. This suggests that the Huaihua JEV strain's antigenicity is the same as the SA-14-14-2 vaccine strain. In theory, the currently used SA-14-14-2 vaccine should be effective in protecting against infection by the Huaihua JEV strain.
In general, the sequence of E protein of virulent isolate of JEV was compared with that of nonvirulent SA14-14-2 to understand neurovirulence and neuroinvasiveness effects. Comparison of the Huaihua mosquito strain and SAl4-14-2 vaccine strain E gene amino acid sequences indicates 11 mutations, including those at loci E138, E176, and E177 (Domain I); E107, E244, E264, and E279 (Domain II); E315 (Domain III); and E415, E439, and E447 outside the active region. Besides the sites mentioned above, there were 4 additional substitutions, including E123, E209, E340, and E459, between the Huaihua pig strain and SA14-14-2 vaccine strain. While high neurovirulence of JEV isolates was reported to be contributed by eight mutations (F107L, K138E, V176I, A177T, H264Q, M279K, V315A, and R439K) in E protein (Banerjee et al. 2017), in which mutations at the E107 or E138 position significantly affect virulence, the other mutations exhibited significant increases in neurovirulence only when combined with E138, E107, and other mutations (Yang et al. 2017). Evidence showed that a single I176R mutation could significantly decrease JEV neurovirulence in mice (Zhou et al. 2018). Of these, the presence of glutamate at E138 and Leucine suggests that the Huaihua strain is more virulent compared with the vaccine strain. Amino acids at the above key E gene sites are the same in the Huaihua JEV strains as in other JEV Type III genotype strains, indicating that no neurovirulence changed.
Our analysis of C/PrM and E gene nucleotide sequence variation indicates that despite differences in time of collection, hosts, and geographic location, both the Huaihua JEV mosquito and pig strains have a relatively high degree of homology with other genotype III JEV strains from different countries and regions, especially India, where northern Indian states have many similarities to China in ecology (Samuel et al. 2009). Nucleotide differences could possibly reflect differences in habitat and vector biology. On the other hand, key E gene sites of the Huaihua strain responsible for antigenic activity and virulence were identical to those of other genotype III JEV strains, which suggests that JEV is genetically relatively stable.
Until the late 20th century, studies indicated that the predominant genotype was genotype III. Since then, there have been multiple reports of genotype I displacing genotype III in many regions (Wang et al. 2007, Li et al. 2011, Chen and Yu 2013, Gao et al. 2015), and in many areas, genotype I is now recognized as the dominant strain. This study reported on the vector mosquito ecology and that genotype III was still the dominant strain in Huaihua. The result provided a scientific basis for forecasting and providing early warnings of JEV outbreaks in Hunan Province. Therefore, it will require implementing long-term continuous monitoring of JEV-infected mosquitoes and obtaining additional relevant information on the biological characteristics of JEV in this region.
Footnotes
Acknowledgments
This work was funded by grants from the Infectious Diseases Prevention and Cure Project of China (no. 2012ZX10004219) and National Science and Technology Support Program of China (2014BAI13B01).
Authors' Contributions
C.C. and T.Z. carried out the molecular genetic studies, participated in the sequence alignment, and drafted the manuscript. The experiments were conceived and designed by C.C., T.Z., Y.J., C.L., Y.D., X.G., D.X., and T.Z.; experiments were performed by C.C., T.Z., Y.J., G.W., and X.G.; and data were analyzed by C.C., T.Z., X.G., and G.W. All authors read and approved the final manuscript.
Author Disclosure Statement
No competing financial interests exist.