
Abstract
An increase in zoonotic infections in humans in recent years has led to a high level of public interest. However, the extent of infestation of free-living small mammals with pathogens and especially parasites is not well understood. This pilot study was carried out within the framework of the “Rodent-borne pathogens” network to identify zoonotic parasites in small mammals in Germany. From 2008 to 2009, 111 small mammals of 8 rodent and 5 insectivore species were collected. Feces and intestine samples from every mammal were examined microscopically for the presence of intestinal parasites by using Telemann concentration for worm eggs, Kinyoun staining for coccidia, and Heidenhain staining for other protozoa. Adult helminths were additionally stained with carmine acid for species determination. Eleven different helminth species, five coccidians, and three other protozoa species were detected. Simultaneous infection of one host by different helminths was common. Hymenolepis spp. (20.7%) were the most common zoonotic helminths in the investigated hosts. Coccidia, including Eimeria spp. (30.6%), Cryptosporidium spp. (17.1%), and Sarcocystis spp. (17.1%), were present in 40.5% of the feces samples of small mammals. Protozoa, such as Giardia spp. and amoebae, were rarely detected, most likely because of the repeated freeze-thawing of the samples during preparation. The zoonotic pathogens detected in this pilot study may be potentially transmitted to humans by drinking water, smear infection, and airborne transmission.
Introduction
An increase in zoonotic infections in humans has led to a high level of public interest in recent years (Hadfield et al. 2011). Although a remarkable number of zoonoses are of parasitic origin and one of the hot spots is Western Europe (Jones et al. 2008, Hadfield et al. 2011), data pertaining to the distribution of zoonotic parasites in domestic and wild animals are surprisingly rare (Klimpel et al. 2007).
Rodents and other small mammals play a significant role in the spread of about 850 species of zoonotic pathogens (Paramasvaran et al. 2009). Five percent of these pathogens are protozoa, and 32% are helminths (Taylor et al. 2001). Among the emerging infectious diseases, 10.7% and 3.3% are caused by protozoa and helminth infections, respectively. However, Cryptosporidium spp., Cyclospora spp., and microsporidia, which are currently classified as either parasitical or fungal organisms, also play a role (Abrahamsen et al. 2004, Jones et al. 2008, National Institutes of Health 2012, Guo et al. 2015, Khan et al. 2017).
The emergence of zoonotic helminth infections is associated with a number of factors: (1) changes in social, dietary, and cultural structures with increased human exposure to zoonotic helminths; (2) environmental changes; and (3) improved recognition and/or diagnostic methods of these neglected infections (McCarthy and Moore 2000). Decreasing biodiversity and rapidly increasing human population density have also been proposed as reasons for increased pathogen transmission to humans (Keesing et al. 2010). But also augmented human mobility can cause an increase and the spread of small mammal populations and therefore favor the dissemination of ecto- and endoparasites and other infectious pathogens (Antoniou et al. 2010). Transmission to domestic animals has already been described. The mechanisms of transmission to humans are, however, less explored (Klimpel et al. 2007).
Previous studies of the network “Rodent-borne Pathogens” focused on viruses and bacteria (Nainys et al. 2015, Heuser et al. 2017, Mrochen et al. 2017, Fischer et al. 2018). The aim of this study was to reveal the prevalence of endemic parasites with zoonotic potential in small mammals sampled at seven sites in Germany.
Materials and Methods
Small mammal collection
Small mammals were collected at seven sites (Fig. 1) from May 2008 to June 2009 and necropsied within the framework of the activities of the “Rodent-borne pathogens” network (Ulrich et al. 2008, Johne et al. 2010). All the 111 sampled free-living wild small mammals were identified to species level morphologically and by cytochrome b gene analysis (Schlegel et al. 2012). In addition, sex, body mass, and tail length were recorded. Intestine samples were obtained and stored in sterile plastic tubes at −20°C until analysis.

Sampling sites of small mammals in Germany.
Macroscopic examination, staining, and concentration methods
During necropsy, the small and large intestines of each animal were excised and stored at −20°C. For the detection of parasites and coccidia, the intestines were thawed, opened longitudinally, and examined macroscopically for the presence of helminths. Afterward, all feces was removed and used for specific staining and concentration methods as follows:
For Heidenhains iron hematoxylene stain, a pea-sized homogenized feces sample was smeared onto a microscope slide and stained as described earlier (Albert 1926) with a modified short protocol (1% hematoxylene solution for 15 min). Five sight field lines in each slide were examined for the presence of intestinal protozoa by using light microscopy with 1,000-fold magnification.
For Kinyoun stain, a pea-sized homogenized feces sample was smeared in a thicker layer onto a microscope slide and stained as described earlier (de Oliveira Lemos et al. 2012), with staining in basic fuchsine solution for 15 min instead of 3 min. After drying, the slide was evaluated for the presence of coccidia (Cryptosporidium spp., Cyclospora spp., and Isospora spp.) by using light microscopy with 1,000-fold magnification.
For the Telemann worm egg concentration method, the remaining feces sample was transferred to 15-mL tubes and treated as described earlier (Telemann 1908). The entire pellet was examined for the presence of helminth eggs and coccidia (Sarcocystis spp. and Eimeria spp.) by using light microscopy at 100- to 200-fold magnification. In addition, the two-layer gauze was examined for the presence of adult worms by using light microscopy with 10- to 25-fold magnification.
All adult worms found during necropsy or Telemann concentration were transferred to 70% ethanol and stained with carmine acid stain (Khrustalev and Hoberg 1996) for macroscopic and microscopic species determination.
Statistical analysis
Statistical analyses were performed by using GraphPad Prism 4.0 and 6.01 (GraphPad Software, San Diego, CA) and SPSS version 20.0. Gender, parasite infestation, infestation with zoonotic parasites, and sampling sites were compared by chi-square test or Fisher's exact test, as appropriate. Differences were considered significant at p ≤ 0.05. Numerical results are given as mean ± standard deviations, unless stated otherwise.
Results
Ninety-nine of 111 investigated small mammals were rodents (order Rodentia) and 12 were insectivores (order Soricomorpha; taxonomy according to Wilson and Reeder 2005). Basic information on the species and family composition is provided in Supplementary Table S1. Fifty-six of the small mammals (50.5%) were females, whereas in three individuals (2.7%) the sex was not determined. Parasite-infested small mammals were detected at all seven sampling sites. In 79 of the 111 (71.1%) small mammals, intestinal parasites were detected, and 38 (34.2%) of them harbored parasites with zoonotic potential. Multiple parasitic infections were detected in 23.4% of the small mammals, though infestations with multiple zoonotic parasites were rare (4.5%; Tables 1 and 2). The proportion of positive small mammals differed significantly between the sampling sites (p = 0.012). Infection rates varied from 16.7% in Hamburg to 53.3% in Lohma. The prevalence of potentially zoonotic parasites ranged from 8.3% in Lucka to 53.3% in Lohma. Differences in prevalence of zoonotic parasites between small mammals' taxonomic family were not detectable (Muridae: 21 of 64, 32.8%; Cricetidae: 12 of 35, 34.3%; Soricidae: 2 of 11, p = 0.585).
Frequency of Intestinal Parasitic Infections by Sampling Site in the 111 Small Mammals Investigated
For localization of sampling sites, see Fig. 1.
Species of Intestinal Parasitic Infections in the Small Mammals
For localization of sampling sites, see Fig. 1.
−, negative samples.
Ten different species of helminths, eight nematodes, and two cestodes were identified. Helminthic infections with zoonotic potential were detected in 30 of 111 (27.0%) small mammals, with Hymenolepis spp. (syn. Rodentolepis spp.) being the most prevalent species (22 of 111 small mammals, 18.0%) at 4 out of 7 sampling sites (Fig. 1 and Tables 1 and 2). Hymenolepis diminuta was found in 18 (16.2%), and Hymenolepis nana was found in 6 (5.4%) small mammals (Table 3). Mixed infections with H. nana and H. diminuta were also present in two samples. Six rodents (5.4%) from five sampling sites were infested with Capillaria spp. (Fig. 2a, c); no insectivore was positive. Six small mammals (5.4%) had a Trichuris spp. infection (Fig. 2b, d). Helminthic infections without zoonotic potential (Aspiculuris tetraptera, Syphacia muris, Syphacia obvelata, Trichostrongylus spp., Nippostrongylus spp., Heligmosomoides spp., Paranocephala spp.) were observed in 21 of 111 (18.9%) small mammals, and 10 of these individuals had double or multiple infections.

Capillaria spp. eggs
Distribution of Hymenolepis spp. in the Small Mammals We Investigated
For localization of sampling sites, see Fig. 1.
Hd, Hymenolepis diminuta; Hn, Hymenolepis nana.
Seven rodents from 3 areas (6.3% of the 111 examined animals) were positive for Giardia spp. (Fig. 3a–e). All insectivores were negative. In three (2.7%) rodents, protozoa without zoonotic potential (two Chilomastix spp., Fig. 3f, and one Entamoeba spp.) were detected.
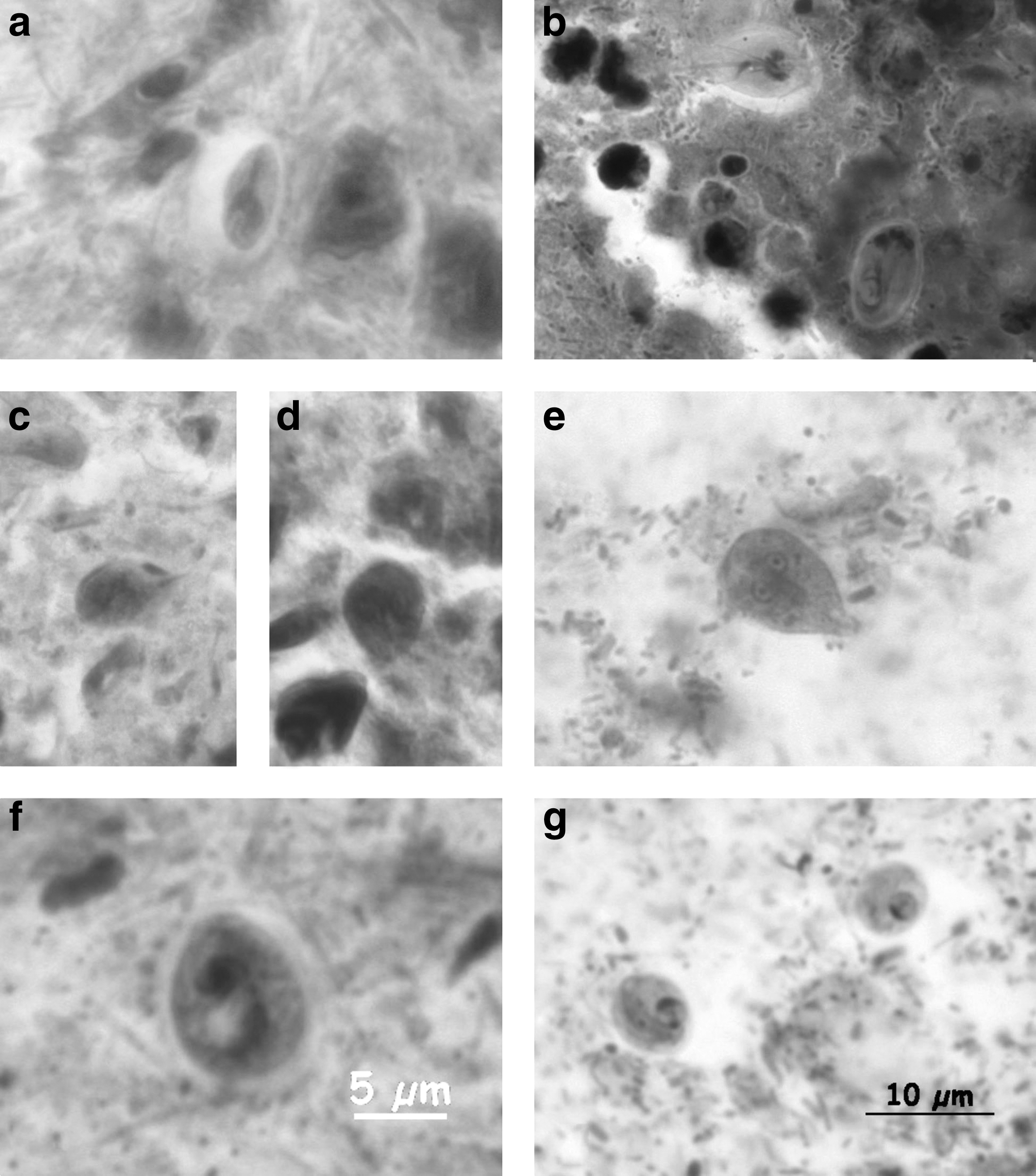
Giardia spp.
Sixty-six (55.6%) small mammals were coccidia-positive, whose distribution did not differ significantly between the mammal families represented (p = 0.089). Eimeria spp. were detected in 34 (30.6%) animals, exclusively in rodents. Both Sarcocystis spp. and Cryptosporidium spp. were present in 18 (16.2%) small mammals. Infections with more than one coccidia species were common (20 of 62 coccidia-positive samples, 32.5%).
In our study, multiple parasitic infections were common in all small mammals, and also when separately analyzed in groups (long-tailed mice, voles, and shrews, p = 0.026), and zoonotic parasites were not significantly more prevalent in any of the groups (p = 0.089). Neither species (p = 0.206) nor gender (p = 0.403) had significant influence on the prevalence of zoonotic parasites.
Discussion
In our study, Hymenolepis spp. was the most prevalent parasite, infesting 17.2% of rodents and 25.0% of insectivores sampled at four of seven sites. These tapeworms were diagnosed either on the basis of worm eggs in feces or on the basis of adult worms in stool debris. The human-pathogenic H. nana cannot be clearly differentiated from H. nana var. fraterna (syn. Rodentolepis fraterna), which is also—or instead—present in small mammals, by the microscopic morphology of their parasite eggs. We did not find adult life stages, so a more clear differentiation was not possible. As seen in the literature, the data are contradictory due to lacking molecular typing results. Some working groups detected all three species in parallel—as described in rats (Rattus rattus and Rattus norvegicus) (Franssen et al. 2016) or did not differentiate it, but discussed it as potentially zoonotic, as in house mice (Mus musculus) and other small mammals (Zarei et al. 2016). In 2016, the mitochondrial genome of H. nana was sequenced by next-generation sequencing for the first time (Cheng et al. 2016). In small mammals, Hymenolepis infections are very common, as shown in the United States (34.4%) and on the Canary Islands (25.4%) (Easterbrook et al. 2008, Foronda et al. 2011). Hymenolepis infections in humans were, depending on the species, caused by the ingestion of eggs, which are transmitted directly via smear infection from subject to subject—as it is common in H. nana (Willcocks et al. 2015, Cheng et al. 2016), or contaminated drinking water or food, or, more rarely, by the ingestion of the cysticercoid larvae found in infected arthropods (Jirovec 1960). Cases of human infections are rare, with only a few hundred cases reported worldwide (Patamia et al. 2010). There have been few cases in Europe: Some were reported in school children from Italy (Lo Nigro et al. 1961), 6 cases from Austria (Tomaso et al. 2001), and 18 cases in the former Czechoslovakia (Rehka 1981), but there are no reports from Germany so far. Despite the fact that parasites are common in rodents and transmission routes are diverse, it is currently not clear why human infections remain rare. Only three hymenolepid species are known to infect humans: the rat tapeworm H. diminuta, the dwarf tapeworm H. nana, and the rodent tapeworm Rodentolepis microstoma (Macnish et al. 2002, 2003). Other Hymenolepis spp. seemed to be rare in humans, or possibly misdiagnosed as H. diminuta or H. nana infections. This may be due to either the difficulties in microscopic differentiation of hymenolepid species or the lack of published gene sequences and established PCR protocols (Casanova et al. 2001, Foronda et al. 2011). Possibly human-pathogenic H. nana and rodent-pathogenic R. fraterna are morphologically similar, but they may be genetically distinct and do not infect the opposite host, but this should be elucidated by genome sequencing.
The flagellate Giardia spp. was detected in only 7.1% of small mammals. Though Heidenhain stain works well in the detection of intestinal protozoa, the very fragile trophozoites and also the more stable cysts were lost or affected by the multiple freeze-thawing procedures in our samples, so that the prevalence of protozoa might actually be higher. Rodent studies in Poland revealed Giardia spp. prevalence rates of 24.4% in yellow-necked field mice, Apodemus flavicollis, 58.3% in bank voles, Myodes glareolus, and 74.2% in common voles, Microtus arvalis (Bajer 2008a). The genetic differentiation between human and zoonotic Giardia spp. is reported only rarely (Thompson 2004, Lebbad et al. 2010, Helmy et al. 2018), and it is not known how many human cases are caused by zoonotic strains. A few studies showed that zoonotic strains may play a role in infections in children (Lebbad et al. 2011). A recent study indicated a frequent detection of Giardia muris and Giardia microti in rodents, but only rare occurrence of zoonotic Giardia duodenalis (Helmy et al. 2018). The German surveillance system records all cases of laboratory or clinically confirmed giardiasis in humans since 2001 with 5,727 infections reported, but without assemblage information (Robert Koch-Institut 2013). Transmission from small mammals to humans might be possible via drinking water from surface water sources or contaminated food (Exner and Gornik 2004, Bajer 2008b). The greatest zoonotic risk is suspected to come from Giardia assemblages found in livestock, but not in free-living wild mammals or their habitats (Thompson 2004, Leonhard et al. 2007).
Intestinal coccidia (including Cryptosporidium spp.) were present in 55.8% of the small mammals without a significant difference in the distribution between the mammal families. Infections with more than one coccidium species were common. Cryptosporidium spp. infections seem to be common in small mammals from Poland (53.8% of M. glareolus, 28.1% of A. flavicollis, and 62.3% of M. arvalis) and Great Britain (Webster 1996, Bajer 2008a) and have occasionally been described in African hedgehogs (Atelerix albiventris) held as pets (Riley and Chomel 2005). In rodents, chronic C. parvum infections are facilitated by the presence of nematodes (Kulis-Malkowska 2007). Many coccidia, including Cryptosporidium spp. and Sarcocystis spp., are known to establish severe zoonotic infections in humans. Cryptosporidia are transmitted from small mammals to domestic animals or humans when their droppings contaminate feed stores (Meerburg et al. 2009) or surface and drinking water (Baldursson and Karanis 2011). From 2004 to 2010, 199 waterborne transmissions and outbreaks of parasitic protozoa occurred worldwide, with 60.3% of them involving Cryptosporidium spp. (Baldursson and Karanis 2011). Several outbreaks due to contaminated drinking water have been reported in recent years (MacKenzie et al. 1995, Exner and Gornik 2004, Brockmann et al. 2008). More than 1,000 different subspecies of Cryptosporidium spp. are described. A lack of consistency in the methods used (morphology of oocysts versus genetic differentiation) have made it difficult to distinguish animal-specific from zoonotic Cryptosporidium spp. to date.
Eggs of the nematode Capillaria spp. (syn. Calodium spp.) were detected in 5.4% of the small mammals, which may be either a contamination from the rodent's liver during dissection (Schmidt 2001) or the small mammals' ingested unembryonated eggs, leading to pseudoparasitosis ( = spurious infections) (Schmidt 2001, Fuehrer 2014). Herbivorous small mammals may be infected by the ingestion of embryonated eggs via contaminated plants (Schmidt 2001, Fuehrer 2014). The taxonomy of Capillaria spp. is also disputed and pending (Fuehrer 2014). Today there are more than 250 described Capillaria spp., with 4 species known to infect humans: Capillaria philippinensis, Capillaria hepatica (syn. Calodium hepaticum), Capillaria aerophilia (syn. Eucoleus aerophilus), and Capillaria plica (Cross 1992, McCarthy and Moore 2000). Human infections are acquired by ingesting larvae via fish, via contaminated vegetables, or by autoinfection. Though clinical symptoms are mild to absent in most cases, the number of zoonotic infections with Capillaria spp. is rising (McCarthy and Moore 2000), with the highest incidence in South East Asia due to consumption of traditional meals containing raw fish (Cross 1992, Saichua et al. 2008, Belizario et al. 2010). Hepatic capillariasis in Germany has only been described in travelers (Berger et al. 1990). The relevance of the Capillaria spp. detected in our study is not clear; molecular epidemiological studies are required.
One drawback of this study is that the differentiation between zoonotic and nonzoonotic species of the parasites was often hampered: In the case of the parasites with zoonotic potential, the lack of molecular species differentiation made it extremely difficult to distinguish the species, particularly due to the distortion and fragmentation of some of the developmental stages in our samples. New qualitative and quantitative PCR methods and whole genome sequencing analysis can help to distinguish between human-derived and animal-derived species, as shown for Hymenolepis spp. (Foronda et al. 2011), Capillaria spp. (McCarthy and Moore 2000), Giardia spp. (Thompson and Monis 2004, Helmy et al. 2018), and Cryptosporidium spp. (Di Giovanni and LeChevallier 2005). Another limitation lies in the pilot character of this study reflected in the broad spectrum of the small mammal species, number of individuals per species and their geographical origin.
Conclusions
This pilot study demonstrates that a broad spectrum of intestinal parasites is present in rodents and insectivores in Germany. The prevalence of zoonotic endoparasites, such as Hymenolepis spp. and Giardia spp., raises the important question about the epidemiological role of small mammals in the transmission of these pathogens to humans. Standardized PCR methods for zoonotic parasites, including tools for differentiating the strain or genotype of closely related species, are urgently needed for future studies. Genetic analyses would have the additional benefit of helping to reveal transmission routes from putative animal reservoirs to humans.
Footnotes
Acknowledgments
The authors thank Denny Maaz for rodent sampling at Welzow site and Hermann Ansorge, Thilo Liesenjohann, and Werner Gerwin for assistance in small mammal sampling. Dörte Kaufmann, Kati Sevke, Bärbel Hammerschmidt, Anna-Lena Schäfer, Eva Rühe, Denny Maaz, Thomas Büchner, Konrad Wanka, Jens-Peter Teifke, and Jens Lewitzki are acknowledged for their assistance during small mammal necropsy. The authors finally thank Denny Maaz, Christian Imholt, and Hortense Slevogt for a critical reading of the article and Martin H. Groschup for continuous support.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study within the network “Rodent-borne pathogens” was funded in part by the German Federal Ministry of Education and Research (BMBF) through the National Research Platform for Zoonoses (project codes 01KI1018 and 01KI1303) to R.G.U.
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.