
Abstract
More than 75 arboviruses (arthropod-borne viruses) have been identified in Australia. While Alfuy virus (ALFV), Barmah Forest virus (BFV), Edge Hill virus (EHV), Kokobera virus (KOKV), Murray Valley encephalitis virus (MVEV), Sindbis virus (SINV), Ross River virus (RRV), Stratford virus (STRV), and West Nile virus strain Kunjin (KUNV) have been associated with human infection, there remains a paucity of data regarding their respective transmission cycles and any potential nonhuman vertebrate hosts. It is likely that these viruses are maintained in zoonotic cycles involving native animals rather than solely by human-to-human transmission. A serosurvey (n = 100) was undertaken to determine the prevalence of neutralizing antibodies against a panel of Australian arboviruses in western gray kangaroos (Macropus fuliginosus) obtained from 11 locations in the midwest to southwest of Western Australia. Neutralizing antibodies against RRV were detected in 25%, against BFV in 14%, and antibodies to both viruses in 34% of serum samples. The prevalence of antibodies against these two viruses was the same in males and females, but higher in adult than in subadult kangaroos (p < 0.05). Twenty-one percent of samples had neutralizing antibodies against any one or more of the flaviviruses ALFV, EHV, KOKV, MVEV, and STRV. No neutralizing antibodies against SINV and KUNV were detected. If this sample of kangaroo sera was representative of the broader Australian population of macropods, it suggests that they are common hosts for RRV and BFV. The absence or low seroprevalence of antibodies against the remaining arboviruses suggests that they are not prevalent in the region or that kangaroos are not commonly infected with them. The detection of neutralizing antibodies to MVEV requires further investigation as this virus has not been identified previously so far south in Western Australia.
Introduction
Arboviruses (arthropod-borne viruses), particularly those transmitted by mosquitoes, present a significant risk to public health globally. More than 75 arboviruses have been identified in Australia, yet only a small number of them have been associated with human infection (Centers for Disease Control and Prevention [CDC] 2019). Ross River virus (RRV) (Fraser 1986), Barmah Forest virus (BFV) (Phillips et al. 1990), West Nile virus strain Kunjin (KUNV) (Muller et al. 1986), and the potentially fatal Murray Valley encephalitis virus (MVEV) (French 1952) are the most common human pathogens. Despite the severity of disease associated with infection and the notifiable status, there remains a paucity of information about the vertebrate hosts involved in the transmission of these viruses. Even less is known of the epidemiology of the remaining “neglected” Australian arboviruses that are associated with human infection, such as the alphavirus Sindbis (SINV) (Boughton et al. 1984) and the flaviviruses Alfuy (ALFV), Stratford (STRV) (Hawkes et al. 1985), Kokobera (KOKV) (Boughton et al. 1986), and Edge Hill (EHV) (Aaskov et al. 1993).
The recent emergence of chikungunya and Zika viruses highlights the potential for a pathogen considered relatively insignificant from a human health perspective to evolve into a major public health threat (Cao-Lormeau et al. 2016). The transmission cycles of many, if not all, of these “neglected” Australian arboviruses may involve a nonhuman vertebrate host, with human infection being only incidental (Russell and Kay 2004, Go et al. 2014). While serological surveys cannot demonstrate, conclusively, the ability of an animal to serve as a reservoir host, they do provide confirmation of infection. Evidence that marsupials, especially macropods (kangaroos, wallabies, wallaroos, and other members of this Australasian family of mammals), are reservoirs for—and focal hosts of—RRV has been derived from serological surveys, laboratory infections, and transmission of RRV to mosquitoes (Doherty et al. 1964, 1966, Kay et al. 1986, Kay and Aaskov 1989, Lindsay et al. 2005).
An extensive serosurvey undertaken by Potter et al. (2014) in the midwest to southwest regions of Western Australia found that 43.9% of blood samples collected from specimens of the western gray kangaroo (Macropus fuliginosus) had been infected with RRV. Since ALFV, BFV, EHV, KOKV, and STRV have been isolated from a number of similar mosquito vectors to those that transmit RRV such as Aedes vigilax, Aedes camptorhynchus, and Culex annulirostris (Poidingera et al. 2000, Johansen et al. 2005b, Macdonald et al. 2010) in Western Australia, the hypothesis for this study was that western gray kangaroos are infected with, and provide potential hosts for, a range of Australian arboviruses in addition to RRV. Sera from a random subpopulation of the samples examined by Potter et al. (2014) were assayed for the presence of neutralizing antibodies against a panel of well-recognized (RRV, BFV, MVE, and KUNV) and “neglected” (ALFV, EHV, KOKV, SINV, and STRV) Australian arboviruses associated with human infection.
Materials and Methods
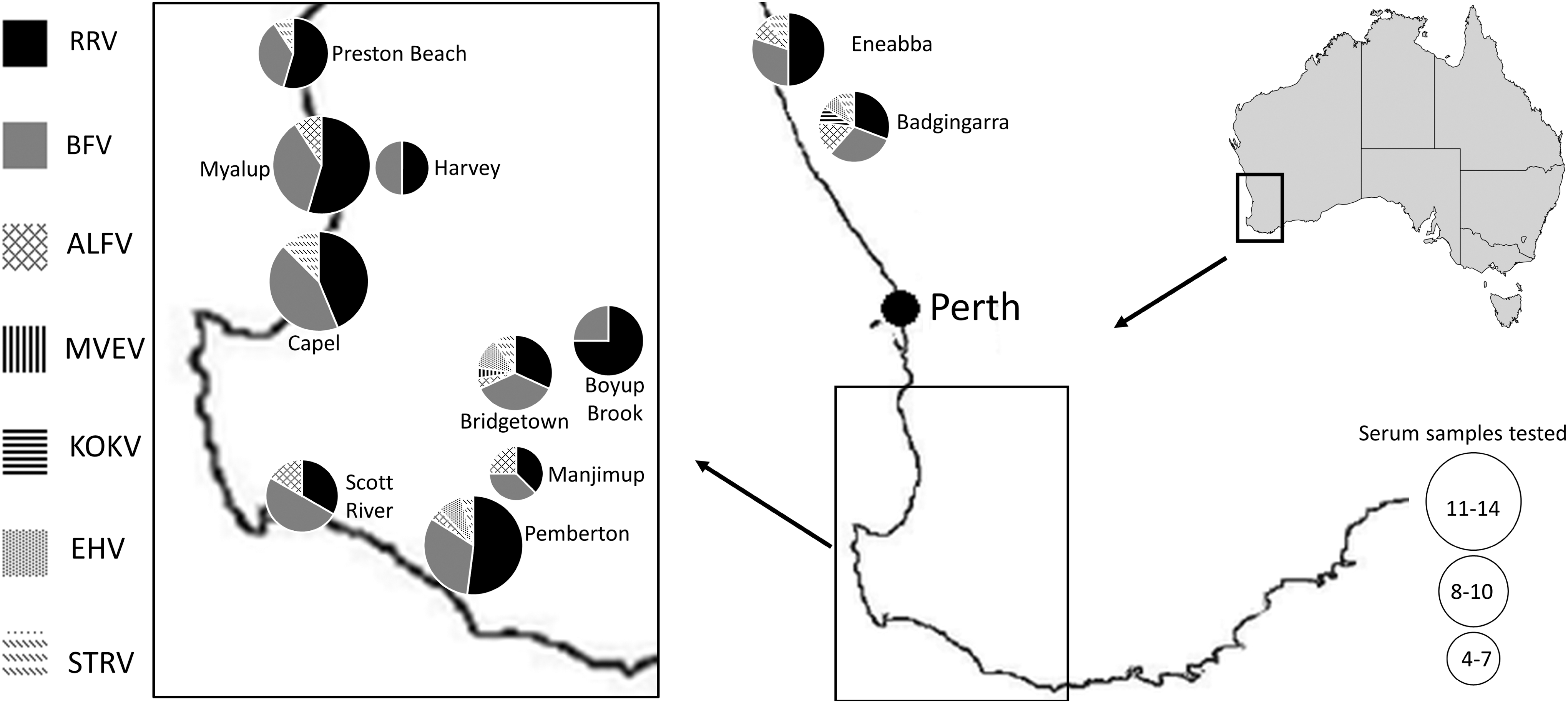
A random subset of 100 sera from those collected by Potter et al. (2014) from 11 different locations throughout the midwest and southwest of Western Australia (Fig. 1) was analyzed. Peripheral blood samples were collected between May 2006 and May 2009 from commercially harvested, wild-caught kangaroos. Persons harvesting kangaroos commercially are licensed and must adhere to a code of practice to ensure the humane culling of animals (Australian Government Department of the Environment and Energy 2019). The Murdoch University Animal Ethics Committee that oversaw the Potter et al. (2014) study requested that it be informed of sample collection, but did not require specific ethics approval. Information on the location of each kangaroo from which serum was obtained, the animal's gender, and approximate age was provided by the harvester responsible. The ages of the kangaroos were categorized as pouch young (still in pouch), subadult (<3 years of age), or adult (>3 years of age) on the basis of size and apparent sexual maturity. Sera were transported from Murdoch University in Perth to the World Health Organization Collaborating Centre for Arbovirus Reference and Research at the Institute of Health and Biomedical Innovation, Queensland University of Technology in Brisbane for analysis.
Using the value of 43.9% for the seroprevalence of anti-RRV antibodies (Potter et al. 2014), and a margin of error of 0.1, a minimum sample size of 96 sera was required to determine the prevalence of antibodies against Australian arboviruses with 95% confidence. One hundred serum samples were analyzed in this study and the following prototype strains of viruses were used: ALFV MRM3929 (Whitehead et al. 1968); BFV BH2193 (Marshall et al. 1982); EHV C281, KOKV CH112820, KUNV MRM16, RRV T48, SINV MRM 39, and STRV C338 (Doherty et al. 1963); and MVEV MVE/1/1951 (French 1952). In Tables 1 and 3, the seroprevalence for each virus was calculated by summing all samples that neutralized the virus by ≥50%.
Neutralization of Ross River Virus, Barmah Forest Virus, or Both Viruses by Sera from Western Gray Kangaroos (Macropus Fuliginosus, n = 100)

The neutralization value is significantly greater (p < 0.05, Student's t-test) than the corresponding neutralization value for other viruses in the same row. Gray shades, both dark and light, represent reactive neutralization values.
Total reactive numbers in bracket are based on an assumption that each kangaroo had been infected with a single virus and it was the one against which the highest neutralizing activity (darker gray shade) was observed.
A, adult; BFV, Barmah Forest virus; SA, subadult; PY, pouch young; RRV, Ross River virus.
Plaque reduction neutralization test
Although most Australian alphaviruses and flaviviruses have been reported to form plaques on monolayers of porcine stable-equine kidney (PS-EK) cells (Gorman et al. 1975), BFV did not do so in this study. As all alphaviruses of interest, including BFV, formed plaques on baby hamster kidney (BHK-21) cells, this cell line was used for the plaque reduction neutralization test (PRNT) with alphaviruses. The flavivirus PRNTs were performed on PS-EK cells.
Equal volumes (500 μL) of sera inactivated by heating at 56°C for 30 min and diluted 1 in 10 in serum-free medium (Roswell Park Memorial Institute 1640 [RPMI-1640]; Sigma-Aldrich, MO) and virus (stock virus diluted to produce between 50 and 60 plaques when added to cell monolayers) were mixed and incubated at 37°C for 60 min. The culture medium from confluent cell monolayers in 12-well tissue culture plates (Nunclon; Thermo Scientific, Victoria, Australia) was removed and 400 μL of the virus/serum mixtures transferred into duplicate wells and incubated at 37°C in an atmosphere of 5% CO2/air for 120 min to enable the non-neutralized virus to adsorb to cells. Following incubation, 2 mL of 0.75% w/v carboxymethyl cellulose (CMC; Sigma-Aldrich)/RPMI 1640 was added. The plates were incubated at 37°C in an atmosphere of 5% v/v CO2/air for 2 days for BFV, RRV, and SINV or 4 days for ALFV, EHV, KOKV, KUNV, MVEV, and STRV. The CMC overlay was removed and cells were fixed and stained with 0.05% w/v crystal violet (Sigma-Aldrich) in formaldehyde (1% v/v) and methanol (1% v/v). The crystal violet solution was recovered from the plates and discarded, the cell monolayers rinsed in tap water, and the plates inverted on a paper towel and allowed to dry before plaques (clear zones in a purple cell monolayer) were counted. Sera that reduced plaque numbers by ≥50% were scored as reactive. The significance of differences between neutralization by different sera or between neutralization of different viruses was calculated using Student's t-test.
Results
Antibodies against 7 different Australian arboviruses were detected in the 100 serum samples collected from western gray kangaroos at 11 locations in Western Australia (Fig. 1). Antibodies in 25 samples neutralized only RRV, antibodies in a further 14 reacted only with BFV, and no neutralizing antibodies against SINV were detected. Antibodies in an additional 34 sera reacted to both RRV and BFV (Table 1).
The seroprevalence of anti-RRV and anti-BFV antibodies among adult kangaroos was significantly higher than in subadults (p < 0.05, Table 2). There was no significant difference (p > 0.05) between males and females in the seroprevalence of either anti-RRV (60.6%: 56.4%) or anti-BFV antibodies (45.5%: 48.7%).
Association Between Animal Age and the Prevalence of Neutralizing Antibodies Against Ross River Virus and Barmah Forest Virus in Western Gray Kangaroos (Macropus Fuliginosus, n Tested = 100)
The term preadult combines both subadult and pouch young age groups.
Neutralization of Flaviviruses by Sera from Western Gray Kangaroos (Macropus Fuliginosus, n = 100)
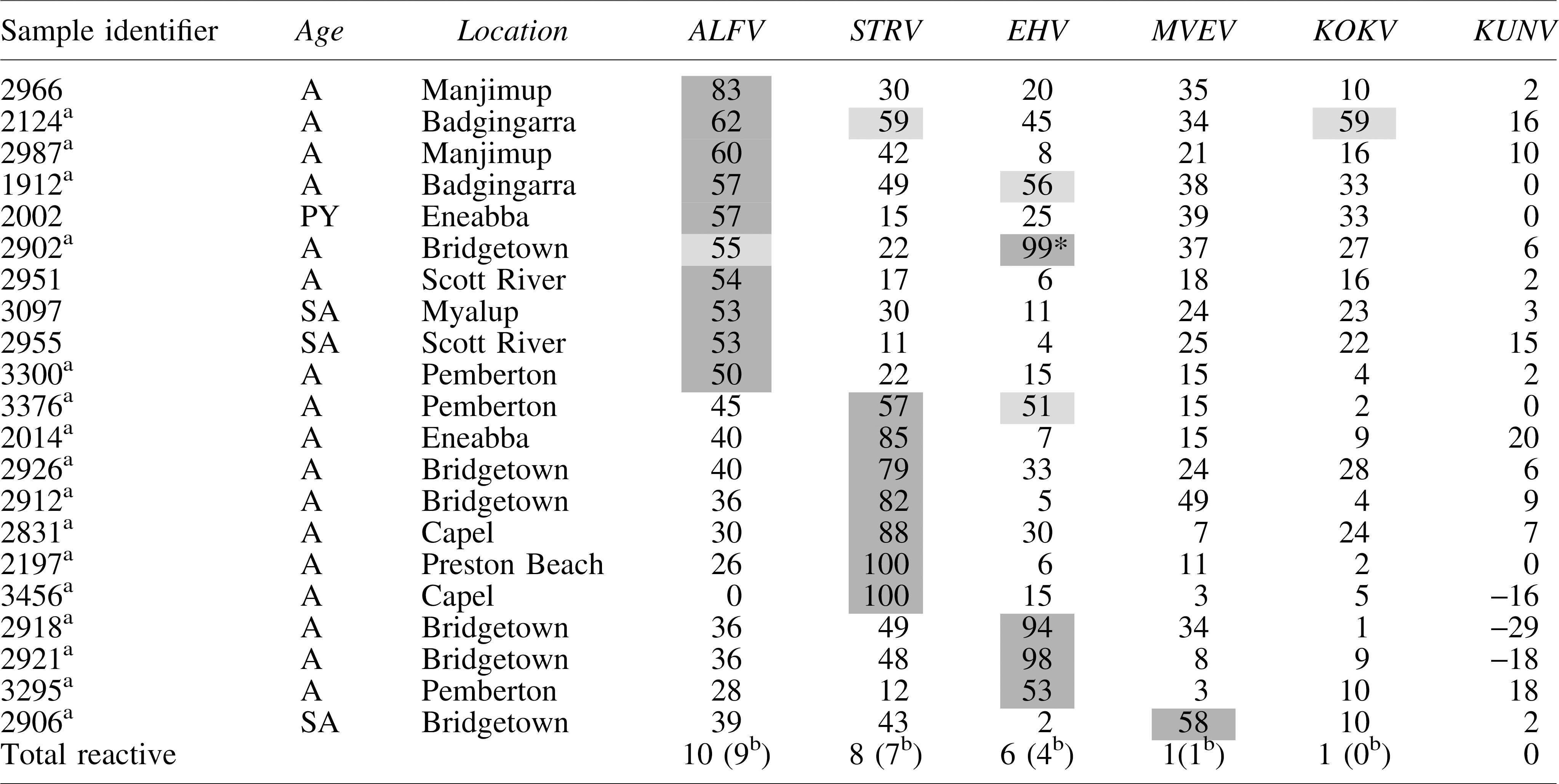
The neutralization value is significantly greater (p < 0.05, Student's t-test) than the corresponding neutralization value for other viruses in the same row.
Samples of those neutralized in both alphavirus and flavivirus. Gray shades, both dark and light, represent reactive neutralization values.
Numbers in brackets are based on an assumption that each kangaroo had been infected with a single virus and it was the one against which the highest neutralizing activity (darker gray shade) was observed.
ALFV, Alfuy virus; EHV, Edge Hill virus; KOKV, Kokobera virus; KUNV, Kunjin virus; MVEV, Murray Valley encephalitis virus; STRV, Stratford virus.
Twenty-one percent of kangaroo sera contained neutralizing antibodies against the flaviviruses used—ALFV (10%), STRV (8%), EHV (6%), KOKV (1%), and MVEV (1%). Four of the 21 sera that neutralized flaviviruses reacted with more than one virus (Table 3). No sera neutralized KUNV, but three (identified samples 3456, 2918, and 2921) consistently enhanced the number of plaques formed by this virus (Table 3). All three of these sera neutralized other flaviviruses (3456, STRV; 2918, EHV; 2921, EHV). Sixteen of the 21 samples that reacted with flaviviruses also reacted with alphaviruses (Table 3). The numbers of flavivirus-reactive sera were too small to draw any conclusions about gender or age relatedness.
Discussion
The present study used a neutralization test in an effort to better discriminate between infections with distinct, but related, arboviruses. This type of assay is more labor-intensive and expensive than a number of the other serological tests available and so constrained the study to the smallest sample able to provide significant data (see Materials and Methods). Nonetheless, the anti-RRV antibody prevalence value obtained with the neutralization test on a subsample of this panel of sera (59%) was similar to that obtained by Potter et al. (2014) on the entire collection of 2632 kangaroo sera, thus suggesting that there was no major bias in the subsampling.
The prevalence of anti-alphavirus antibodies (73%) was higher than that for anti-flavivirus antibodies (21%), despite performing assays for antibodies against twice as many flaviviruses as alphaviruses. The study was not able to determine whether this difference, and the difference in the prevalence of antibodies against the various members of the alphavirus and flavivirus groups, was due to the availability of the viruses to cause infection or to the susceptibility of the kangaroos to infection with each virus. Widespread infection of macropods throughout Australia with RRV has been reported previously—89% in eastern gray kangaroos (Macropus giganteus) at Mitchell River Mission in north Queensland (Doherty et al. 1971), 40% in eastern gray kangaroos in Tasmania (McManus and Marshall 1986), and 50% in eastern gray kangaroos in New South Wales (Vale et al. 1991), and the results obtained here are consistent with these earlier studies. In addition, RRV was recovered from wallabies in north Queensland (Doherty et al. 1971), while Kay et al. (1986) detected viremia for 4–6 days after experimental infection of eastern gray kangaroos, high enough [up to 105 suckling mouse intra-cerebral inoculation, median lethal dose (SMIC LD50/mL)] to infect a number of common mosquito vectors of RRV. All these observations point to macropods having a potential role in the transmission cycle of RRV.
Antibodies against BFV in kangaroos have been reported only once previously (Vale et al. 1991), and then, in only 1 of 25 eastern gray kangaroos sampled. Fourteen percent of the animals sampled in the study reported here showed a monotypic anti-BFV antibody response, suggesting that they had been infected with this virus. It is also likely that many of the sera that contained both anti-RRV and anti-BFV antibodies represented animals that had been infected with BFV. These results are in contrast to those of Johansen et al. (2005a) who did not detect anti-BFV antibodies in 128 kangaroo sera collected in 1992 and 1993 in similar localities to those from which the animals were sampled in the present study. BFV was recovered for the first time in Western Australia, from mosquitoes, in the east Kimberly region in 1989 (Broom et al. 1993), and the first clinical cases were recognized in the southwest of Western Australia in 1992 and 1993. This was almost two decades after the first isolation of BFV from mosquitoes in the eastern states of Australia in 1974 (Marshall et al. 1982). The simplest explanation for the proportion of kangaroos with anti-BFV antibodies observed herein is that BFV was introduced into Western Australia around 1989, since when it has become established in rural/urban cycles similar to those that occur for RRV transmission (Gyawali et al. 2017). This is supported by the reports of increasing numbers of cases of BFV infection in humans in Western Australia in the last two decades (Australian Government Department of Health 2019).
Given the similarities in the epidemiology of BFV and RRV, including their mosquito vectors, it was not surprising that 34% of animals appeared to have been infected with both viruses. Although serological cross-reactivity between alphaviruses is reported to be low (Schmaljohn and McClain 1996), it is not possible to discount the likelihood that some of the reactions to RRV or to BFV were due to cross-reactions following infection with only one of these viruses.
The significantly higher seroprevalence in adult compared with subadult kangaroos, for both RRV and BFV, remains consistent with Potter et al. (2014). An increased possibility as the host ages of being infected on multiple occasions by these arboviruses might have resulted in a higher chance of seroconversion. The detection of anti-BFV antibody in the pouch young of a kangaroo from Eneabba (sample 2375 in Table 1) is a significant finding. This animal was less likely to have been bitten by a mosquito while in the pouch than to have acquired the antibodies from its mother. Potter et al. (2014) provided compelling evidence for the transmission of anti-RRV antibodies from kangaroo does to their young.
The absence of antibodies against SINV was considered surprising but, arguably, is not unexpected since the preferred hosts for SINV are avian species (Whitehead et al. 1968, Doherty 1972). The detection of antibodies to SINV in western quolls, emus, rabbits, and horses in the same localities in Western Australia from which the samples for this study were obtained (Johansen et al. 2005a) suggests that macropods are not a preferred host for mosquitoes infected with SINV in this region of Australia.
The prevalence of anti-flavivirus antibodies in the kangaroo sera sampled was significantly lower than that observed for anti-RRV and anti-BFV antibodies (Tables 1 and 3), and lower than those against ALFV, EHV, KOKV, and KUNV observed by Doherty et al. (1971) in kangaroos and wallabies in northeastern Australia. This may reflect the difference in ecological niches between the Doherty et al. (1971) study (tropical northern Australia) and this one (cool, Mediterranean climate). EHV, KOKV, and STRV have been isolated from A. vigilax and A. camptorhynchus in the regions from which these kangaroo sera were obtained (Poidingera et al. 2000, Johansen et al. 2005b, Macdonald et al. 2010). However, there is no experimental evidence of replication of these viruses in macropods so it is unclear whether the low prevalence of antibodies against these viruses in the present study was because the animals had not been infected at all or because they had been infected, but virus replication was insufficient to elicit a serological response.
The KUNV has not been recovered from the southwest region of Western Australia, and so, the absence of antibodies to this virus in the present sample of kangaroo sera might have been predicted. What was surprising, however, was the enhancement of KUNV infection with antibodies against STRV (one example) and against EHV (two examples) (Table 3). There is an extensive literature describing enhancement of flavivirus infection in vitro by cross-reactive and non-neutralizing antibodies, beginning with that of Hawkes (1964) who also described enhancement of infection by WNV. It is not known whether or not the PS-EK cells used in this study express the Fc receptors required for antibody-dependent enhancement of viral infection to occur (Kliks and Halstead 1983) or if another mechanism was responsible for our observations.
Doherty et al. (1971) detected antibodies against MVEV in 85% (24/28) of kangaroos sampled from northern Queensland, while a kangaroo infected by the bite of an MVEV-infected mosquito developed a viremia of 4.6 ± 2 SMIC LD50/mL (Kay et al. 1986). However, MVEV is not known to be active in the region from which the kangaroo sera in this study were sourced (Broom et al. 2001a, 2001b). The only serum sample (2906, Table 3) that was reactive to MVEV neutralized ALFV and STRV to similar levels, thereby making it impossible to draw any conclusion about whether or not an MVEV infection had occurred. Extensive cross-reactivity between flaviviruses has been reported previously when using polyclonal antisera for testing (De Madrid and Porterfield 1974, Calisher et al. 1989, Hall et al. 1990).
The presence of significant numbers of sera containing anti-ALFV antibodies was not anticipated, but this virus has been isolated from different places in the Kimberly (northern) region of Western Australia (May et al. 2006) and these data may indicate a southward movement of the virus.
Conclusions
This study suggests that western gray kangaroos are likely to be significant hosts in the transmission cycle of RRV in Western Australia. Moreover, if these animals develop BFV viremia similar to those in kangaroos infected experimentally with RRV, then they are probably significant hosts for BFV as well. The high prevalence of antibodies against RRV and BFV suggests that mosquitoes able to transmit the Australian flaviviruses ALFV, EHV, KOKV, KUNV, MVEV, and STRV, as well as the alphavirus SINV, feed on western gray kangaroos. The absence of antibodies to these viruses implies that this native macropod species, the most common kangaroo species in Western Australia, is unlikely to play a major role in cycles of transmission involving these “neglected” arboviruses, at least in the settings studied.
Authors' Contributions
N.G., A.W.T.-R., R.S.B., and J.G.A. conceived the project. N.G. carried out laboratory experiments and drafted the article. A.P. collected the samples. J.G.A. supervised the project. N.G., A.W.T.-R., R.S.B., A.P., and J.G.A. critically reviewed and revised various versions of the article. All authors contributed to the preparation of the final version and agreed to its submission.
Footnotes
Authors' Disclosure Statement
The authors declare that no competing financial interests exist. R.S.B. is coauthoring this article in his personal capacity and his capacity as an adjunct academic at Central Queensland University.
Funding Information
N.G. was in receipt of an International Postgraduate Research Scholarship and an Australian Postgraduate Award administered by Central Queensland University. This study was supported, in part, through a Health Collaborative Research Network Merit Grant and funds provided from the Cook Estate.