
Abstract
Zika virus (ZIKV) is an emerging mosquito-borne pathogen belonging to the genus Flavivirus of the family Flaviviridae. Aedes albopictus is widely distributed in China. However, little is known about the vector competence of Ae. albopictus in China. The present study presents the oral susceptibility and vector competence of Ae. albopictus Guangzhou strain to ZIKV. Additionally, vertical transmission of ZIKV is described. The results demonstrated the susceptibility of local Ae. albopictus mosquitoes to ZIKV with an extrinsic incubation period of 6 days. Disseminated infection was observed in Ae. albopictus starting on day 2 postinfection (PI). Starting on day 6 PI, the saliva of Ae. albopictus exhibited ZIKV infection, and the transmission rate was 36.4%. Vertical transmission was observed during the first gonotrophic cycle. The minimum infection rate was observed in third-to-fourth instar larvae.
Introduction
Zika virus (ZIKV) is an emerging mosquito-borne virus belonging to the genus Flavivirus of the family Flaviviridae (Kuno et al. 1998). It is a positive single-stranded RNA virus with a 10,794-nucleotide genome that is closely related to the Spondweni virus (Kuno and Chang 2007, Hayes 2009). ZIKV was initially isolated in Uganda in 1947 (Dick et al. 1952), and its tendency to spread outside its known geographic range as well as its ability to cause large-scale epidemics together constitute a major threat to global health in both developing and developed countries. Imported cases, as well as the first ZIKV strain isolate, have been reported in China (Deng et al. 2016). The primary vectors of ZIKV are Aedes spp. mosquitoes; specifically, the virus has been isolated mainly from Aedes africanus (Haddow et al. 1964) and Aedes apicoargenteus (McCrae et al. 1982) in Uganda and the Central African Republic; from Aedes luteocephalus in Nigeria in 1969 and 1972 (Fagbami 1979); from Aedes vittatus and Aedes furcifer in Ivory Coast in 1999 (Akoua-Koffi et al. 2001), and from Aedes aegypti in Malaysia (Marchette et al. 1969). Thus, ZIKV has potentially emerged in many countries where Aedes spp. mosquitoes are present.
Aedes albopictus Skuse is a main vector or potential vector of many human and veterinary pathogens. Viral isolation and vector competence studies have established the efficiency of this mosquito in the transmission of more than 20 arboviruses (Paupy et al. 2009). In mainland China, Ae. albopictus is the primary vector for dengue virus and chikungunya. This mosquito is widely distributed, with a range extending from the southern reaches to both the northern and western parts of China, with northern fringes in Shenyang in Liaoning Province, through Tianshui and Longnan in Gansu Province, to Motuo in Tibet in China (Lu 1990). Ae. albopictus is also an efficient laboratory vector of ZIKV (Wong et al. 2013), also the virus has been isolated from field-collected mosquitoes (Wong et al. 1993, Grard et al. 2014). Moreover, some experimental studies have described the transmission and dissemination of ZIKV by Ae. albopictus in the Americas (Chouin-Carneiro et al. 2016) and in Italy (Di Luca et al. 2016) and so on. However, little is known about the vector competence of Ae. albopictus in China.
The identification of potential vectors of either ZIKV or any other mosquito-borne diseases in a given geographical area has important implications for the control of disease outbreaks. Due to the known epidemic vector role and widespread distribution of Ae. albopictus in China, along with its opportunistic feeding behavior, ecological adaptability, and propensity, it is important to determine the competence of Ae. albopictus to transmit ZIKV. The present study presents the oral susceptibility and vector competence of Ae. albopictus Guangzhou strain to ZIKV. Additionally, vertical transmission of ZIKV is described.
Materials and Methods
Mosquito strain
Ae. albopictus mosquitoes (Guangzhou strain) were originally collected in Guangzhou city (GPS location: 23°08′N and 113°14′W), Guangdong Province. Mosquitoes were reared at 25°C ± 1°C at 75% ± 5% relative humidity (RH) under a 14-h light/10-h dark (LD) photoperiod. The F9 generation of Ae. albopictus mosquitoes were used in this study. Adult mosquitoes were regularly fed on a 10% sugar solution.
Virus strain and cell line
The SZ01 ZIKV strain (GenBank No. KU866423) was obtained from the Microbial Culture Collection Center of the Beijing Institute of Microbiology and Epidemiology. This virus was originally isolated from the blood of a patient who had returned from Samoa to China in 2016 (Deng et al. 2016). The SZ01 strain was propagated in C6/36 cells and passaged three times.
Ae. albopictus C6/36 cells were grown in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% and 100 U/mL each of penicillin and streptomycin (Gibco). Infected cells were incubated for 3–5 days in DMEM (GIBCOTM; Invitrogen, Beijing, China) with 2% fetal bovine serum (FBS) and 1% penicillin and streptomycin. Viruses were harvested and stored as individual 1-mL aliquots in freezing tubes at −80°C. Viral titer was expressed in plaque-forming units (PFUs).
Oral infection in mosquitoes
Seven-day-old female mosquitoes were deprived of sucrose solution for 12 h before being offered an infectious blood meal. The infectious blood meal consisted of 1:1 mouse blood and virus suspension. The viral titer of infectious blood meals was 3 × 105 pfu/mL. Mosquitoes were allowed to feed for 1.5 h using a Hemotek membrane feeding system housed in a feeding chamber that was constantly warmed to 37°C. Fully engorged females were transferred to 300-mL plastic cups and maintained at 29°C ± 1°C and 75% ± 5% RH with a 14-h light/10-h dark cycle photoperiod. Three days after ingesting the infectious meal, mosquitoes were blood fed to enable egg production. Concurrently, uninfected control mosquitoes were blood fed and eggs collected.
ZIKV detection in the midgut, salivary glands, and ovaries of oral-fed mosquitoes by real-time RT-PCR
To evaluate ZIKV replication in the midgut (MG), salivary glands, and ovaries of individual Ae. albopictus mosquitoes, 25 female mosquitoes were sampled on days 2, 4, 6, 8, 12, 16, and 20 postinfection (PI). To prevent crosscontamination of virus between the MG, salivary glands, and ovaries of each mosquito, organs were carefully dissected using different dissecting needles and then iteratively rinsed in phosphate-buffered saline (PBS). All dissecting needles were dipped in 80% ethanol and cleaned before reuse. When dissecting organs, all dissecting needles were soaked in 80% ethanol, then burnt on alcohol lamp, followed by dipping into sterile PBS until the needles cooled. The MG, salivary glands, and ovaries from each mosquito were individually transferred into 1.5-mL microtubes containing 100 mL of DMEM (GIBCO™; Invitrogen, Beijing, China) supplemented with 2% FBS. Organs were then homogenized using 5 mm stainless steel grinding balls with a Bullet Blender™ 24 mixer mill that was set to a frequency of 12 rotations/s for 1 min. The mice (Kunming mice) were provided by the Experimental Animal Center at the Military Medical Science Academy, China.
Total RNA was isolated from the MGs, salivary glands, and ovaries of mosquitoes using the QIAampViral RNA Mini Kit (Qiagen, Germany) according to the manufacturer's recommendations. ZIKV was detected in these organs using a Detection Kit for ZIKV RNA (Daan Gene, China) that included a reverse primer 5′-TACAAGTACCATCCTGACTCCC-3′, a forward primer 5′-CYGTCAGTTGYACTCCATTCTC-3′, and a locked nucleic acid probe (5′-FAM-AGCTCCCCTTCYACTGATYTCCACAT-BHQ1-3′). The following reagents were used for PCR reactions: 5 μL of each RNA sample, 17 μL of PCR Mixture A, and 3 μL of PCR Mixture B to yield a 25 μL final reaction volume. Amplification reactions were performed in a real-time PCR system from Applied Biosystems (ABI 7500) programmed as follows: 1 cycle at 50°C for 15 min, 95°C for 15 min, 40 cycles at 94°C for 15 s, and 55°C for 45 s.
Viral titration
ZIKV supernatants were serially diluted in the same diluent and inoculated at 0.1 mL volumes into baby hamster Syrian kidney (BHK) cell culture under agar overlay. Cells were incubated for 7 days at 37°C and were then stained with a Crystal Violet solution (0.2% in 10% formaldehyde and 20% ethanol). The presence of viral particles was assessed by detecting cytopathic effect. Titers were expressed in pfu.
Assessment of vector competence
Saliva was collected from individual mosquitoes on days 4, 6, 8, 10, 12, 14, 16, 18, and 20 PI as previously described by Dubrulle et al. (2009). Briefly, the wings and legs of each mosquito were removed, and the proboscis was inserted into a quartz capillary containing 5 μL of FBS. After 30 min, FBS-containing saliva was expelled in 100 μL of DMEM to detect ZIKV using real-time RT-PCR as previously described.
On day 8 PI, 30 mosquitoes were fed on four 2-day suckling mice to investigate horizontal transmission capacity. Suckling mice were observed for sickness, and brains were harvested on day 10 and assessed for the presence of virus by performing RT-PCR and plaque assays. Blood-engorged mosquitoes were removed from each cage under CO2 anesthesia. The salivary gland of each blood-engorged mosquito was dissected and analyzed using real-time RT-PCR.
Assessment of vertical transmission
Eggs obtained from the first gonotrophic cycle were used in this experiment. Fully fed mosquitoes were allowed to oviposit, and egg rafts from individual mosquitoes were maintained under a humid environment for 48 h, then were briefly air dried. Eggs were maintained in desiccators at 25°C ± 1°C and 90% RH with a 14-h light/10-h dark daily photoperiod for 6 days to permit embryonation (Hanson and Craiq 1994). After eggs were laid, parental females were tested for the presence of virus by RT-PCR.
A total of 30 eggs represented a single pool for virus detection. Following embryonation, Third-to-fourth instar larvae were pooled at a rate of 5 per pool; male adult mosquitoes (3–4 days after emergence) were pooled at a rate of 5 per pool. Individual pools of adult mosquitoes (♂)/eggs/larvae were homogenized and assessed for the presence of virus by performing real-time RT-PCR, as described above. The salivary glands of each female mosquito were dissected and analyzed. The minimum infection rate (MIR) of adult mosquitoes (♂)/eggs/larvae was calculated as the ratio of the number of positive pools to the total number of tested mosquitoes.
Data analysis
Mosquito MGs were analyzed to evaluate the infection rate (IR), which was calculated as the number of ZIKV-positive MGs compared with the total number of fed females. Salivary glands and ovaries were tested to assess the dissemination rate (DR), which was calculated as the number of specimens with ZIKV-positive salivary glands plus ovaries among the number of ZIKV-positive MGs. The saliva of infected females was processed to assess the transmission rate (TR), which corresponded to the number of mosquitoes with infectious saliva/mosquitoes with salivary gland infection. p-Values of <0.05 were considered significant. A χ 2 test for linear trend analysis was used to assess the IR.
Results
Oral susceptibility of Ae. albopictus to ZIKV
The ZIKV IRs of individual mosquito MGs, salivary glands, and ovaries were measured in Ae. albopictus female mosquitoes. Beginning on day 2 PI, ZIKV was detected in mosquito MGs, salivary glands, and ovaries on each sampling day. There was a significant increase in the IR (p = 0.011, χ 2 = 15.767) (Table 1). Disseminated infection was observed in Ae. albopictus starting on day 2 PI. By day 12 PI, the DR was 100% (Table 1). Beginning on day 6 PI, Ae. albopictus saliva tests revealed ZIKV infection with a TR of 44.4% (Table 1).
Infection, Dissemination, and Transmission Rates for Aedes albopictus Orally Fed with Zika Virus at Various Days Postinfection
DR, dissemination rate; IR, infection rate; MG, midgut; SG, salivary gland; TR, transmission rate.
ZIKV growth kinetics in the MGs, salivary glands, and ovaries of mosquitoes
Virus RNA copy numbers were calculated by performing real-time RT-PCR and were expressed as the equivalent log10 RNA copy/mL. Standard RNA used for nucleic amplification assays was extracted from virus dilutions containing known titers (Supplementary Table S1), which had previously been determined using a plaque assay. Infectious virus particles represent a small portion of the total number of RNA transcripts in a sample, including defective interfering particles and unpackaged viral RNA.
ZIKV infection growth curves in the MG, salivary glands, and ovaries are shown in Fig. 1. RNA copy numbers and growth rates for the MG were higher than those of the salivary glands and ovaries. Starting on day 4 PI, the titers and growth rates in the salivary glands and ovaries were similar. Peak titers in the salivary glands and ovaries were 3.5 and 4.0 log10 RNA/mL, respectively, in 20 days.
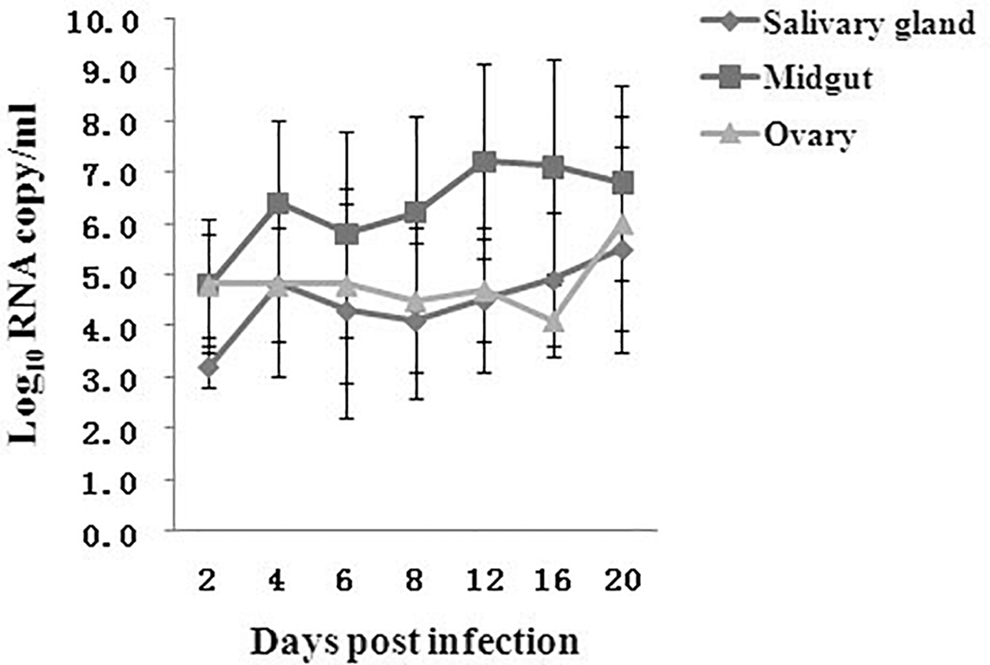
Virus growth in the midguts, salivary gland, and ovaries of Aedes. albopictus mosquitoes. The virus copy shown represents the mean titer ± standard deviation (log10 RNA copies/mosquito) from three mosquitoes collected at each time point.
Presence of virus in saliva
ZIKV was detected in mosquito saliva, indicating viral transmission by mosquitoes. Although ZIKV was detected in 3 of 20 mosquito salivary glands on day 4 PI, virus was not detected in the saliva (Table 2). ZIKV was detected in saliva starting on day 6 PI onward in Ae. albopictus mosquitoes. RNA was present on day 6 PI at 3.2 copies/mL. Maximum RNA replication was observed on day 12 PI at 5.1 log10 RNA copies/mL.
Zika Virus Infection Rates in Aedes albopictus Saliva on Various Days Postinfection
A total of 19–25 mosquitoes was sampled each day.
TR = number of mosquitoes with infectious saliva/mosquitoes with salivary gland infection.
TE = number of mosquitoes with infectious saliva/total number of mosquitoes tested.
PI, postinfection; TE, transmission efficiency.
Horizontal transmission of ZIKV in Ae. albopictus mosquitoes
On day 8 PI, 30 mosquitoes were fed on four 2-day-old suckling mice. A total of 14 blood-engorged mosquitoes were collected and 4 were subsequently detected as virus-positive in salivary glands. At day 10 postsuckling, all brains of mice were detected as ZIKV positive, with titers ranging from 1 × 104 to 2 × 105 pfu/mL.
Vertical transmission of ZIKV in Ae. albopictus mosquitoes
Oviposition commenced from day 3 PI and continued until day 6. By real-time RT-PCR, ZIKV could be detected in eggs laid by infected mosquitoes or in immature (larvae) and adult female mosquitoes (Table 3). The MIR per egg was ∼4.4/1000 for 450 eggs tested in 15 pools. Pools of third-to-fourth instar larvae were processed to confirm the presence of virus, which revealed that the MIR was ∼53.3/1000 for 75 larvae tested in 15 pools. Virus could not be detected in 13 pools comprising 65 male mosquitoes. Only a single salivary gland of adult females tested positive for ZIKV and the RNA copy number was 4.1 log10 RNA copies/mL.
Minimum Infection Rate for Zika Virus Positive in F1 Generations of Aedes albopictus Mosquitoes
Discussion
ZIKV is an emerging mosquito-borne pathogen that represents a major threat to global health in both developing and developed countries. A large-scale outbreak in the Americas and Asia has shown the propensity of ZIKV to spread outside of its known geographical range and its potential to cause large-scale epidemics (Haddow et al. 2012). This virus has the potential to spread globally through the carriage of goods and travelers (Bogoch et al. 2016). Currently, ∼14 cases of ZIKV infection have been reported in mainland China. The absence of vaccines and specific treatments for ZIKV means that prevention and control rely heavily upon vector control. To date, little is known about the vectors of ZIKV in China.
In the present study, the susceptibility, replication kinetics, transmission potential, and vertical transmission of Ae. albopictus mosquitoes from China were evaluated for the first time using the SZ01 strain of ZIKV. Based on our work, Ae. albopictus mosquitoes are susceptible to ZIKV and demonstrate increasing infection and DRs. Growth patterns and viral RNA copy numbers were nearly identical in the salivary glands and ovaries (Fig. 1). However, RNA copy numbers and growth rates in the MG were higher than those in the salivary gland and ovaries. By day 6 PI, 33.3% (n = 4/12) of mosquitoes exhibited infected salivary glands, and among these, four (20%) had detectable ZIKV RNA in the saliva. By day 8 PI, 63.6% of infected mosquitoes were capable of transmitting the virus. Thus, Ae. albopictus are susceptible to ZIKV, with an extrinsic incubation period (EIP) of 6 days. The EIP corresponds to the time required for the virus to reach mosquito saliva after ingesting an infectious blood meal (Kramer and Ebel 2003). In a previous study, Ae. albopictus mosquitoes from Singapore were competent to harbor ZIKV with an EIP of 4 days, and all infected mosquitoes were capable of transmitting virus on day 7 PI (Wong et al. 2013). A longer EIP was observed in Ae. aegypti infected with ZIKV from Italy (Di Luca et al. 2016). The differences between the EIP and TRs observed in this study and those reported for other studies are attributable to genetic variability between Ae. albopictus populations from different geographical origins. The impact of this variability on vector competence has been demonstrated for several vector/virus associations (Tesh et al. 1976, Gubler et al. 1979). Lambrechts et al. (2009) demonstrated that the vector competence is determined by the specific combination of vector and virus genotypes, which also contributed to the virus adaptation to local vectors.
Additionally, ZIKV-infected Ae. albopictus mosquitoes were maintained under insectary conditions at 29°C ± 1°C and 80% ± 5% RH above 20 days and the survival rate was about 90% in our laboratory. Ae. albopictus in China appear to be highly efficient vectors of ZIKV, which may be balanced by the large number of susceptible humans and their close contacts with Aedes mosquitoes.
Horizontal transmission of ZIKV by different vector mosquitoes has been reported in several studies. In 1956, Boorman and Porterfield successfully transmitted virus to both mice and monkeys using ZIKV-infected laboratory strains of Ae. aegypti. Cornet et al. (1979) further observed ZIKV transmission by a high percentage (88%) of intrathoracically infected Ae. aegypti to experimental mice within 7 days. Salivary gland infection and the presence of virus in saliva are prerequisites for the designation of a mosquito as a vector. In the present study, virus was first detected in saliva on day 6 PI. Moreover, successful transmission to infant mice through biting was also demonstrated after 10 days of incubation. Infant mice exhibited virus titers ranging from 1 × 104 to 2 × 105 pfu/mL of ZIKV in the brain on day 10.
Vertical transmission of ZIKV has been observed during field mosquito surveillance (Diallo et al. 2014). The detection of ZIKV in a pool of male Ae. furcifer mosquitoes strongly suggests vertical transmission in this species; this transmission may represent an important mechanism for local maintenance (Diallo et al. 2003, 2012, Mavale et al. 2010). Although Ae. albopictus has long been suspected to be a vector of ZIKV (Olson et al. 1981), previous experimental studies have demonstrated that ZIKV is vertically transmitted by infected Ae. albopictus females inoculated intrathoracically with a ZIKV stock containing 106 pfu/mL, and none of 803 F1 Ae. albopictus adults (32 pools) yielded ZIKV (Thangamani et al. 2016). The current study represents the first report of the vertical transmission of ZIKV by oral infected Ae. albopictus females in China. Furthermore, vertical transmission was observed during the first gonotrophic cycle. For eggs laid on day 5 PI, the MIR was ∼6.7/1000 and 53.3/1000 for eggs and larvae, respectively. The highest MIR was observed in larvae, and 2.5% of female mosquitoes were positive, whereas parent females were 65% positive. An earlier study demonstrated the vertical transmission of WNV by Culex tarsalis mosquitoes in eggs laid on day 5 PI (Anderson et al. 2012). The results suggest that vertical transmission may provide a potential mechanism for the virus to survive during adverse conditions.
In summary, the Ae. albopictus population in China is susceptible to ZIKV and is competent for viral replication and dissemination from the salivary glands. Thus, local Ae. albopictus mosquitoes have potential roles as ZIKV vectors. Based on our results, the threat of ZIKV may be addressed by existing dengue control programs. However, the susceptibility of other common mosquito species must be evaluated to design a comprehensive vector control strategy for managing ZIKV infections.
Conclusions
The present study provides the first information on oral susceptibility and vector competence of local Ae. albopictus populations to ZIKV. Infected mosquitoes successfully transmitted the virus by biting infant mice after incubation for 8 days. Vertical transmission was observed during the first gonotrophic cycle. Thus, local Ae. albopictus mosquitoes have potential roles as ZIKV vectors.
Footnotes
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This work was funded by grants from the Infective Diseases Prevention and Cure Project of China (no. 2017ZX1000303404), the National Key Technology Support Program (no. 2014BAI13B01), and the Zika Prevention and Control Project of China.
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.