
Abstract
Dengue virus (DENV), a member of the family Flaviviridae, is the causative agent of dengue fever, the most prevalent mosquito-borne viral illness in humans, representing a major public health concern in the tropical countries. Although humoral immunity to DENV has been extensively studied and widely used, little is known about the potential diagnostic use of T cell response for DENV diagnosis. The aim of our study was to characterize the T cell immunity in subjects with acute or past DENV infection, using an original and easy to perform ex vivo ELISpot assay, and to evaluate the role of cross-reactivity between the four DENV serotypes and between DENV and Zika virus (ZIKV). As controls, DENV-seronegative healthy subjects were enrolled and a cutoff of positive DENV-specific T cell response was calculated. DENV-specific T cell response for at least one DENV serotype was detected among all DENV-specific neutralization positive subject. Furthermore, our data showed that in acute DENV infection, the DENV-specific effector memory T cell response against the relevant serotype was predominant. However, a high level of cross-reactivity among all DENV serotypes was also documented. DENV-specific T cell response was almost undetectable among DENV-seronegative subjects with ZIKV acute infection, supporting the hypothesis that the assay could be useful in differential diagnosis between ZIKV and DENV infection.
Introduction
Dengue virus (DENV) infection has become one of the major emerging infectious diseases. DENV is classified into four closely related serotypes (DENV 1–4) and infection can result in asymptomatic manifestation, dengue fever, or severe clinical manifestations (Malavige et al. 2004). Primary infection with any of the four DENV serotypes could confer long-term protection to the homologous serotype. However, a secondary infection with a different serotype represents one of the major risk factors for severe DENV infection. The protective response to DENV infection is sustained by the presence of neutralizing antibodies and specific T cell response (Stettler et al. 2016). The antibodies produced after primary DENV infection bind to the virions of subsequent infecting DENV serotypes without neutralizing effect.
This phenomenon is known as antibody-dependent enhancement (ADE) of DENV replication. Fcγ receptors recognize these virus-immune complexes, allowing the virus entry and replication in target immune cells that are responsible for the immune cascade resulting in vascular leak and severe dengue disease (Zompi and Harris 2013, Katzelnick et al. 2017). There is sufficient amino acid sequence homology within antigenically relevant regions of the viral proteins to allow cross-reactivity by memory T cells generated in response to previous infection with heterologous DENV serotype (Imrie et al. 2007, Weaver and Vasilakis 2009).
During DENV secondary infection, cross-reactive memory T cells with low activation threshold, originated from DENV primary infection, compete with naive T cells with high affinity for the secondary-infecting serotype, resulting in skewed control of viral replication. This phenomenon is known as original antigenic sin (Mongkolsapaya et al. 2003, Imrie et al. 2007).
Recently, the closely related Zika virus (ZIKV) infection has posed crucial issues in terms of differential diagnosis and clinical outcome in areas where the two viruses cocirculate. ZIKV displays an overall 43% homology of amino acid sequence with DENV (Lazear and Diamond 2016), causing high cross-reactive T cell response between structural and nonstructural proteins (Olkowski et al. 2013, Stettler et al. 2016, Xu et al. 2016). Because of their genetic relationship, a previous DENV infection could have a pathogenetic (Stettler et al. 2016) or a protective effect (Malavige et al. 2011). DENV-specific CD4+ and CD8+ T cells elicited by multiple DENV proteins have been identified (Scaramozzino et al. 2001, Weiskopf et al. 2013).
The aim of our study was to assess the DENV-specific T cell response among patients with acute or previous DENV infection in comparison with humoral response detected by neutralization assay to better evaluate the cross-reactivity between DENV serotypes and between DENV serotypes and ZIKV.
Materials and Methods
Samples
Clinical samples used in this study were collected from 43 subjects, including Italian healthy volunteers, foreign residents in Italy, and Italian travelers who were referred to our center for DENV or ZIKV infection (22 men/21 women, median age = 35.5 years, range = 18–69 years) (Table 1). The diagnostic assessment included the following: (1) detection of DENV 1–4 immunoglobulin M (IgM) and IgG antibodies in serum samples (Dengue Virus IgM Capture DxSelect and Dengue Virus IgG DxSelect; Focus Diagnostics) and ZIKV IgM and IgG antibodies (Anti-Zika Virus ELISA IgM and Anti-Zika Virus ELISA IgG; Euroimmun, Germany); (2) neutralizing antibodies assay, as described hereunder; (3) detection of DENV and ZIKV RNA in plasma and urine with a pan-flavivirus heminested RT-PCR assay targeting a conserved region of the NS5 gene (Scaramozzino et al. 2001) and DENV- and ZIKV-specific real-time RT-PCRs (Lanciotti et al. 2008, Huhtamo et al. 2010, Timiryasova et al. 2013) and sequencing of positive pan-flavivirus amplicons.
Subjects Included in the Study
DENV, dengue virus; ZIKV, Zika virus.
DENV acute infection was defined as the presence of DENV-specific IgM and/or positive DENV RT-PCR or pan-flavivirus RT-PCR, whereas DENV past infection was diagnosed as the absence of DENV-specific IgM, presence of DENV-specific IgG confirmed by neutralization test and negative molecular tests. ZIKV acute infection was defined as the presence of ZIKV-specific IgM and/or positive ZIKV RT-PCR or pan-flavivirus RT-PCR.
Analysis were performed according to guidelines of the institutional review board of the Fondazione IRCCS Policlinico San Matteo on the use of biological specimens for scientific purposes, in keeping with Italian law (art.13 D. Lgs 196/2003).
DENV-specific neutralization assay
The plaque reduction neutralization test (PRNT) was performed as follows: fourfold serial dilution of plasma samples from DENV-seropositive subjects and seronegative healthy controls were tested. On day 5 postinoculation, 50% PRNT (PRNT50) was read according to a previous procedure (Simmons et al. 2015). A neutralization titer <10 was considered negative, whereas a fourfold increased titer to one or more serotypes on two consecutive serum samples was considered specific.
Synthetic peptides
Peptide arrays were obtained by National Institutes of Health (NIH) Biodefense and Emerging Infections Research Resources Repository and National Institute of Allergy and Infectious Diseases (NIAID), NIH. In detail, NS3 peptide arrays of DENV1 Singapore/S275/1990 (NR-2752; 14–17 mers with 11 or 12 aa overlap), DENV2 New Guinea C (NR-509; 13–19 mers with 10 aa overlap), DENV3 Philippines/H87/1956 (NR-2754; 14–17 mers with 11 or 12 overlap), and DENV4 Singapore/8976/1995 (NR-2756; 15–17 mers with 11 or 12 overlap) were used. Peptides of 15–18 mers with 11 amino acid overlapping of the protein NS3 of DENV1, 2, 3, and 4 from Biodefence and Emerging Infections Research Resources Repository (BEI Resources) resources were used as pools at the final concentration of 1 μg/mL for each individual peptide in the corresponding pool.
Evaluation of DENV-specific T cell response
Peripheral blood mononuclear cells (PBMCs) were isolated by density gradient centrifugation (Lymphoprep; Axis-Shield, Oslo, Norway), cryopreserved in freezing medium (65% RPMI-1640 supplemented with 2 mM
Human interferon gamma (IFN-γ) ELISpot kits (Diaclone, Cedex, France) and MultiScreen-IP membrane-bottomed 96-well plates (Merck Millipore, Darmstadt, Germany) were used. Plates were coated overnight with monoclonal capture antibody against IFN-γ. After washing, plates were blocked with culture medium (RPMI-1640 supplemented with 2 mM
Streptavidin-alkaline phosphatase-conjugate was added after washing and plates were incubated at 37°C for 1 h. Finally, 5-bromo-4-chloro-3-indolylphosphate/nitro blue tetrazolium was added for 20 min. Wells were washed under running water and air-dried overnight. Spots were counted by using an AID ELISpot reader system (AutoImmun Diagnostika GmbH, Strasburg, Germany). The mean number of spots from duplicate were adjusted to 1 × 106 PBMC. The net spots per million PBMC was calculated by subtracting the number of spots in response to negative control from the number of spots in response to the corresponding antigen.
Characterization of DENV-specific T cell response
PBMC from selected patients were used for intracellular cytokine staining. A total of 500,000 cells were transferred to a 96-well round bottom plate in 100 μL of culture medium in the presence of the corresponding DENV antigen or staphylococcal enterotoxin B (Sigma-Aldrich) or culture medium alone.
After 1-h incubation at 37°C, 10 μL culture medium containing brefeldin A (Sigma-Aldrich) at a final concentration of 10 μg/mL were added. After 16–h incubation, cells were washed in phosphate-buffered saline (PBS) containing 2 mM ethylenediaminetetraacetic acid (EDTA) and incubated with the Live/Dead® Fixable Far Red Dead Cell Stain kit (Thermo Fisher Scientific) for 30 min at 4°C. Cells were then washed with PBS, fixed, and permeabilized with BD Cytofix/Cytoperm™ (BD Biosciences) according to the manufacturer's instructions for intracellular staining with the following monoclonal antibodies: CD3 PC5, CD4 ECD, CD8 PC7, and IFN-γ FITC (all from Beckman Coulter). Cells were resuspended in 1% paraformaldehyde and analyzed with a Navios flow cytometer (Beckman Coulter).
Data analysis
Data were reported as mean, standard deviation (SD), and standard error (SE). Comparison between groups was determined by Mann–Whitney test. Spearman's test was used for correlation analysis. All tests were two-tailed. A value of p < 0.05 was considered statistically significant. Analyses were performed using GraphPad Prism 5 (GraphPad Software, CA).
Results
DENV-specific T cell response in DENV- and ZIKV-seronegative volunteers
Serological assays were performed in serum samples from 22 healthy volunteers, confirming the absence of ZIKV and DENV IgM and IgG-specific antibodies. Neutralization assay for all the four DENV serotypes was assessed to corroborate serological results. In 22 of 22 (100%) volunteers a negative DENV-specific neutralization titer was reported (Supplementary Fig. S1a). DENV-specific T cell response was also evaluated (mean, SD and SE of T cell response to NS3 protein of DENV1, 2, 3, and 4 were 6.86 (SD = 9.5; SE = 2.03), 5.46 (SD = 11.6; SE = 2.47), 3.82 (SD = 6.52; SE = 1.39), and 6.36 (SD = 9.4; SE = 2.00) net spots/million PBMC, respectively (Fig. 1). Mean NS3 DENV-specific T cell response plus three SD in healthy DENV- and ZIKV-seronegative subjects denotes a positive response.
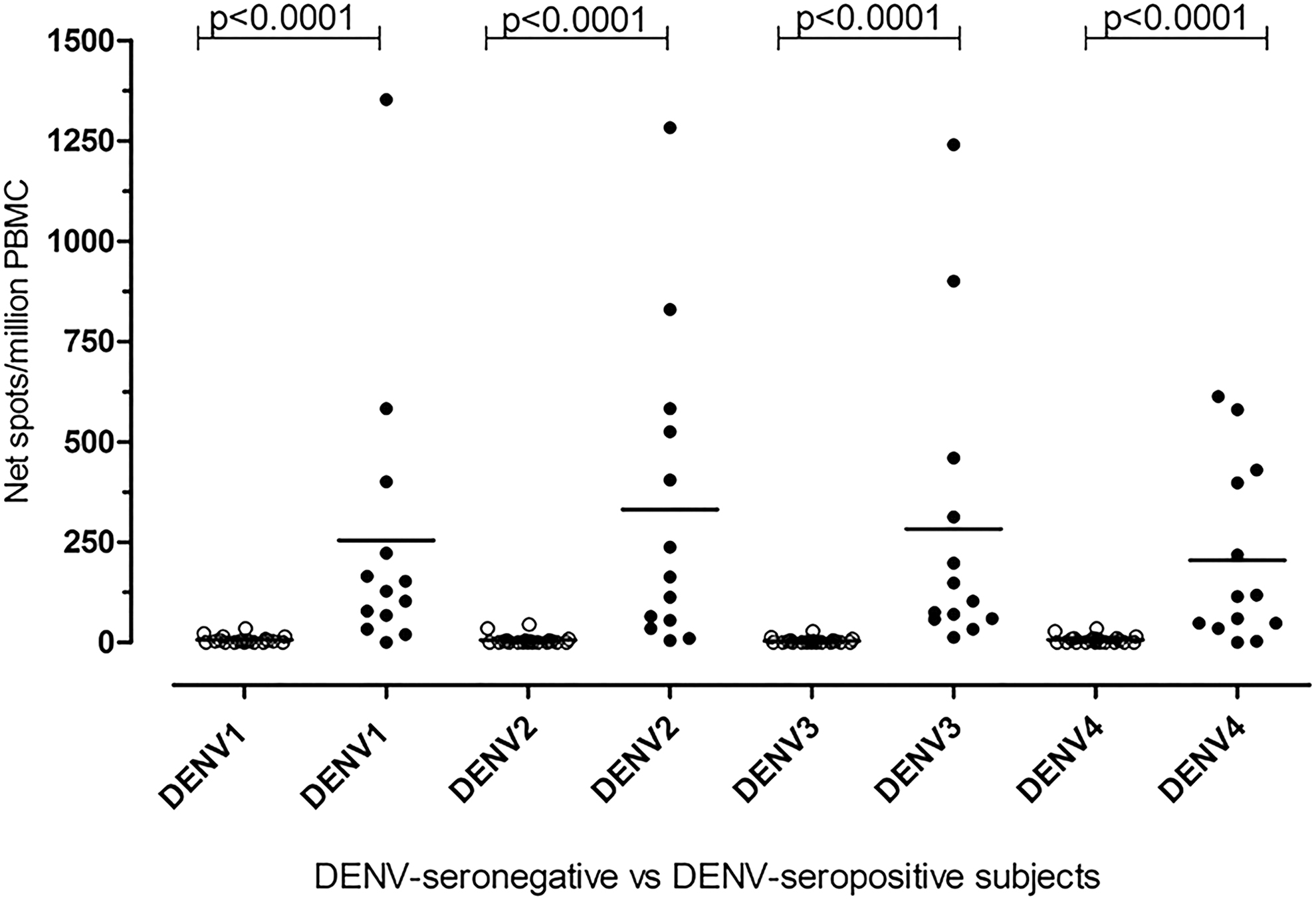
DENV-specific T cell response in DENV-seronegative and DENV-seropositive subjects. DENV1, 2, 3, and 4 NS3-specific T cell response was evaluated in 13 DENV-seropositive subjects (black dots) and 22 DENV-seronegative and ZIKV-seronegative subjects (white dots). The horizontal line represents the mean NS3-specific T cell response for each DENV serotype. DENV, dengue virus; ZIKV, Zika virus.
DENV-specific T cell response in DENV-seropositive patients
DENV-specific T cell response was quantified in 13 patients with acute or past DENV infection. Patients were divided into two groups: group A included six patients (46.2%) who were diagnosed as acute DENV infection by detection of viral RNA in blood and IgM presence; group B included seven subjects (53.8%) showing past DENV infection (IgM negative, IgG positive, and RNA negative). In this group, three patients (42.9%) presented with acute ZIKV infection (ZIKV-IgM positive and ZIKV RT-PCR positive).
Serologic results were confirmed by neutralization assay. A positive DENV-specific neutralizing titer (>40) was reported in all subjects (Supplementary Fig. S1a). In detail, mean DENV1-specific T cell response in DENV-seropositive patients (group A and group B) was 275.6 (SD = 376.4; SE = 108.7) net spots/million PBMC, whereas mean DENV2-, 3-, and 4-specific T cell responses was 358.8 (SD = 390.2; SE = 112.6), 300.1 (SD = 388.7; SE = 112.2) and 222.2 (SD = 223.1; SE = 64.4) net spots/million PBMC, respectively. As shown in Fig. 1, DENV-specific T cell responses detected in DENV-seropositive patients were significantly higher than those observed in healthy seronegative volunteers (p < 0.0001) (Fig. 1).
Focusing on DENV-seropositive patients we observed that DENV-specific T cell response was higher in those patients who have shown past and multiple DENV infection (group B), although the difference was not always statically significant (Fig. 2).

DENV-specific T cell response to NS3 protein of DENV1, 2, 3, and 4 was compared in seven patients with past and multiple DENV infections (black dots) and six patients showing acute DENV infection (white dots). The horizontal line represents the mean NS3-specific T cell response for each DENV serotype.
In addition, in patients of group A, the highest NS3 ELISpot-specific T cell response was found against the DENV serotype detected in blood (Table 2). A cross-reactivity between different serotypes was also present. Phenotypic characterization of DENV-specific T cell response from three of six DENV-seropositive patients with acute infection was investigated. In these patients, we observed that T cell response was mainly mediated by CD8+ T cells (Fig. 3a); for comparison phenotypic characterization of DENV-specific T cell response among subjects with past DENV infections were performed on residual samples (Fig. 3b). Similarly, we observed a predominant CD8+ T cell response also in subjects with past DENV infection. On the contrary, we observed that CD4+-mediated T cell response was increased in patients with past DENV infection than with respect to patients with acute DENV infection.
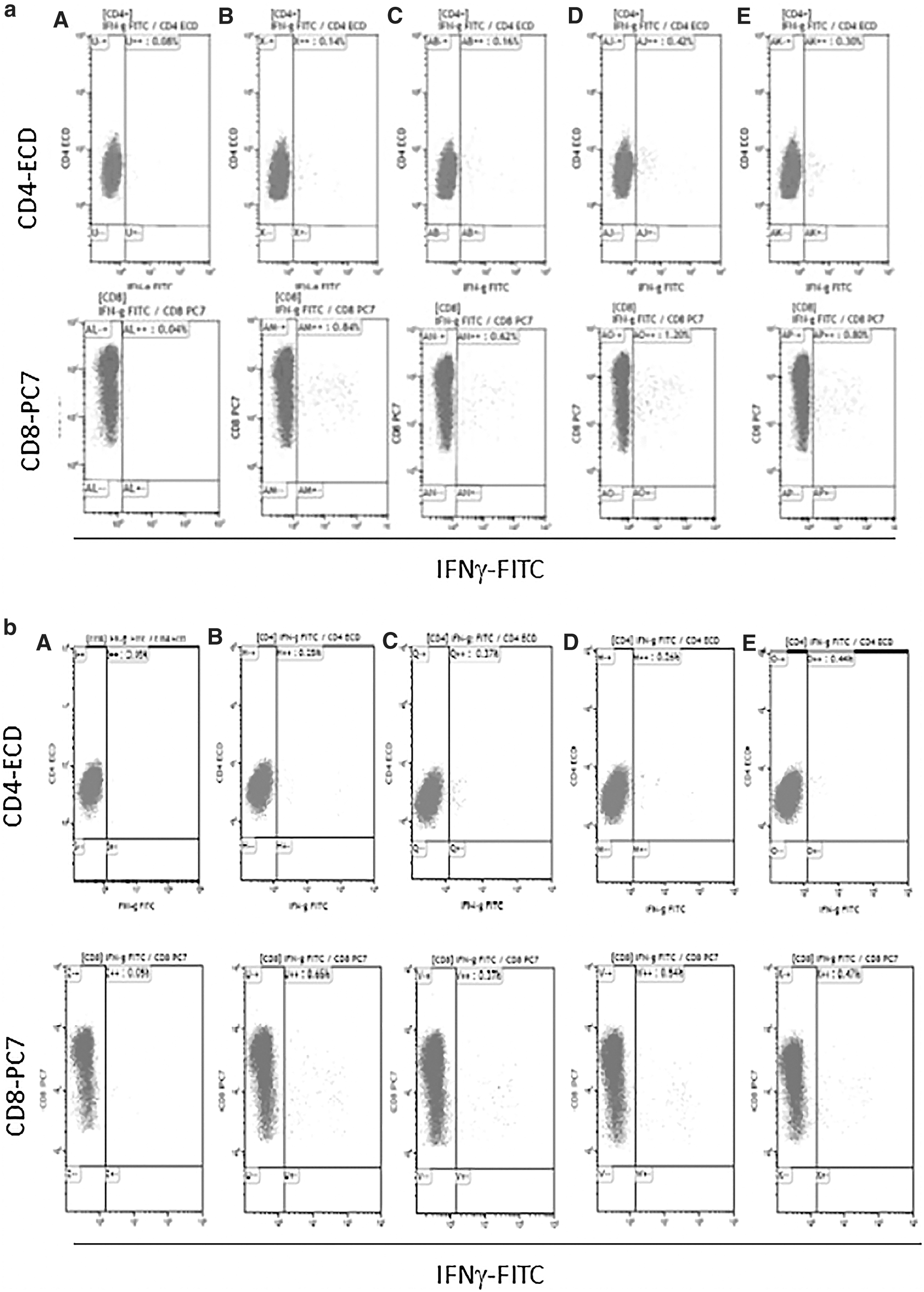
Phenotypical characterization of DENV-specific T cell response in a patient with acute DENV infection
Correlation Between DENV-Specific T Cell Response and DENV Serotype Detected in Blood in Patients with DENV-Acute Infection
The highest level of DENV-specific T-cell response for each patient are in bold.
DENV-specific T cell response is expressed as net spots/million peripheral blood mononuclear cell.
Cutoff for each DENV-specific T cell response was calculated as mean DENV-specific T cell response in healthy DENV-seronegative subjects plus three standard deviation.
DENV-specific T cell response in ZIKV-acute/DENV-naive patients
The specificity of the assay was also evaluated in eight DENV-naive patients with ZIKV acute infection (DENV-IgM and IgG negative and DENV RT-PCR negative; ZIKV-IgM positive and ZIKV RT-PCR positive). Neutralization assay was assessed; a DENV-specific negative neutralization titer was reported in 8 of 8 (100%) patients (Supplementary Fig. S1b). DENV-specific T cell response was measured and compared with DENV-specific T cell response observed in DENV-seropositive patients. Mean DENV1-specific T cell response was 9.43 (SD = 9.36; SE = 3.54) net spots/million PBMC, whereas mean DENV2-, 3-, and 4-specific T cell responses was 13.75 (SD = 9; SE = 3.18), 7.38 (SD = 7.67; SE = 2.71), and 13.88 (SD = 8.97; SE: = 3.17) net spots/million PBMC, respectively. As given in Fig. 3, a statistically significant difference among ZIKV-acute/DENV-naive patients and DENV-seropositive patients in terms of DENV1-, DENV2-, DENV3-, and DENV4-specific T cell responses (p = 0.0033, 0.0043, 0.0006 and 0.0140, respectively) was observed (Fig. 4).
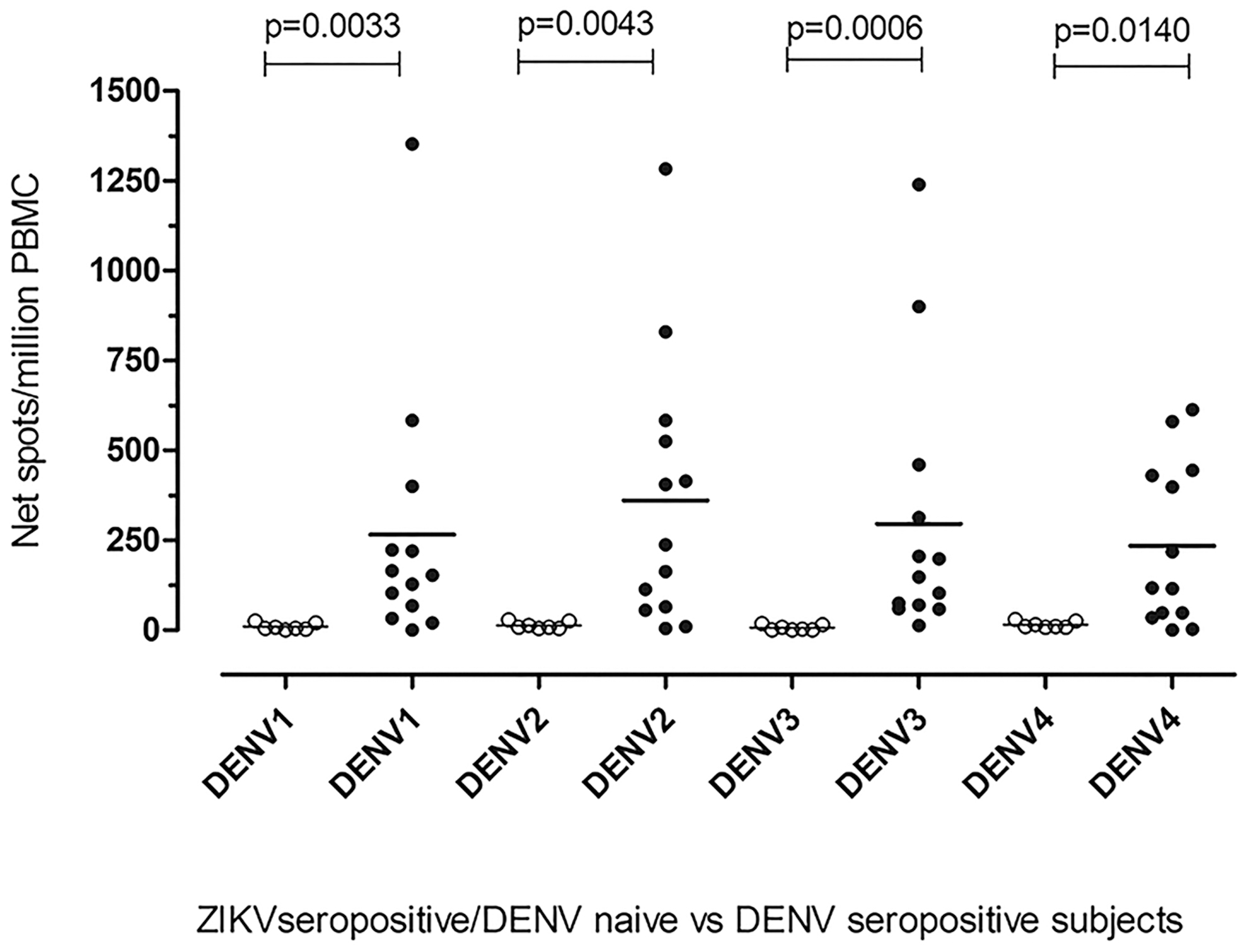
DENV-specific T cell response in ZIKV seropositive/DENV-naive and DENV-seropositive subjects. DENV-specific T cell response to DENV1, 2, 3, and 4 NS3 peptide pools was compared in ZIKV-seropositive/DENV-naive subjects (white dots) and DENV-seropositive subjects (black dots). The horizontal line represents the mean NS3-specific T cell response for each DENV serotype.
Correlation between ELISpot and neutralization assays
A significant correlation between ELISpot and neutralization assays was shown (p < 0.0001). Each DENV serotype-specific T cell response measured by ELISpot assay was in agreement with the corresponding DENV serotype-specific titer measured by neutralization assay (Fig. 5). Moreover, in immune patients with acute DENV infection, the ELISpot-specific T cell response seemed to correlate with specific titer observed in neutralization assay.
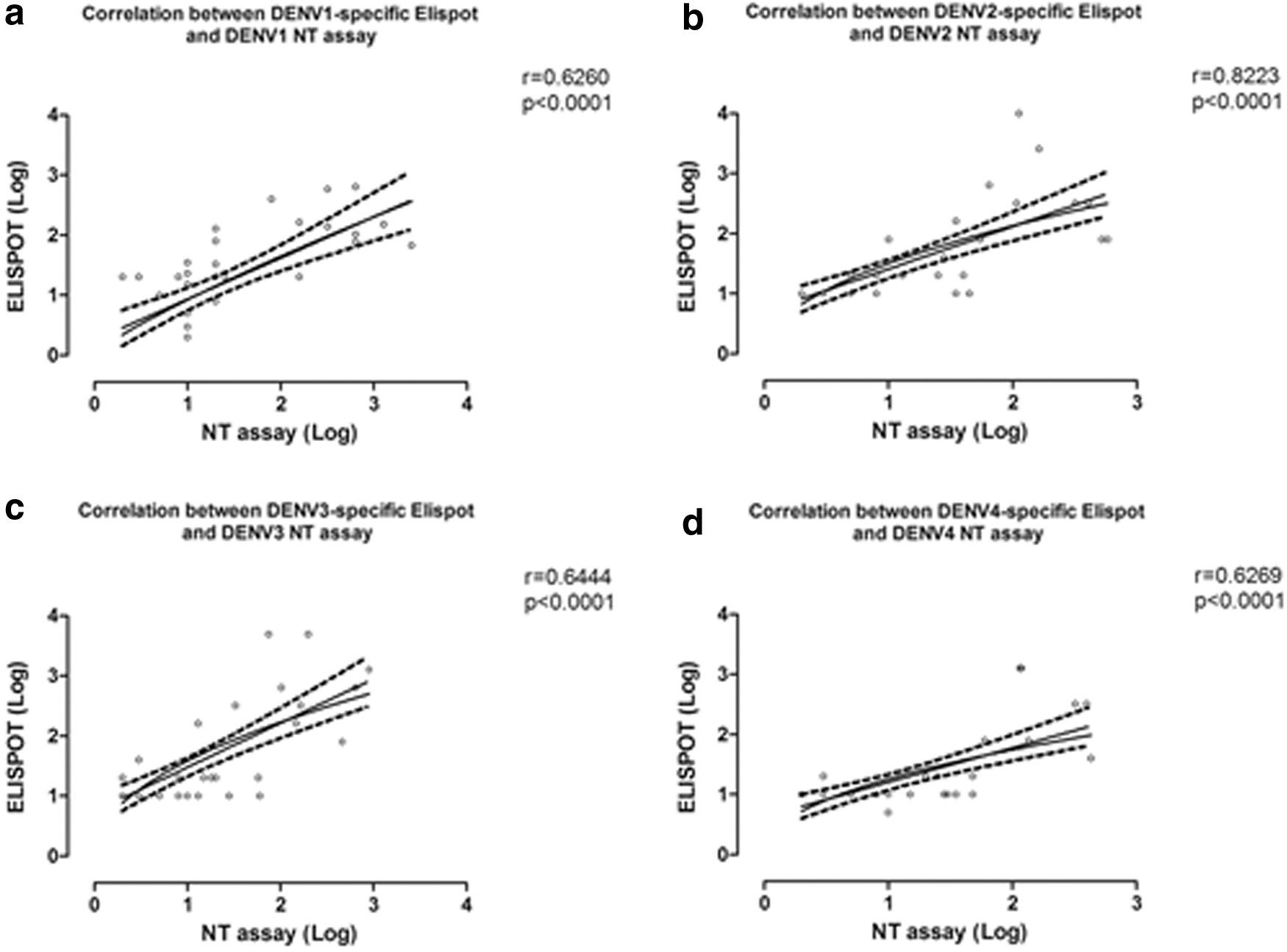
Correlation between DENV-specific ELISpot and neutralization (NT) assays. Correlation between DENV-specific ELISpot and neutralization test (NT) assays was evaluated for each DENV serotype. DENV1
Discussion
Several advancements have been achieved in characterizing humoral immunity to DENV. On the contrary, despite a significant amount of work on T cell response against DENV has been produced, the immunological evaluation of DENV-specific T cell response has not been extensively explored on the diagnostic side.
NS3 is considered one of the most immunodominant protein and many CD8+ and CD4+ T cell epitopes have been identified (Appanna et al. 2007, Moran et al. 2008, Duangchinda et al. 2010, Rivino et al. 2013, Wallace et al. 2013). We analyzed the DENV-specific T cell response against NS3 protein in a cohort of Italian subjects with a travel history in DENV-endemic countries and in Latin American residents travelling between Italy and their native country.
Unfortunately, we could examine only a small number of patients referred to our laboratory and the limitation was mainly because of the use of residual specimens after the conclusion of the diagnostic procedures.
Patients with past DENV infection were all from Latin America and showed high neutralizing titers against two or more DENV serotypes, not allowing the diagnosis of type and number of past DENV infections. At present, the role of a protective DENV-specific immune response is not well known (Coller and Clements 2011, Priyamvada et al. 2016). Previous studies have shown that cross-reactive antibody elicited by either DENV or ZIKV infection can mediate heterologous ADE (Stettler et al. 2016). Alongside, antibody responses in DENV and other flavivirus infections have also been implicated in exacerbating disease (Stettler et al. 2016).
A positive response to at least one DENV-specific NS3 peptide pool was observed among DENV-seropositive patients, suggesting that the assay could be useful for helping the diagnosis of past or acute DENV infection. Among patients with past DENV infections, it is not clear how the number and the type of past DENV infections were associated with the frequency and the functionality of T cell responses to DENV proteins. It was observed that memory T cells stimulated with heterologous virus variant antigens may display altered function compared with stimulation with related epitope peptide, resulting in a failure of the control of viral replication (Imrie et al. 2007). Of interest, in this group of patients DENV-specific T cell response was higher than that observed in patients with acute primary infections, thus helping the differential diagnosis between the two groups.
Focusing on acute DENV infections, we observed a predominant T cell response specific for the serotype involved in the infection in all the six patients examined in group A. However, cross-reactive T cell response in acute DENV infection was present as previously described (Imrie et al. 2007). Phenotypic characterization by flow cytometry of IFN-γ-producing T cells in three subjects with acute DENV infection showed a predominant CD8+ DENV-specific T cell response. According to a previous study, CD8+ T cell epitopes preferentially target nonstructural proteins (NS3 and NS5) (Wallace et al. 2013). On the contrary, it was observed that CD4+ T cells target the same proteins recognized by B cells, mainly structural proteins such as E and C (Wallace et al. 2013).
Duangchinda et al. observed that NS3 was immunodominant for both CD4+ and CD8+ T cells (Duangchinda et al. 2010). DENV-immune/ZIKV-acute patients showed a level of cross-reactivity between different DENV serotypes and ZIKV, both in humoral and T cell responses, making the establishment of the current infection difficult. It could be related to the phenomenon of “original antigenic sin,” defined as the dominance of cross-reactive antibodies or T cell response to the first infecting DENV serotype over the current infecting one (Zompi and Harris 2013). Because ZIKV is strictly related to DENV, the phenomenon of “original antigenic sin” could be observed also in DENV-seropositive patients with ZIKV infection.
The sequential exposure of these patients to the two viruses generates a response against conserved sites between the viruses. Of interest, as previously described, we observed that all ZIKV-seropositive patients who did not experience previous DENV infection had a DENV-specific T cell response lower than the established positive response cutoff, calculated from DENV-specific T cell response detected in DENV-seronegative healthy controls. In ZIKV-seropositive/DENV-naive patients, DENV-specific T cell response was always significantly lower than that observed among DENV-seropositive patients. In this setting, the assay could be useful for the diagnosis of primary ZIKV infection in DENV-naive patients.
We also observed a positive correlation between neutralization assay and ELISpot results. A better correlation was observed for DENV2-specific T cell response and neutralization assay, followed by DENV3, DENV1, and DENV4. However, despite the significant correlation observed between T cell response and neutralizing antibodies, both the neutralizing antibodies and T cell-mediated immunity seem to be insufficient for the differential diagnosis between DENV and ZIKV when a previous DENV immunity is present.
Conclusions
The major limitation of the present study is mainly related to the low number of patients recruited, even if our patient groups represent a real setting for a reference European laboratory.
Although the small number of patients enrolled in this study can represent a limitation for the strength of the results, it is conceivable that they can represent an important starting point for further analysis on a larger group of patients.
Indeed, it is believable that immunological assays for the evaluation of DENV-specific T cell response should be further investigated to evaluate the potential usefulness for the assessment of cross-reactivity between different flavivirus infection in a diagnostic setting.
Footnotes
Acknowledgments
The authors thank Daniela Sartori for editing of the article and Sandra A. Calarota for English revision.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This study was supported by funds from Lombardy Region and from the Ministero della Salute, Ricerca Corrente Fondazione IRCCS Policlinico San Matteo, grant no. 80206 to Dr. Elena Percivalle.
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.