
Abstract
Although malaria is endemic to the Amazon region, little is known about the susceptibility of potential parasite vectors in Brazil. Assessing the vector susceptibility of Anopheles mosquitoes will increase our understanding of parasite–vector interactions and aid the design of vector control strategies. This study assessed the susceptibility of three Anopheles species to midgut infection by Plasmodium vivax, the predominant malaria species in Rondônia State, Brazil. Blood from P. vivax infected patients was fed to Anopheles aquasalis, Anopheles darlingi, and Anopheles deaneorum mosquitoes using a membrane feeding assay (MFA). Gametocytemia was estimated by microscopic examination of blood smears and oocyst prevalence, and infection intensity was assessed. The presence of oocysts was determined by microscopy, and the infection rates and infection intensity were determined for all species. Data from six MFAs showed that An. darlingi and An. deaneorum exhibited the highest infection rates (97% and 90%, respectively) and developed a similar median number of P. vivax oocysts (142 and 123, respectively), while An. aquasalis exhibited the smallest infection rates (77%) and the median number of oocysts (88). Established laboratory colonies of An. darlingi and An. deaneorum and susceptibility to plasmodial infection would be beneficial for modeling P. vivax vector–parasite interactions in Brazil.
Introduction
Despite huge efforts by the global community, malaria remains a significant public health problem worldwide; in 2017 alone, there were 219 million cases of malaria and 435,000 related deaths (WHO 2018). In Brazil, malaria cases increased 19% from 2016 to 2017, with 99.8% of cases occurring in the Amazon region (SIVEP, 2018).
More than 450 species of Anopheles have been described; 30 of these species occur in the Brazilian Amazon (Harbach and Kitching 2016), but only Anopheles darlingi, Anopheles aquasalis, and Anopheles albitarsis s.l. are considered primary vectors of human Plasmodium (Pimenta et al. 2015).
Anopheles darlingi is widely distributed in the Americas. Its highly anthropophilic behavior and capacity to adapt to environmental changes make it one of the most efficient malaria vectors (Deane et al. 1948, Barros et al. 2007, Manguin et al. 2008). This species comprises over 90% of the Anopheles population in several endemic locations within the Brazilian Amazon (Deane 1988, Tadei and Thatcher 2000, Gil et al. 2003, 2007, 2015, Moutinho et al. 2011). Seasonality patterns of An. darlingi are closely related to the annual cycle of rainfall and to the nature and availability of suitable water bodies to use as breeding sites (Gil et al. 2007, Moutinho et al. 2011).
In laboratory experiments, An. darlingi exhibits higher susceptibility to Plasmodium infection than other Amazonian species (Klein et al. 1991a, 1991b). However, the natural infection rate of An. darlingi ranges from 0.1 to 4.2% (Arruda et al. 1986, Tadei et al. 1988, Oliveira-Ferreira et al. 1990, Schoeler et al. 2003, Flores-Mendoza et al. 2004, Gil et al. 2007). This natural infection rate is considered low compared to African vectors, such as Anopheles funestus (s.l.) and Anopheles gambiae (s.l.), in which infection rates reach 22% (Ndo et al. 2018). Despite its natural infection rates tending to be low, it is known that this mosquito species is capable of maintaining a relatively high transmission of malaria even when found in low densities (Deane et al. 1948, Póvoa et al. 2006).
Anopheles aquasalis is also a primary vector of both Plasmodium vivax and Plasmodium falciparum, but its distribution is limited to regions that have brackish water where immature stages breed. Anopheles aquasalis is considered to be more zoophilic than anthropophilic, which may have an impact on its vectorial capacity (Deane et al. 1948, Da Silva et al. 2006b). Although An. aquasalis is not highly susceptible to Plasmodium, it can maintain malaria transmission when it occurs in high density, especially during the rainy season (Forattini 2002). Its natural infection rate ranges from 0.26 to 1.18% (Galvão et al. 1942, Póvoa et al. 2003). Anopheles aquasalis is considered a secondary malaria vector in some areas of the Amazon and southeastern Brazil because its vector status is dependent on species density (Flores-Mendoza and Lourenço-de-Oliveira 1996).
Anopheles deaneorum is a member of the An. abitarsis complex and was described as responsible for malaria transmission in agricultural settlement and riverine region of Acre State, Brazil (Branquinho et al. 1993). Like An. darlingi, An. deaneorum is highly anthropophilic and frequently found inside human dwellings (Klein and Lima 1990, Klein et al. 1991c). Mosquitoes of this species were found to be naturally infected with both P. vivax (1.17%) and P. falciparum (2.76%) (Branquinho et al. 1993).
Anopheles deaneorum is considered a potential vector in Brazil and its susceptibility to infection is comparable to that of An. darlingi (Klein et al. 1991a, 1991b). Anopheles deaneorum has been recorded in the Brazilian Amazon region, in western Paraná and São Paulo, and in Paraguay, Bolivia, and Argentina (Klein et al. 1991c, Branquinho et al. 1993, Li and Wilkerson 2005). Although the exact distribution of An. deaneorum is unknown, An. deaneorum may be a primary malaria vector in other parts of Brazil (Klein et al. 1991b).
Knowledge of vector–parasite interactions aids the development of control strategies. Unfortunately, studies of the P. vivax life cycle have been hampered in the Amazon region by a lack of parasites in continuous culture and by the scarcity of laboratory colonies of P. vivax susceptible Anopheles (Vallejo et al. 2016).
Anopheles aquasalis is the only species that has been colonized under laboratory conditions in Brazil for an extended period of time. As a result, An. aquasalis has been the sole vector for modeling vector–parasite interactions in studies that have used membrane feeding assay (MFA) with blood from malaria patients in endemic areas (Rios-Velásquez et al. 2013, Martins-Campos et al. 2018). In previous studies, An. deaneorum has been colonized using the forced mating technique (Klein et al. 1990), and An. darlingi has been maintained as a free-mating colony (Moreno et al. 2014, Villarreal-Trevino et al. 2015).
Relative susceptibility to P. vivax infection has been assessed previously among An. aquasalis with four groups of field-collected Anopheles species (An. darlingi, An. albitarsis s.l., An. nuneztovari s.l., and An. triannulatus s.l.) (Rios-Velásquez et al. 2013) and for An. darlingi with seven other anopheline species (An. deaneorum, An. albitarsis, An. mediopunctatus, An. triannulatus, An. oswaldoi, An. brasiliensis, and An. benarrochi) (Klein et al. 1991b). This is the first study that assesses the P. vivax susceptibility of An. aquasalis, An. darlingi, and An. deaneorum by MFA. Susceptibility to P. vivax was assessed on the basis of oocyst count. Adaptability to laboratory conditions and susceptibility to plasmodial infection suggested that An. darlingi and An. deaneorum can also be used as a vector model to study the P. vivax interaction with an American mosquito vector.
Materials and Methods
Ethics
All experiments followed the guidelines laid out by the Ethics Committee of the Research Center of Tropical Medicine (CEPEM) (CAAE: 26302113.4.0000.0011). Samples from volunteers were anonymized and not linked to the identity of the donors. The volunteers in this study were all adults (>18 years of age) who were able to read and to sign the informed consent forms. The decision to participate had no effect on malaria treatment.
Blood collection and parasitemia
Plasmodium vivax-infected human blood samples were collected from symptomatic patients seeking malaria diagnosis at the Centro de Medicina Tropical de Rondônia—CEMETRON (located in the city of Porto Velho). Malaria infection was confirmed microscopically. A total of 10 mL of blood was drawn from each volunteer by venipuncture using heparinized Vacutainer tubes and maintained at 37°C. Following blood collection, all patients were treated at the CEMETRON in accordance with ethical procedures designed by the Brazilian Health Ministry. This study excluded the following: patients with zero gametocytes by thick blood-smear count, patients with severe or complicated malaria, patients with serious comorbid conditions (e.g., HIV/AIDS or malnutrition), pregnant women, children, and indigenous people.
Gametocyte density was calculated as the number of gametocytes per 200 leukocytes, assuming an average number of 6000 leukocytes/μL. Counts were performed under light microscopy with a 100 × immersion oil lens (Shute 1988). Gametocytes were counted by at least two trained microscopists working independently.
Mosquito rearing
To obtain F1 generation mosquitoes of An. darlingi and An. deaneorum, wild-caught mosquitoes were fed on anesthetized laboratory mice to induce egg laying. Mosquitoes were collected using the BG-Malaria trap (Gama et al. 2013) and protected human landing. Collections were made at dusk (from 6:00 to 9:00 PM), during April and July of 2018, in peri-domicile environments near Porto Velho (08°39.145′ S/063°56.155′ W) and São Francisco do Guaporé (8°39′8. 874′ S/63°56′8.106 W), in Rondônia State, Northwestern Brazil. Anopheles darlingi was relatively abundant near Porto Velho, while An. deaneorum was more abundant near São Francisco do Guaporé.
Mosquitoes from field were first anesthetized with ethyl acetate solution and then morphologically identified under stereomicroscope using morphological identification keys of Consoli and Lourenço-de-Oliveira (1994) to screening only An. darlingi and An. deaneorum species. Seventy-two hours after blood feeding, oviposition was induced by removing one wing from each mosquito (Lanzaro et al. 1988). A group of 15 females of each species were placed on plates with wet filter paper for egg laying, as previously described (Villarreal-Trevino et al. 2015). After hatching, the larvae were reared in plastic trays and fed with fish food (TetraMin® Tropical Flakes) until they pupated, according to the protocol for An. darlingi described by Araújo et al. (2012) and the protocol for An. deaneorum described by Klein et al. (1990).
Anopheles aquasalis mosquitoes originated from a colony established in 1995 and were reared from eggs to adults as previously described (Da Silva et al. 2006a).
Pupae were removed and placed in emergent cups and were placed in screened cages (35 × 35 × 35 cm). Emerging adults were fed on a 10% sucrose solution before and after the infective bloodmeals and maintained in controlled conditions at 26 ± 1°C and 70 ± 1% relative humidity for a 12-h day/12-h night photoperiod. All experiments used adult females 3–5 days old.
Mosquito blood-feeding experiments
Adult mosquitoes were subjected to overnight fasting before infection using an artificial membrane feeding system (Rios-Velásquez et al. 2013). Two milliliters of infected blood was added to a 5 cm diameter handblown glass feeder. Laboratory-reared mosquitoes were placed in separate cages (25–50 mosquitoes/cage) and given 90 min to feed on the membrane feeder. Unfed mosquitoes were discarded. Fully engorged mosquitoes were transferred to rearing containers and maintained in the insectary at 26 ± 1°C and 70 ± 1% relative humidity and fed a 10% sugar solution daily.
Infected mosquitoes
On day 7, surviving mosquitoes were dissected in phosphate buffered saline. Midguts were stained with 0.2% mercurochrome, placed under a cover glass, and examined for the presence of oocysts. The number of oocysts per midgut was counted under normal light microscopy (40 × ) to estimate oocyst prevalence (percentage of mosquitoes infected) and infection intensity (number of oocysts/mosquito).
Data management and statistical analyses
Descriptive analysis was used to summarize the results of proportion of mosquitoes engorged and proportion of mosquitoes which survived until the time of dissection. Infection rate was obtained dividing the number of infected mosquitoes by the number of dissected mosquitoes. The number of oocysts found in midgut of infected mosquitoes was used as measure of infection intensity. Point estimates and credible interval of infection rate and infection intensity were obtained using Bayesian model. To compute the posterior 15,000 samples were drawn using NUTS algorithm and two chains. The first 5000 were used to tune the algorithm than sample. The analyses were performed using Python 3.7 and with the libraries pymc3, pandas, numpy, and plotnine. All the codes were available as Supplementary Data (S1 and S2).
Results
Paired-feeding assays using three different mosquito species were performed using six blood samples from P. vivax-infected donors. In all blood samples gametocytes were identified, varying between 120 and 1050 gametocytes per μL with mean of 485 ± 138.9 SD and median of 465. About 700 female mosquitoes were used in the MFA (25–50 females per experiment) of which more than 400 were dissected (15–40 females per experiment).
The engorged feeding percentages of An. aquasalis, An. darlingi, and An. deaneorum were 78.0% (195/250), 80.5% (149/185), and 90.6% (204/225), respectively (Table 1). The number of mosquitoes available for examination of midgut depended on the proportion that blood was fed and survival. Anopheles darlingi had the lowest proportion of mosquitoes which survived until the time of midgut dissection, 77.8% (116/149), followed by An. deaneorum (91.2%; 186/204) and An. aquasalis (99.0%; 193/195).
Infection of Anopheline by Plasmodium vivax
The proportion of mosquitoes infected was different among species: An. darlingi showed the highest infection rate (mean value: 0.97; credible interval [CrI]: 0.95–0.99), followed by An. deaneorum (mean value: 0.90; CrI: 0.86–0.94) and An. aquasalis (mean value: 0.77; CrI: 0.71–0.82) (Table 1; Fig. 1A). The results of comparative susceptibility showed that An. darlingi was about 20% more susceptible than An. aquasalis (Fig. 1B). Likewise, An. deaneorum demonstrated higher susceptibility than An. Aquasalis; in this case the difference was about 14% (Fig. 1C). And between An. darlingi and An. deaneorum, the difference of susceptibility was 7% higher for An. darlingi (Fig. 2D).
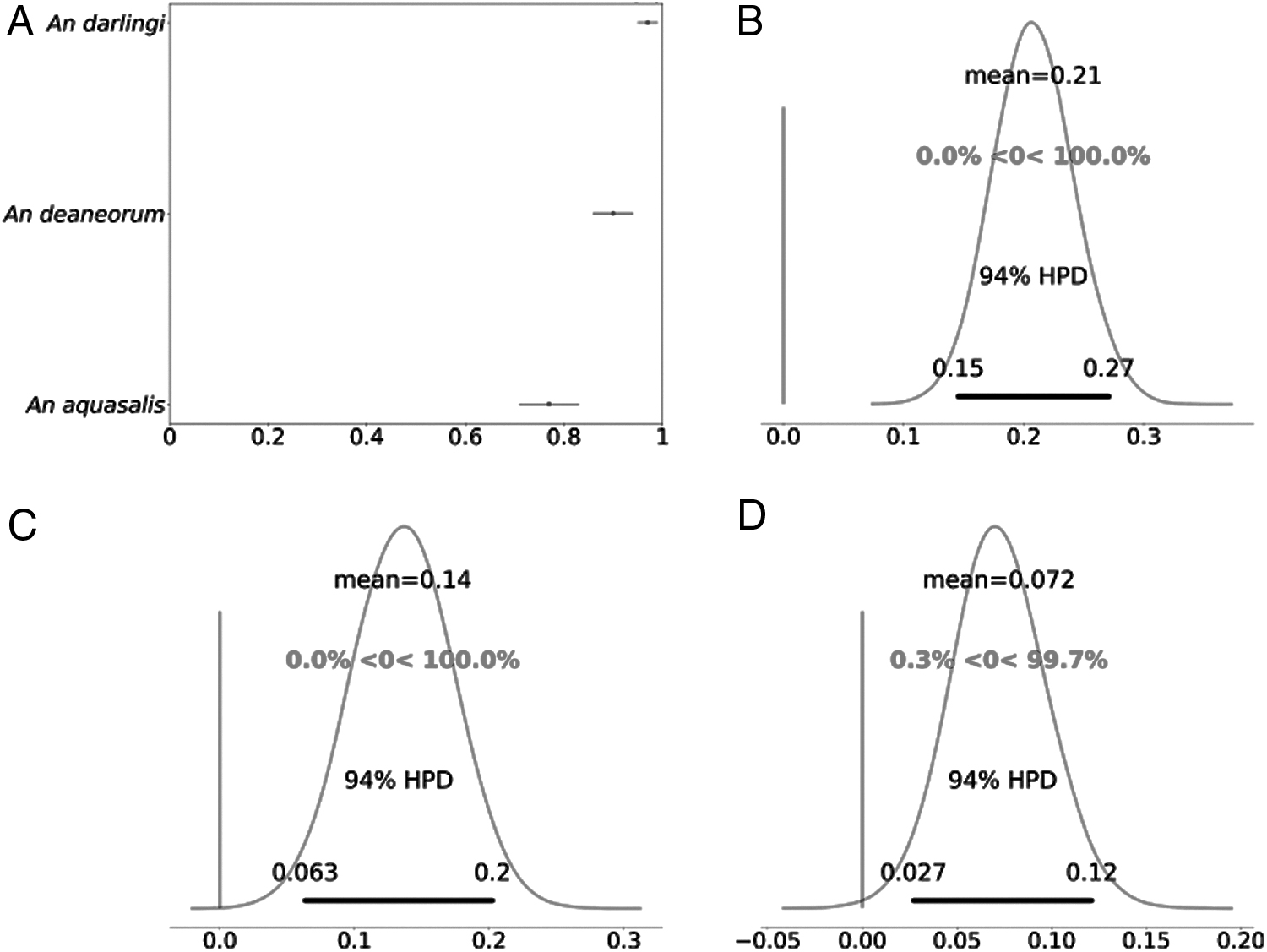
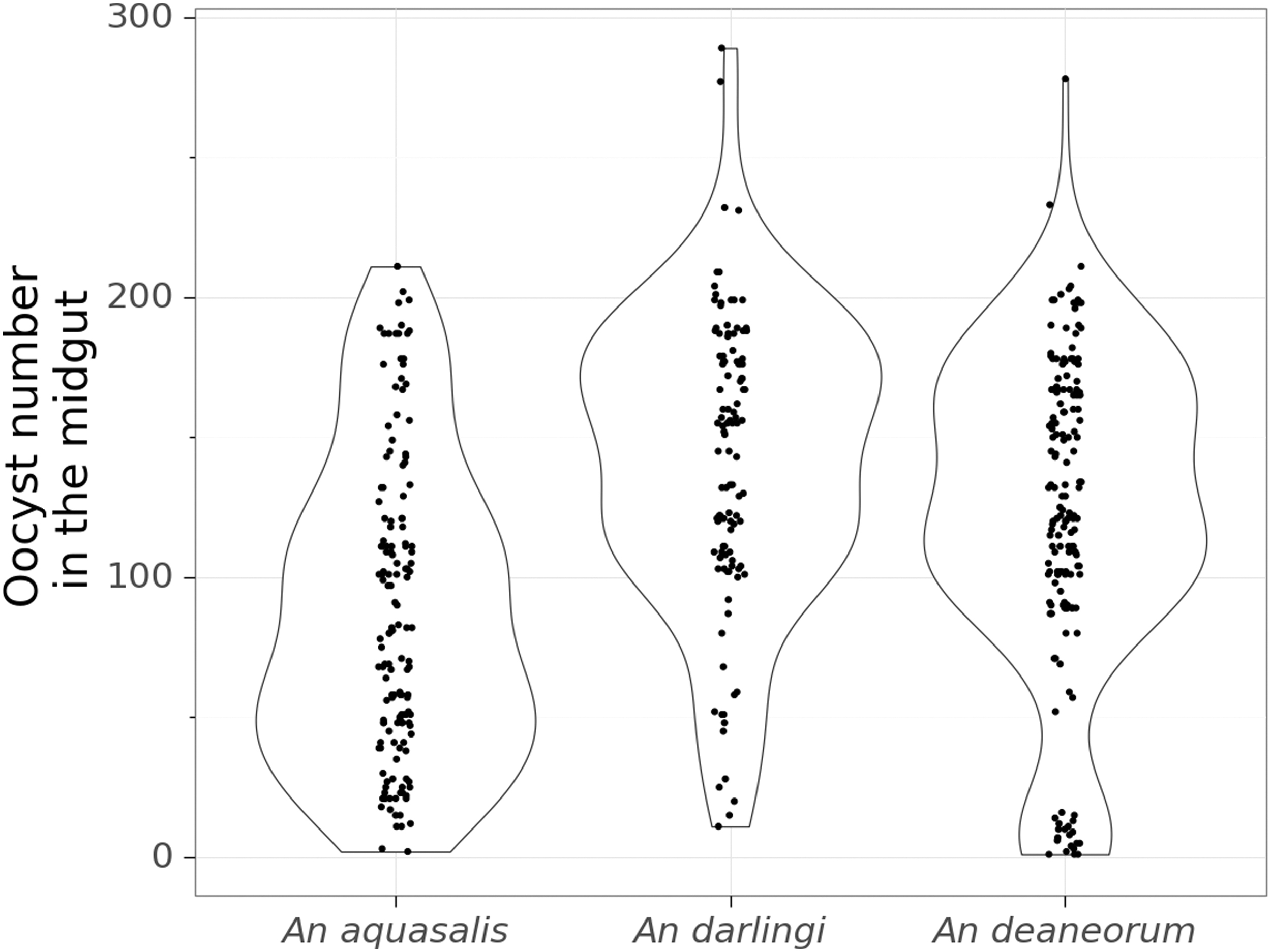
Violin plot of the oocysts number found in the midgut of each species after the blood-feeding experiments. Total number of mosquitoes included in the analysis for oocysts/midgut (n) = 149 for An. Aquasalis, (n) = 114 for An. darlingi, and (n) = 169 for An. deaneorum.
The intensity of infection, measured by the numbers of oocysts per infected mosquito, ranged from 1 to 289. The highest median oocyst count was observed in An. darlingi (median value: 142; CrI: 139–144), followed by An. deaneorum (median value: 123; CrI: 121–124) and An. aquasalis (median value: 88; CrI: 86–89) (Table 1; Fig. 2). An average difference of 53.8 (CrI: 51–56.3) oocysts/mosquito was found between An. darlingi and An. aquasalis; 34.8 (CrI: 32.6–37.0) oocysts/mosquito between An. deaneorum and An. Aquasalis; and 18.9 (CrI: 16.1–21.6) oocysts/mosquito between An. darlingi and An. deaneorum.
Discussion
Experimental P. vivax infection of three malaria vectors from South America was performed through MFA using patient blood containing gametocytes under laboratory conditions. All the three species were susceptible to P. vivax infection as evidenced by presence of oocysts into midgut. However, the species which showed the highest infection rate were An. darlingi and An. deaneorum, compared with An. aquasalis. Oocyst numbers (intensity infection) in An. aquasalis also were lower than An. darlingi and An. deaneorum, although the median of oocysts was 88 oocysts/mosquito.
The experimental infection of mosquito vectors can be achieved either by direct feeding on a patient's skin or by offering bloodmeal through a membrane-feeding device. This is the first study to evaluate the infection of An. deaneorum using MFA and to compare the P. vivax infection rates of An. darlingi and An. aquasalis. Our findings corroborate previous studies that have compared the susceptibility of Brazilian Amazon anopheline.
In a study where mosquitoes fed directly on P. vivax patients, An. darlingi and An. deaneorum developed a similar number of oocysts (over 100 oocysts per mosquito) and exhibited the highest oocyst infection rates (97–82%) relative to six other Anopheles species (which did not include An. aquasalis) (Klein et al. 1991b). In a study that assessed the susceptibility of An. aquasalis and An. darlingi to two strains of P. vivax, An. darlingi exhibited a higher infection rate than An. aquasalis (93% vs. 53%) and a higher mean number of oocysts (Da Silva et al. 2006b). However, recent studies of Neotropical anophelines have observed significantly higher infection rates in An. aquasalis and An. albitarsis than in An. darlingi (Rios-Velásquez et al. 2013).
Artificial infection rates have been shown to vary in relation to study area and Plasmodium species (Bharti et al. 2006, Kiattibutr et al. 2017, Martins-Campos et al. 2018), and infection rates may also be affected by rearing conditions. Under laboratory conditions, larval diet affected mosquito development and susceptibility to Plasmodium (Linenberg et al. 2016).
It is well known that malaria susceptibility differs among mosquito species. Propensity to infection is influenced by parasite species and strain, environmental conditions, biological factors (such as midgut and salivary gland barriers), and behavioral factors (such as anthropophilic vs. zoophilic feeding) (Adak et al. 1999, Kaur et al. 2000, Abduselam et al. 2016). The blood-host preference of An. aquasalis varies by locality in Brazil. Anopheles aquasalis seems to be anthropophilic in the northeast (Deane et al. 1948, Rachou et al. 1950), but predominately zoophilic or opportunistic in the Amazon and other regions (Galvão et al. 1942, Flores-Mendoza et al. 1996). This habit variation has influenced An. aquasalis' performance as a malaria vector (Flores-Mendoza et al. 1996).
Moreover, An. aquasalis is the only species that has been colonized for several years under laboratory conditions (Da Silva et al. 2006a) and that may affect its susceptibility. Colonized mosquito population can lose alleles over a period of several generations, potentially affecting biological interactions with and response to a pathogen (Norris et al. 2001, Lainhart et al. 2015). Anopheles aquasalis mosquitoes used for MFA were colonized at our laboratory for a longer time, while An. darlingi and An. deaneorum mosquitoes were laboratory reared F1 generation.
Another factor that may have significant influence on the Plasmodium susceptibility is mosquito gut microbiota (Dong et al. 2009). It is possible that the altered gut microbiota in colonized mosquitoes may elicit stronger basal immunity than in wild ones (Mohanty et al. 2018). However, we have not explored this in the present study.
The number of mosquito specimens available for experimental infection was limited because we did not have access to laboratory colonies of An. darlingi and An. deaneorum; however, at the time, we were using natural copulation induction (Moreno et al. 2014, Villarreal-Trevino et al. 2015) and the forced copulation method (Klein et al. 1990) to establish laboratory colonies of both species.
Evaluating the variable susceptibility of An. aquasalis, An. darlingi, and An. deaneorum will require future studies designed to assess gametocyte maturity and gender ratio, the role of midgut and salivary gland barriers, immune factors that may affect the ability of P. vivax gametocytes to infect mosquitoes, and the effect of different Plasmodium genotypes on transmission. Comparative studies of this kind are now possible because our laboratory has recently established colony of An. darlingi (Araújo et al. 2019) and An. deaneorum is in process of colonization. Previously, the only established laboratory colonies in Brazil were An. aquasalis colonies. Anopheles deaneorum and An. darlingi are ideal subjects for modeling vector–parasite interactions because both species exhibit high susceptibility to P. vivax infection and both develop high numbers of oocysts.
Footnotes
Author's Contributions
M.S.A. and J.F.M. were involved in interpreting data and preparing the article; A.O.A., N.A.C.S., R.B.C., A.M.P.J., L.P.C.C., G.S.C., and M.S.A. reared the mosquitoes; A.O.A. and M.S.A. performed the experiments; M.M.S.R. performed the data analysis; and D.B.P. met and treated the patients. All authors read and approved the final version of this article.
Acknowledgments
The authors thank: the malaria team at CEPEM; Franklin Sa. Barbosa; Antonio M. Gurgel; and Rogério L. de Souza for making and reading the malaria thick smear; the technicians Rwan R.M.O. Costa and Martina R. Lobato for recruiting patients and blood collection; and the technicians at the Fiocruz Rondônia Entomology Lab, Raimundo Nonato and Marlon Simplício, for rearing mosquitoes. The authors especially thank all the unnamed malaria patients who participated in this research.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This study was funded by the Fundação de Amparo à Pesquisa do Estado de Rondônia (Fapero) (grant N02/2014) and by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior—Brasil (CAPES)—Finance Code 001.
Supplementary Material
Supplementary Data S1
Supplementary Data S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.