
Abstract
Crimean-Congo hemorrhagic fever (CCHF) is a highly lethal infectious disease in humans caused by tick-borne Crimean-Congo hemorrhagic fever virus (CCHFV). To determine the potential risk of CCHF in the pastoral area of Northwestern China, the presence of antibody to CCHFV in the sera of two most common tick hosts, sheep and camels, in Inner Mongolia of China was evaluated. The purified recombinant nucleocapsid protein (NP) of CCHFV was prepared from a prokaryotic vector expressing the NP encoding gene, which was employed as the antigen to probe antibody against CCHFV in tick vectors using an immunoblotting assay. In total, 58.3% (35/60) and 54% (12/22) of camels were positive of antibody to CCHFV in sera at Alxa Youqi County and Ulan Hudu Gacha Village of Inner Mongolia Province, respectively. However, only 6.7% (2/30) of sera of sheep were determined positive in antibody to CCHFV in the Wulan Hudug check area in this study. Consequently, these results indicate that 54–58.3% camels were infected by CCHFV after exposure to tick bites in Inner Mongolia, which was significantly higher than 6.7% of infection in the sheep in this area, suggesting there is a certain relationship between the serological reactivity and exposure time to ticks, range of activity, living behaviors, and breeding time. Further intensive surveillance of livestock and exposed population is required to better understand the spread of CCHFV in this area.
Introduction
Crimean-Congo hemorrhagic fever virus (CCHFV) is an important zoonotic viral pathogen causing acute hemorrhagic fever. The global mortality rate of human infected by this virus is up to 30–50% (Messina et al. 2015). Pathogenetically, CCHFV is a single negative-stranded RNA virus belonging to the genus Orthonairovirus, Nairoviridae, and Bunyavirales. The genome of CCHFV is divided into three segments: large (L), medium (M), and small (S) segments that are responsible for encoding respective RNA-dependent RNA polymerase (RdRp), glycoprotein (G) precursor, and nucleocapsid protein (NP) (Zhang et al. 2018). Based on its S gene, CCHFV can be further divided into seven different evolutionary branches (Hewson et al. 2004, Shahhosseini et al. 2017). Epidemiologically, the CCHFV is widely distributed and reported in many countries and regions around the world. The infection has been mostly reported in Africa countries including Uganda (Balinandi et al. 2018), Mauritania (Sas et al. 2017), South Africa (Vawda et al. 2018), and Nigeria (Bukbuk et al. 2016). It was also reported in European countries such as Hungary (Németh et al. 2013), Macedonia (Schuster et al. 2017), Russia (Lukashev and Deviatkin 2018), Bulgaria (Christova et al. 2017), Kosovo (Jameson et al. 2012), Turkey (Albayrak et al. 2012), and Greece (Papa et al. 2014), as well as the Middle East and other Asian countries including Iran (Champour et al. 2016), United Arab Emirates (Khan et al. 1997), Iraq (Tantawi et al. 1981), Saudi Arabia (Elazazy and Scrimgeour 1997), Oman (Williams et al. 2010), China (Sun and Dai 2009), Kazakhstan (Barbara et al. 2012), Mongolia (Voorhees et al. 2018), India (Mourya et al. 2012), Uzbekistan (Lyudmila et al. 2003), and Pakistan (Masroor et al. 2013).
Ticks are the common vectors for CCHFV transmission. Indeed, CCHFV has been isolated from various species of ticks, including Hyalomma dromedarii (Yaser et al. 2011), Hyalomma marginatum (Gergova and Kamarinchev 2013), Hyalomma lusitanicum (Estrada-Pena et al. 2012). Rhipicephalus bursa (Sherifi et al. 2014), Rhipicephalus sanguineus (Gergova and Kamarinchev 2013), and Rhipicephalus eversti (Sas et al. 2017). Human or animals can be infected by CCHFV via virus-borne tick biting. In addition, humans exposed to the blood of CCHFV-infected mammals without a protection may lead the infection of CCHFV (Mertens et al. 2016).
Inner Mongolia is a province located in North China adjacent to Mongolia and Russia, where grassland is well developed, and domestic animals such as camels, cattle, horses, and sheep are highly threatened vectors of ticks. It has been well established that domestic ruminants play an important role in the life cycle of ticks as viral vectors and in the transmission of viruses. The prevalence of antibodies in these animals is an indirect indicator of the presence of CCHFV. To investigate the epidemic situation of CCHFV in animals of this area, the antibody to CCHFV in the sera of camels and sheep in Inner Mongolia was, therefore, examined.
Materials and Methods
Animal blood and ticks sample collection and metagenomic analysis
Sixty samples of camel serum were collected from Youqi County in Inner Mongolia Province (Location A in Fig. 1). Twenty-two camel and 30 sheep serum samples were collected from Ulan Hudu Gacha Village in Inner Mongolia (Location B in Fig. 1). At the same time, 5000 ticks were collected from camels and sheep at these 2 locations. Morphological grouping and molecular identification (Cyt b gene) indicated that these ticks belonged to the Hyalomma asiaticum. All samples were stored at −80°C till further analysis.
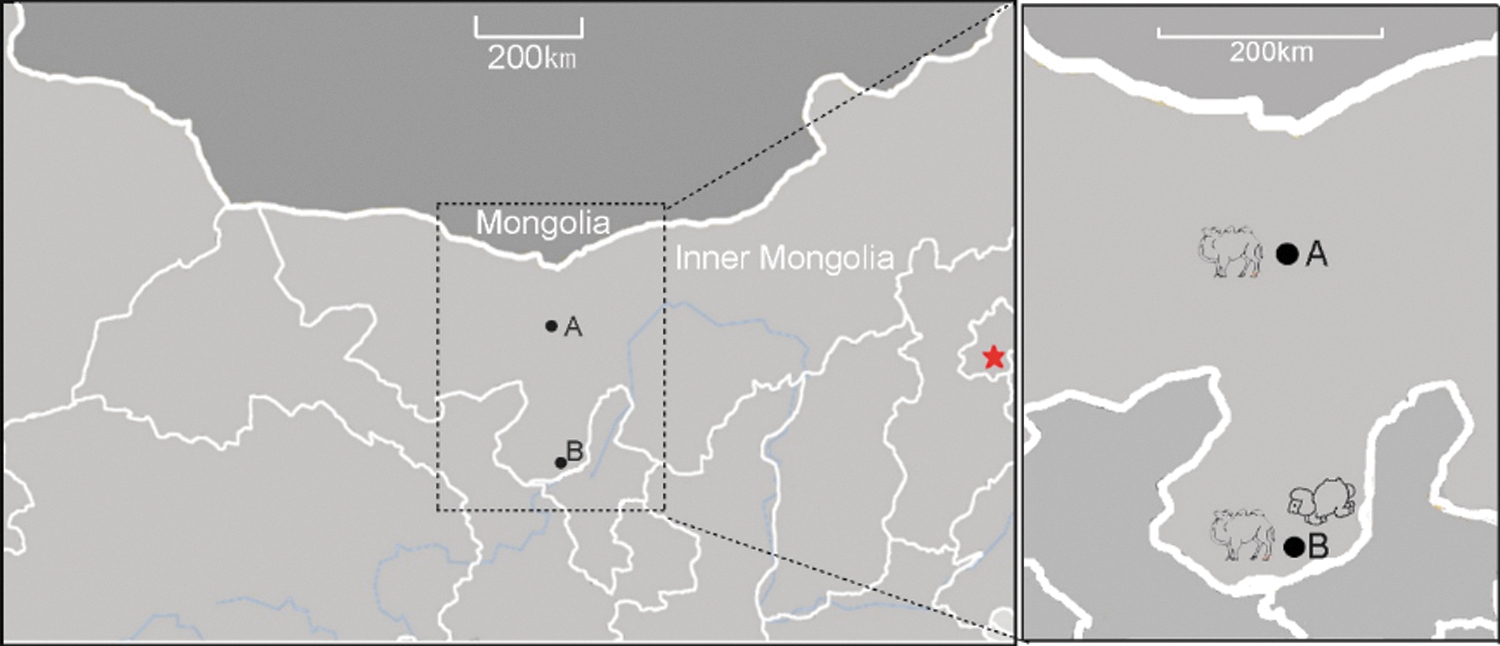
Sampling location of ticks' host sera in Inner Mongolia Province, China.
In brief, to obtain the CCHFV sequence information, the ticks grouped on sampling site, host and tick species, were pooled and homogenized in a blender (Jingxin, Shanghai, China) with Dulbecco's modified Eagle's medium (DMEM; Gibco, Carlsbad, CA) at 4°C. The viral RNAs were extracted, reverse transcribed into complementary DNA (cDNA), and then synthesized double-stranded cDNA, and employed a sequence-independent single primer amplification to get sufficient viral nucleic acid using an established method in our laboratory (He et al. 2013). The mentioned qualified PCR products of the tick groups were pooled together and then subjected to Solexa sequencing in one lane by the Beijing Genome Institute (BGI, Shenzhen, China).
Amplification S fragment of CCHFV
Based on the results of viral metagenomic analysis, the tick groups containing CCHFV viral nucleic acid were ground by a glass grinder, virus nucleic acids were extracted using an RNeasy mini kit (Qiagen). Primers were designed according to the open reading frame (ORF) sequence of S fragment of CCHFV published in GenBank (KX013455). Site of restriction enzymes BamHI and XhoI were incorporated at ends of the forward and reverse primers, respectively. The sequence of forward primer was 5′-GCGGATCCATGGARAAYAARATCGAGGTKAR-3′, and that of the reverse primer sequence was GCCTCGAGYYARATRATRTTRGCACTDGTRGC. The S fragment ORF was amplified by PCR using Taq PCR MasterMix (KT201; Tiangen Biotech Co. Ltd.). The PCR parameters is as follows: 94°C for 3 min, then 94°C for 30 s, 60°C for 30 s, and 72°C for 90 s for 35 cycles, with final 72°C for 7 min followed at 4°C for 10 min. At the end of the PCR, 1% agarose gel electrophoresis was performed to analyze the PCR products. The PCR products with expected sizes were sequenced for confirmation at Comate Bioscience Co. Ltd. (Changchun, China).
Construction of NP prokaryotic expression vector
The ORF of S fragment of CCHFV was amplified by high-fidelity enzyme PrimeSTAR HS DNA Polymerase (R010A; TaKaRa). The purified PCR product was ligated to pLB vector (Tiangen Biotech Co. Ltd.). The plasmid pLB-NP was transferred into Escherichia coli DH5a and selected on an LB medium plate (1% ampicillin resistance). After 12 h, single colonies were picked and further identified. The identified pLB-NP was used for isolation of NP fragment by BamHI (R0136S; NEB) and XhoI (R0146S; NEB) enzyme double digestion. Meanwhile, prokaryotic expression vector backbone PET-28A was also modified with BamHI and XhoI. The digestion reaction system was as follows: 3 μL of 10 × K Buffer (NEB), 1 μL of BamHI (NEB), 1 μL of XhoI (NEB), 1 μg of plasmid DNA, and ddH2O added to a final volume of 30 μL. The mentioned solution was mixed and reacted in a water bath at 37°C for 3 h. The modified NP DNA fragment and restricted enzyme-modified PET-28a backbone were ligated to generate a recombinant plasmid pET-28a-NP. The ligation reaction system was as follows: 2 μL of 10 × buffer (NEB), 1 μL of T4 DNA ligase (NEB), 1 μg of NP DNA fragment, 15 ng of pET-28a backbone, and ddH2O added to a final volume of 20 μL. The mentioned solution was mixed and reacted at 16°C for 12 h. The resultant plasmid (pET-28a-NP) was then transformed into BL-21 cells for expression of His-tagged protein after the recombinant plasmid was confirmed by sequencing (Comate Bioscience Company).
Expression and purification of NP protein
The bacterial BL-21 containing recombinant plasmid pET-28a-NP was added to 200 mL culture medium with 170 rpm shake for ∼3 h. When the optical density 600 nm reached 0.2, IPTG (isopropyl β-
Identification of E. coli-produced NP
The immunogenicity and integrity of the purified His-tagged CCHFV NP were examined by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and immunoblotting assay against antibody to His tag. Mouse Anti-6X His tag® antibody (Abcam) was diluted 1/1000 as a primary antibody, IRDye® 680 donkey antimouse IgG (LI-COR) was diluted 1/1000 as a second antibody, and 20 μg of total protein was loaded in each lane. After integrity and immunogenicity of the NP protein were confirmed, the purified protein was used for further study.
Analysis of CCHFV in serum
For the detection of reactivity to NP of animal sera in Inner Mongolia by an immunoblotting assay, the 20 μg purified NP protein was boiled in 15 μL of 1 × SDS-PAGE loading buffer for 10 min, and the proteins were resolved by SDS-PAGE under reducing conditions using 15% gels. And then the proteins were electrotransferred onto a 0.2 μm nitrocellulose membrane (Whatman). After cleaning and blocking, the nitrocellulose membrane was subsequently treated with the sera of camels or sheep (1:250 dilution in phosphate buffered saline containing 0.05% Tween 20 and 1% milk powder). Specific antigen–antibody reactions on the membrane were visualized using rabbit antisheep IgG (H&L) antibody (HRP; Abbexa), or rabbit anticamel IgG (H&L) antibody (HRP; Abbexa) at a 1:3000 dilution. The blot was performed using ECL detection reagent (GE Healthcare) according to the manufacturer's instructions. A serum sample (1:250 dilution) collected from camel or sheep with confirmed CCHFV infection was used as a positive control. A serum sample from a known healthy camel or sheep with no history of CCHFV infection was used as a negative control. And the host serum with obvious immune reaction bands were determined by positive serum with a history of exposure to CCHFV or infection.
Results
Cloning, expression, purification, and identification of NP
The NP fragment of CCHFV was amplified by PCR, and agarose gel electrophoresis is shown in Fig. 2. After sequencing, BLAST validated the sequencing results of the purified product of PCR, which confirmed the sequence of S fragment of CCHFV, with the highest amino acid homology to NP of Kazakhstan virus strain (96%). The NP gene was cloned into a pLB vector to construct the pLB-NP plasmid. The pLB-NP and pET-28a backbone plasmids were then modified with restricted enzymes for subcloning (Fig. 3). The NP gene was finally subcloned into PET-28a to construct the prokaryotic expression plasmid pET-28a-NP. The recombinant plasmid was then transformed into BL-21 cells for induction of NP protein expression.
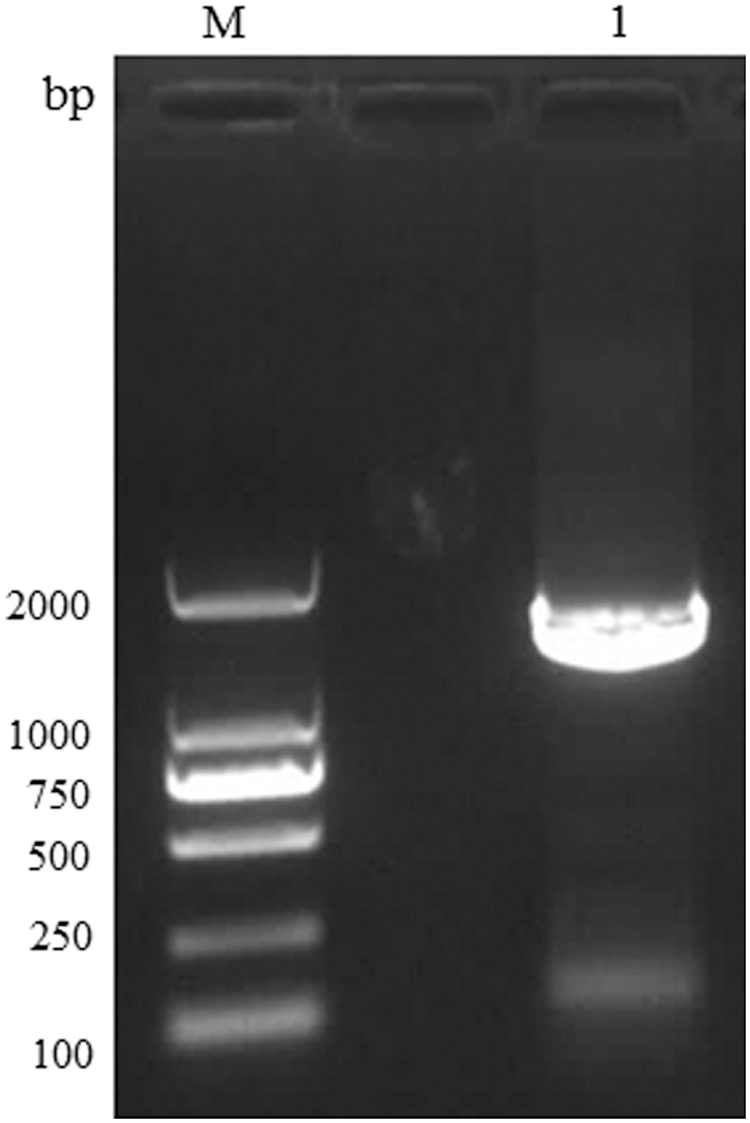
Detection of CCHFV-NP gene amplification by agarose gel electrophoresis. M, DL2000 marker. Lane 1, PCR amplification of NP fragment. CCHFV, Crimean-Congo hemorrhagic fever virus; NP, nucleocapsid protein.
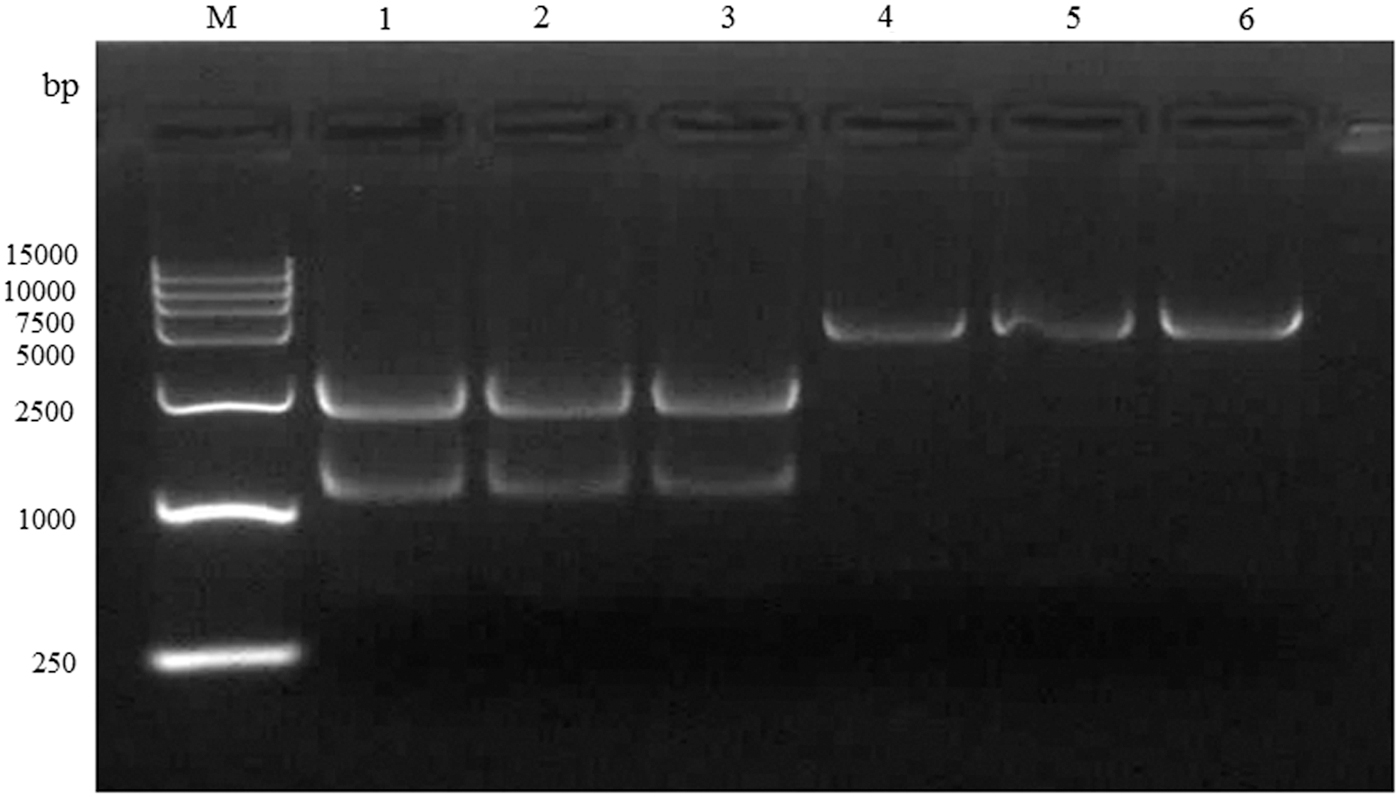
Detection of plasmid pET-28a and pLB-NP double digestion with BamHI and XhoI by agarose gel electrophoresis. M, DL15000 marker. Lanes 1–3, pLB-NP vector digestion product. Lanes 4–6, pET-28(a) digested product.
The expected molecular weight of the NP protein is 54 kDa, and majority of protein of interest was expressed in a form of inclusion bodies in bacterial cells. The recombinant protein was preliminarily isolated from inclusion bodies and then purified using nickel column affinity chromatography. The purity and integrity of protein were accessed by SDS-PAGE (Fig. 4) Supplementary Table S1. The high purity of NP recombinant protein was then resolved in SDS-PAGE before being transferred to polyvinylidene fluoride, and the anti-His mouse monoclonal antibody was used as the primary antibody to detect the specificity of the protein of interest. Immunoblots showed a targeted band with expected molecular weight, indicating the His-tagged recombinant protein (Fig. 5).
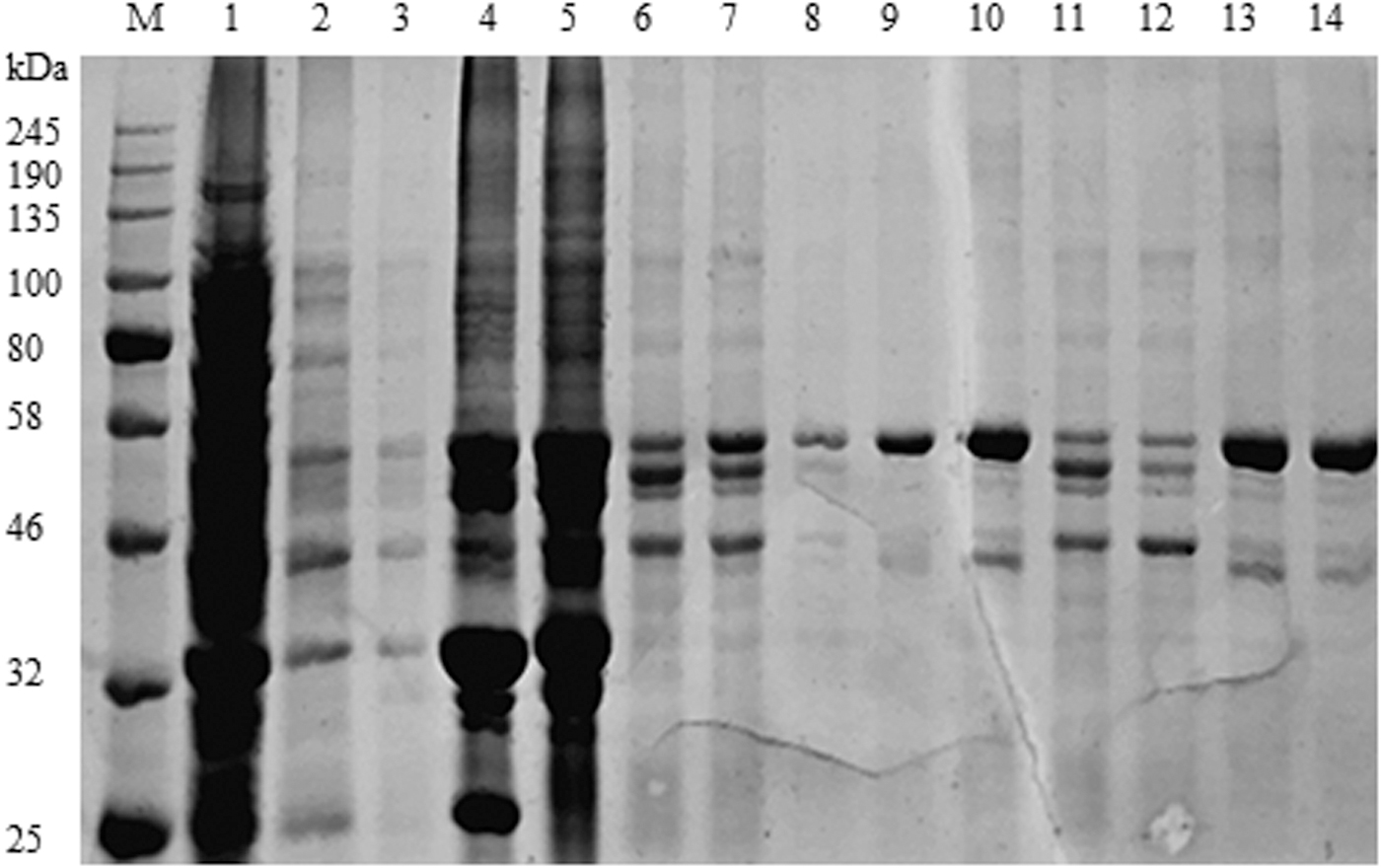
The Escherichia coli-produced NP protein of SDS-PAGE identification. M, color prestained protein standard, broad range (11–245 kDa; NEB). Lane 1, the supernatant after ultrasonic lysis. Lane 2, the supernatant washed after washing liquid I (pH 8.0). Lane 3, the supernatant washed after washing liquid II (pH 8.0). Lane 4, adding buffer B (pH 8.0) to precipitate after overnight dissolution. Lane 5, the supernatant filtered through a 0.22 nm filter of buffer B dissolution. Lane 6, the filtrate of buffer B dissolution. Lanes 7, 11, 12, the filtrate of buffer C (pH 6.3) washing solution. Lane 8, the filtrate of buffer D (pH 5.9) washing solution. Lanes 9, 10, 13, 14, the filtrate of buffer E (pH 4.5) washing solution. SDS-PAGE, sodium dodecyl sulfate–polyacrylamide gel electrophoresis.
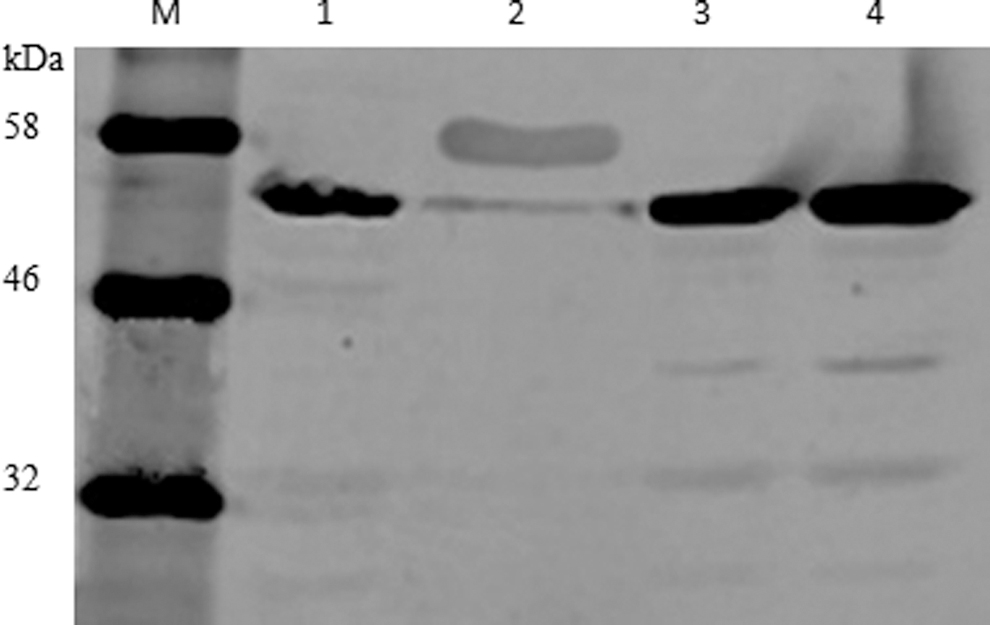
His-tagged NP recombinant protein detected by immunoblotting. M, color prestained protein standard, broad range (11–245 kDa; NEB). Lane 1, the E. coli-produced NP protein not induced. Lane 2, positive control. His-probe antibody (H-3; Santa). Lane 3, the E. coli-produced NP protein induced with 1 mM IPTG for 6 h. Lane 4, the E. coli-produced NP protein induced with 1 mM IPTG for 8 h. IPTG, isopropyl β-
Detection of antibody to CCHFV in serum
The CCHFV NP antigen is not commercially yet available. We used the E. coli-based expression system to produce the NP antigen, and then the expressed NP antigen was used to detect the antibody, which may have occurred in host serum during virus infection. To get better results, we tested the serum dilution ratio for sheep and camels in the preliminary experiments. The appropriate dilution ratio is 1:100–1:300 for the sheep serum and 1:200–1:300 for the camel serum (data not shown).
We used purified NP protein as antigen to probe antibody to CCHFV in sera of tick vector animals (camels and sheep) by Western blotting assay (Supplementary Fig. S1.). Results showed that the positive rate of camel sera at the junction of the left and right borders of Inner Mongolia was 58.3% (35/60); the seroprevalence rate of camel in Wulan Huduge of Wulan, Inner Mongolia, was 54% (12/22). The seroprevalence rate of Huduge sheep was 6.7% (2/30). The total positive rate of antibody to CCHFV in camels was 57.3 (47/82), which was significantly higher than that in sheep in Inner Mongolia region of China. This discrepancy may be due to different range of activities and behavior between camels and sheep. Camels and sheep in Inner Mongolia are stocked, but the range of activity of camels was significantly broader than that of sheep, and the outdoor residence time of camels was also significantly longer than that of sheep. This might cause the camel to be exposed to tick bites much longer than sheep. As a result, the reaction rate of antibody to CCHFV in camels was, therefore, significantly higher than that of sheep in this investigation. In addition, differences in breeding time could also lead to differences in the seroprevalence of CCHFV in animals.
Discussion
CCHFV is widely distributed and can be transmitted in a variety of vertebrates and humans in various forms. Serological detection of animals in particular regions has become an important tool for monitoring the prevalence and spread of CCHFV. To investigate the prevalence of CCHFV in animals in Inner Mongolia, Western blotting assay was performed using sera of camels and sheep collected from two regions of Inner Mongolia Province of China.
Seroprevalence may vary due to sample size, seasonality, and diversity of sampling locations. After comparing animal serological testing techniques for other zoonotic viral hemorrhagic fever, nine techniques have been reported to successfully detect CCHFV infection by serological diagnosis (Swanepoel et al. 1986). Among them, the enzyme-linked immunosorbent assay (ELISA) was generally considered to be the preferred method for CCHFV serological investigations because it could detect CCHFV antibodies in small quantities of infectious or inactivated antigens. It was more sensitive, specific, and rapid than complement-fixation test (CF), fluorescent antibody method (FA), reversed passive hemagglutination inhibition test (RPHI), and agar gel diffusion precipitation (Donets et al. 1982).
There were few studies investigating the prevalence of CCHFV in camel sera. In this study, we found the positive rate of camel sera at Alxa Youqi County of Inner Mongolia was 58.3% (35/60); the seroprevalence of camel in Ulan Hudu Gacha Village is 54% (12/22). Compared with the positive rate of CCHFV antibody in central Sudan (21.3%) (Suliman et al. 2017) reported in 2017 and the positive rate in northern Iran (5.3%) (Champour et al. 2016), the risk of CCHFV infection of camels in Inner Mongolia of China was higher.
In contrast, the seroprevalence rate of sheep in Ulan Hudu Gacha Village was significantly lower than that of camel, probably because the prevalence of CCHFV serology was related to the age of the host animals, as sera from adult camels were tested in this report. Sheep usually go out to graze during the day and return to the sheepfold to rest at night. Compared with sheep, camels are often freely farmed outdoor for a long time in a large area, and take rest in the wild desert grassland at night; therefore, camels might be exposed to CCHFV-infected ticks for much longer time relative to sheep. In addition, animal seroprevalence might be related to the range of activities and behaviors of camels and sheep. Camels and sheep in Inner Mongolia are reared in pens, but the range of activity of camels was significantly broader than that of sheep, and the outdoor retention time was also significantly longer than that of sheep, which might also be a cause of the significantly higher seroreactivity in camels than in sheep.
The seroprevalence rate of sheep was only 6.7% (2/30) in Ulan Hudu Gacha Village of Inner Mongolia, which was similar to the seroprevalence rate of 6.3% (17/270) in sheep reported in Egypt in 2008 (Mohamed et al. 2008), but it was significantly lower relative to camels, and the seroprevalence of sheep (85.7%) and goats (66.6%) reported in northern Turkey (Albayrak et al. 2012). Interestingly, in a serological test of Italian sheep, no antibody was detected using a recombinant protein antigen, but the presence of CCHFV was detected in ticks from sheep (De Liberato et al. 2018). This indicated that there was a large variation in the positive rate of antibody to CCHFV in sheep sera, which might be related to the difference of detection method and antigen used.
Our results presented in this report indicate that CCHFV is prevalent in the Inner Mongolia Province in China. As early as 2012, studies have shown that the ticks carrying the CCHFV in the border areas of Mongolia were very similar to the strains found in Kazakhstan and China, which belong to CCHFV type 2 in Asia (Voorhees et al. 2018). Therefore, it was not surprising that CCHFV antibodies were detected in the sera of camels and sheep bitten by locusts in this area. In addition, because no commercial IgG antibody assay kits was available for detecting CCHFV infection in animals, we established immunological methods for CCHFV IgG serological detection in our laboratory. But there are maybe some defects in sensitivity and high-throughput screening host sera compared with ELISA. We will improve the CCHFV sera antibody detection methods and techniques for camels and sheep in future.
The livestock industry in Inner Mongolia has been well developed, and the number of cattle, sheep, horses, and camels bred in the area is quite large. The summer from March to May is the main outbreak time of ticks, which may lead to an increased risk of CCHFV infection among tourists and residences and animals in the area. Therefore, further studies monitoring livestock, birds, and ticks, as well as high-risk populations in the area, will allow us determine the dynamic spread of CCHFV in Inner Mongolia and Mongolia and surrounding areas.
Footnotes
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This study was supported by grants from the National Natural Science Foundation of China (grant no. 31760736) and China postdoctocral science foundation (grant no. 2016T91006).
Supplementary Material
Supplementary Figure S1
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.