
Abstract
There is a considerable increase in vector-borne zoonotic diseases around the world, including Turkey, such as Crimean–Congo hemorrhagic fever (CCHF), tick borne encephalitis (TBE), Rift Valley fever (RVF), and West Nile fever (WNF), causing disease and death in humans and animals and significant economical losses. Hence, the aim of this study was to investigate the presence of CCHF virus (CCHFV) and TBE virus (TBEV) in ticks and RVF virus (RVFV) and WNF virus (WNV) in mosquitos, as well as in sheep and cattle, in the Thrace district of the Marmara region, which borders Bulgaria and Greece. Buffy-coat samples from 86 cattle and 81 sheep, as well as 563 ticks and 7390 mosquitos, were collected and examined by quantitative real-time RT-PCR for the presence of CCHFV, TBEV, RVFV, and WNV. All buffy-coat samples from cattle and sheep were negative for these viruses. Similarly, all tick samples were negative for CCHFV-RNA and TBEV-RNA. Among 245 pools representing 7390 mosquitos, only 1 pool sample was found to be positive for WNV-RNA and was confirmed by sequencing. Phylogenetic analysis revealed that it was WNV lineage-2. No RVFV-RNA was detected in the 245 mosquito pools. In conclusion, results of this study indicate that CCHFV, TBEV, and RVFV are not present in livestock and respective vectors in the Thrace district of Marmara region of Turkey, whereas WNV-RNA was found in mosquitos from this region.
Introduction
Crimean–Congo hemorrhagic fever (CCHF), tick borne encephalitis (TBE), West Nile fever (WNF), and Rift Valley fever (RVF) are zoonotic vector-borne diseases affecting people and animals worldwide. The etiological agents of CCHF, TBE, WNF, and RVF are CCHF virus (CCHFV), TBE virus (TBEV), West Nile virus (WNV), and RVF virus (RVFV), respectively. The TBEV and WNV are enveloped, single-stranded, positive-sense RNA viruses belonging to the genus Flavivirus within the family Flaviviridae (Klaus et al. 2010, Süss 2011, Chancey et al. 2015). The CCHFV and RVFV belong to the Bunyavirales order, to the families Nairoviridae and Pheniuviridae in the genus Orthonairovirus and Phlebovirus, respectively. Both viruses are enveloped and possess single stranded, negative-sense, segmented RNA genomes (Süss 2011, Hawman and Feldmann 2018).
CCHFV and TBEV are mainly transmitted by Ixodes spp. (Süss 2011, Hawman and Feldmann 2018). Humans are infected through the bite of infected ticks and direct contact with tissues or body fluids from viremic animals and humans (Klaus et al. 2010, Bogovič and Strle 2015). A short duration of viremia occurs in ruminants after exposure to CCHFV and TBEV (Swanepoel et al. 1987, Mansfield et al. 2016). RVFV and WNV are mainly transmitted by Culex mosquitos and sometimes by blood transfusion or organ transplantation and by direct contact with tissues or body fluids of viremic animals and humans in laboratory and field settings and abattoirs (Chancey et al. 2015). A short term viremia in RVFV and WNV infected ruminants was reported (Barnard and Voges 1986, Faburay et al. 2016).
Birds are the main reservoirs for WNV, and other animals like wild ruminants, rodents, and bats seem to play an important role in the ecology of WNV, as well as RVFV; however, the role of the rodents, bats, and ruminants in virus transmission is still unclear. Clinical signs can be observed in ruminants after RVFV infection, especially in sheep, but not with WNV infections. No chronic infections are seen in ruminants infected with either of these viruses (Shepherd et al. 1987, Evans et al. 2008, Chancey et al. 2015, Romeo et al. 2018).
After transmission of CCHFV, TBEV, RVFV, and WNV, a wide variety of symptoms develop in humans ranging from fatique to fever, nausea, vomiting, abdominal pain, back pain, hemorrhages (CCHF), and neurological signs (TBEV, RVF, and WNF) (Madani et al. 2003, Pepin et al. 2010, Süss 2011, Bogovič and Strle, 2015, Chancey et al. 2015, Papa et al. 2017b). RVFV is characterized by mass abortion and newborn deaths in ruminants (Moussa et al. 1986). WNV causes neurological symptoms mainly in horses. However, signs of CCHF and TBE are not well characterized in animals at present (Süss 2011, Hawman and Feldmann 2018).
It is rather difficult to diagnose CCHF, TBEV, RVF, and WNF infections in humans and animals based on clinical signs, since the symptoms are often unspecific. A history of vector bite/contact might support a suspect diagnosis. In any case, laboratory confirmations are needed and generally made by virus isolation, as well as serological and molecular tests (Saksida et al. 2005, Klaus et al. 2010). Virus isolation is the gold standard for the diagnosis of these infections. However, for the routine diagnosis, molecular detection and serology are mainly used. Serological tests are mainly based on indirect enzyme-linked immunosorbent assay (ELISA) systems; they detect virus-specific antibodies, but do not determine the presence of virus. It is important to note that a rather short viremic period is seen with CCHFV, TBEV, WNV, and RVFV infections of humans and animals.
Although serology may be preferred to indirectly determine the presence of ruminant infections with these viruses, our goal for this study was to obtain direct evidence of infection. The detection of viral nucleic acids and subsequent sequencing of the respective amplicons would allow determination of the genetic lineage of the respective viruses present in Turkish ruminants. This is not possible with serology. In addition, there is usually no antibody response at early time points of acute infections during viremia.Therefore, molecular tests such as real-time RT-PCR (RT-qPCR) is the preferred method to detect viral RNA in blood, sera, or tissues.
Specificity and sensitivity of virus isolation, RT-qPCR, and indirect ELISAs vary. In general, molecular tests are very sensitive, specific, and rapid, whereas virus isolation is less sensitive and time and labor intensive. The sensitivity and specificity of ELISAs often depend on the immunogenicity of the target antigen/s used for coating the ELISA plates (Barnard and Voges 1986, Holzman 2003, Saksida et al. 2005, Klaus et al. 2010, Faburay et al. 2016, Mansfield et al. 2016).
Since these diseases are vector borne, endemic regions and countries need to perform active monitoring for the presence of CCHFV and TBEV in ticks and RVFV and WNV in mosquitos (Evans et al. 2008, Bente et al. 2013, Bogovic and Strle 2015, Chancey et al. 2015). Therefore, the aim of this study was to investigate the presence of CCHFV and TBEV in ticks and RVFV and WNV in mosquitos, as well as in sheep and cattle, in the Thrace district of the Marmara region, which borders the European Union.
Materials and Methods
Study area and sampling
In this study, mosquitos, ticks, cattle, and sheep were sampled in the Thrace district of the Marmara region in Turkey, bordering Greece and Bulgaria. To collect mosquito samples, night traps and gravid traps were used. Selection criteria to set up mosquito traps were the geographic area and the presence of animal farms in the area. Ticks were collected from cattle and sheep, and the blood from those animals was also collected in parallel.
Collection of mosquitos
Gravid (The CDC Gravid Trap Model 1712, John W. Hock Co., USA) and light traps (CDC miniature light trap-model 512, John W. Hock Co., USA) were used to collect mosquitos. Traps were set up in the evening in 14 different localities within the Thrace district and were collected the next morning. The sampled mosquitos were exposed to triethylamine (Sigma-471283, USA) to anesthetize them and then transferred to the laboratory in dry ice. They were then specified under a light microscope. In total, 7390 mosquitos were collected (Table 1), and these mosquitos were identified on the genus level using the morphological keys by Samanidou-Voyadjoglou and Harbach 2001. Finally, a total of 245 pools were formed.
The Localities and Species of Mosquitos Collected in This Study
Collection of ticks and blood samples
Ticks were collected by hand from 86 cattle and 81 sheep, which were infested by ticks on their skin. At least five ticks from moderately infested animals and at most 10 ticks from different skin regions of heavily infested animals were collected. Blood tubes containing ethylenediaminetetraacetic acid (EDTA) were used to draw blood from these animals. Ticks and blood samples were transferred to the laboratory using a cold chain. In total, 563 ticks were collected (Table 2), and 140 pools were formed based on genus identification made by observation on light microscopy as described previously (Keirans and Litwak 1989). Ticks from different animals were never mixed in pooling.
Species and Localities of Ticks Collected in This Study
RNA extraction
RNA extraction from mosquitos and ticks was performed using the RNeasy Mini Kit (Qiagen, Germany) as described by the manufacturer. Nucleid acids were purified from pools of mosquitoes (10–30 specimens) and ticks (8–10 specimens) from the same animal. Ticks and mosquitoes were cut into small pieces before RNA extraction was performed. Fragments were then transferred to Eppendorf tubes, and 0.3 g of 1 mm still beads and 800 μL lysis buffer (RLT) (with beta mercaptoethanol) were added into each tube. The tubes were then placed into a tissue homogenizer (Ribolyser, Hybaid) and homogenized for 40 s with 4 m/s speed. The homogenates were then centrifuged at 14,000 rpm at 4°C for 2 min. The supernatant was removed, and an equal volume of 96–100% ethanol was added. The RNA extraction continued as decribed by the manufacturer. The eluate was collected, and the RNA concentration was measured using a Nanodrop. The extracted RNA was either used directly for analysis or stored at −80°C until used.
For the extraction of viral RNA from buffy-coat samples from animal blood, QIAamp Viral RNA Mini Kit (Qiagen, Germany) was used. Extraction procedures were performed as described by the manufacturer. The amount of RNA in the extracted material was measured using a NanoDrop spectrophotometer (NanoDrop 1000c; Thermo Scientific).
Reverse transcription
Reverse transcription (RT) was performed as described previously (Yilmaz et al. 2017). In brief, RT was performed in two steps. For the first step, 9 μL of RNA template (∼400 ng) was mixed with 1 μL (0.4 μg) of random hexamers (Promega). For the second step, a total volume of 20 μL reaction mixture was prepared that consisted of 10 μL RNA/primer mixture from the first step, 4 μL 5 × RT buffer, 2.4 μL 25 mM MgCl2, 1 μL 10 mM dNTPs, 1.6 μL nuclease-free water, and 1 μL reverse transcriptase (ImProm II; Promega). After completion of the RT reaction at 55°C for 30 min (Yilmaz et al. 2017), 30 μL of nuclease-free water was added to each cDNA sample.
Quantitative real-time RT-PCR for detection of CCHFV-RNA, TBEV-RNA, WNV-RNA, and RVFV-RNA
TaqMan real time RT-PCR assays were performed to analyze all vector samples and the blood of animals. The primers, probes, reaction mixtures, and amplification conditions for the TaqMan real-time PCR to detect CCHFV-RNA (Atkinson et al. 2012), TBEV-RNA (Schwaiger and Cassinotti 2003), WNV-RNA (Linke et al. 2007), and RVFV-RNA (Bird et al. 2007) in mosquitos, ticks, and buffy-coat samples are described in Table 3. Quantitative real-time RT-PCRs were performed in a 25 μL PCR always, including positive and negative controls, which were provided by: Dr. Robert S. Lanciotti, Centers for Disease Control and Prevention, USA; Dr. Christine Klaus, Friedrich-Loeffler-Institute, Germany; and Professor Juergen Richt, Kansas State University, Kansas, USA. Amplification was performed in a Stratagene MX3005P real-time PCR instrument.
Primers And Probes Used For the Real Time RT-PCR Assays in This Study
CCHFV, Crimean–Congo hemorrhagic fever virus; RVFV, Rift Valley fever virus; TBEV, tick borne encephalitis virus; WNFV, West Nile fever virus.
RT-PCR for sequencing
Samples found to be positive by RT-qPCR were also analyzed by conventional RT-PCR. For this purpose, conventional RT-PCR was performed for the amplification of WNV-RNA using the primers previously published by Lanciotti et al. (2000). Briefly, an optimized 50 μL RT-PCR mixture consisted of 7 μL of RNA and 20 pmol of each primer, 25 μL Platinum™ SuperFi™ PCR Master Mix (12358010; Invitrogen), and negative and positive controls (provided by Dr. Robert S. Lanciotti, Centers for Disease Control and Prevention, USA). Amplification was performed in a Stratagene MX3005P PCR machine by following the instructions included in the Platinum SuperFi PCR Master Mix Kit. Products were analyzed by gel electrophoresis using a 1.5% agarose gel.
Phylogenetic analysis
Alignments of the partial sequences of the nucleocapsid and premembrane protein genes of WNV were made using the MEGA-7 software. Phylogenetic analyses were carried out using the criterion of neighbor-joining trees based on genetic distance model by Tamura et al. (2004). The reference sequences from the GenBank for WNV classification were used to reconstruct the topology of the partial sequences of the nucleocapsid and premembrane protein genes of WNV in this study. The partial sequences of the nucleocapsid and premembrane protein genes of WNV obtained in this study were submitted to GenBank (MN816439).
Results
Animals, identification of tick, and mosquito samples
No clinical signs of disease were observed in the sheep and cattle from which ticks were collected except that emaciation was observed in heavily tick-infested animals. Among the 7390 mosquitos collected in this study, 4085 were identified as Culex, 3153 as Anopheles, 99 as Aedes, and 53 as Culiseta (Fig. 1A and Table 1). Among the 563 ticks collected from 167 animals, 506 were identified as Boophilus-Rhipicephalus, 29 as Ixodes, and 28 as Hyalomma (Fig. 1B and Table 2).
Mosquitos and ticks collected in this study.
Quantitative real time RT-PCR
Among the 245 pools representing 7390 mosquitos, only 1 pool containing 18 mosquitos (Culex) gave a positive amplification signal using the WNV-specific real time RT-PCR. The C T value of this sample was 34, while the C T value of positive control was 24. No C T value was obtained with the negative control (Fig. 2). The mosquito pool was collected from the Igneada site near the Bulgarian border. The other 244 mosquito pools were negative for WNV-RNA, and all 245 pools were negative for RVFV-RNA.
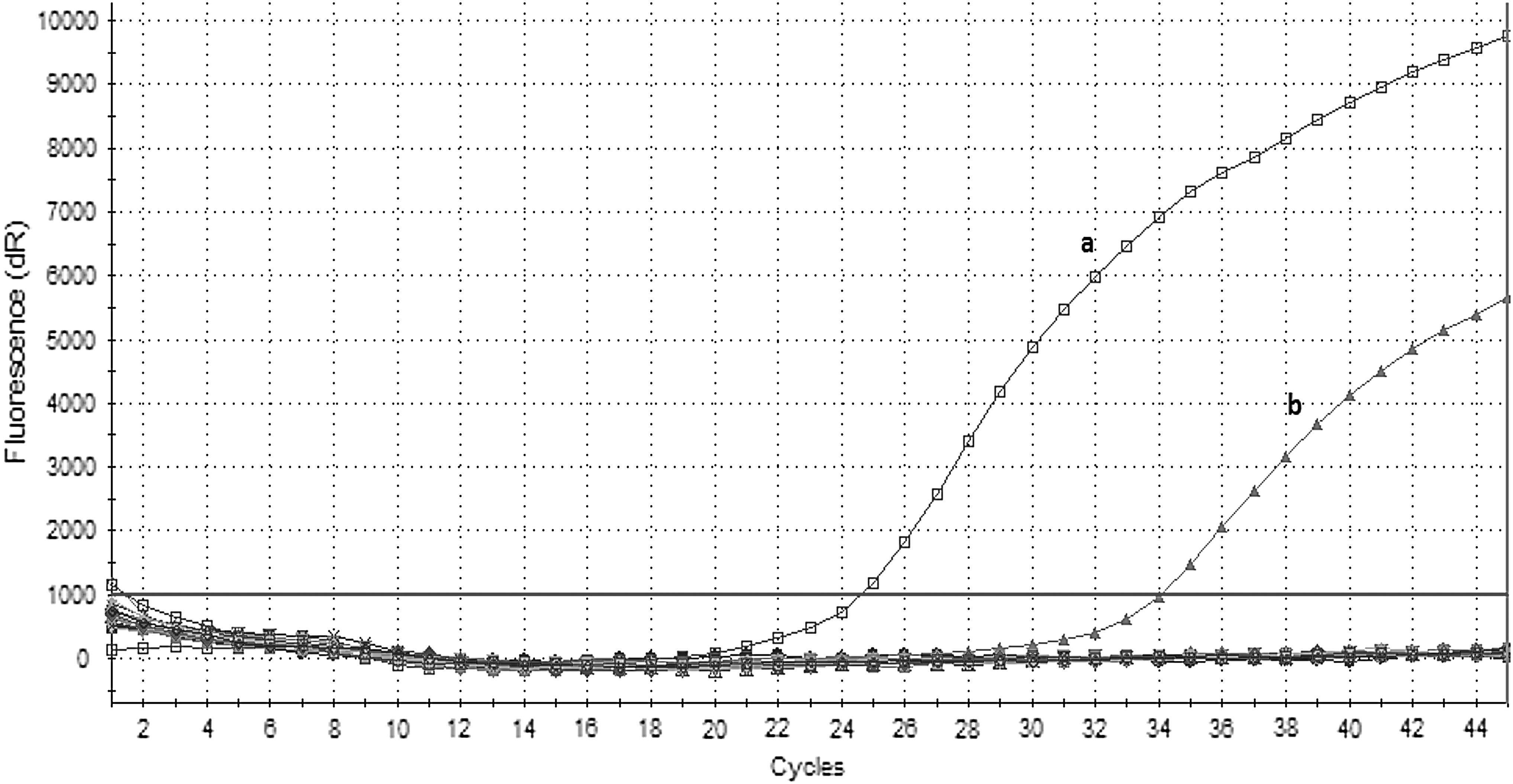
Quantitative real time RT-PCR of WNV for mosquito samples.
All 140 tick pools were found to be negative for CCHFV-RNA and TBEV-RNA by real-time RT-PCR. In addition, no CCHFV-RNA, TBEV-RNA, RVFV-RNA, and WNV-RNA were detected in buffy-coat samples taken from the cattle and sheep. Positive controls always gave positive amplification signals in the respective real time RT-PCR assays, but no signal was seen in negative control samples.
RT-PCR and sequencing
The mosquito pool 242, which was found to be positive for WNV-RNA by real time RT-qPCR, was subjected to conventional RT-PCR for subsequent sequencing. A 408-bp PCR product was detected on gel electrophoresis with the positive mosquito pool and the positive control, but not with negative control. The amplified PCR product from the positive mosquito pool was sequenced, and phylogenetic analysis was performed as described in Materials and Methods section.
Phylogenetic analysis
The phylogenetic analysis revealed that the partial sequences of the nucleocapsid and premembrane protein genes of WNV were closely related to WNV lineage-2 (Fig. 3), similar to WNV isolates that were reported from Italy (KP789960; 97.8% homology), Czech Republic (KM 203863; 98.3% homology), Serbia (KT 757318; 98.5% homology), Austria (KM 659876; 98.2% homology), and Greece (HQ 537483; 98.2% homology). However, the Turkish WNV lineage-2 (MN 816439) sequence was slightly different from the WNV lineage-2 viruses detected in Romania (KJ 934710; 95.1% homology), Italy (KT 207791; 95.1% homology), and South Africa (JN 393308; 96.2% homology) and, therefore, placed in a different cluster (Fig. 3).

Maximum likelihood phylogenetic tree constructed based on partial sequences of the nucleocapsid and premembrane protein genes of WNV. The classification has been performed using the reference sequences submitted to GenBank. Circular dot indicates the WNV detected in this study.
Discussion
Vector-borne diseases are affecting animals and people globally and can cause severe health problems and significant economic losses. Monitoring the vectors transmitting vector-borne viruses is an important biosurveillance tool to understand which viruses are circulating in a specific geographic region of concern (Pepin et al. 2010, Süss 2011, Bente et al. 2013, Engler et al. 2013, Himeidan et al. 2014, Petterson et al. 2014). Ticks and mosquitos are major vectors for viruses affecting both animals and humans. Therefore, in this study, ticks were analyzed for the presence of TBEV and CCHFV and mosquitoes for the presence of WNV and RVFV. In parallel, blood was also collected from cattle, sheep, and goats to investigate the presence of RNA for TBEV, CCHFV, WNV, and RVFV. For this, real time RT-PCR was the method of choice in both blood and pooled vector samples as previously decribed by others (Schwaiger and Cassinotti 2003, Bird et al. 2007, Linke et al. 2007, Atkinson et al. 2012).
TBE is endemic in Europe, Syberia, Eastern Russia, Northern China, and Japan. About 10,000–15,000 human TBE cases have been reported in 2018 in Europe and Asia. TBE has been diagnosed in 27 European countries, and among them it was more frequently reported in Slovenia, Estonia, Lituania, and Latvia (Petterson et al. 2014, Bogovic and Strle 2015). Antibodies to TBEV in humans have been reported in Turkey (Gunes et al. 2010, Ergunay et al. 2011, Yilmaz et al. 2019). However, till now, neither antibodies to TBEV nor TBEV-specific RNA has been detected in animals in Turkey. The aim of the present study was to detect acute infections when animals exhibit viremia, and all blood samples taken from cattle and sheep were found to be negative for TBEV-RNA by real time RT-PCR. This may be because of the short period of viremia associated with TBEV infections in ruminants (Mansfield et al. 2016).
TBEV is transmitted by Ixodes ticks. Ixodes ricinus in Europe, Ixodes persulcatus in Eastern Europe, Russia, and Far East Asia, and Ixodes ovatus in Japan have been found responsible to transmit TBEV to humans. TBEV-RNA was detected in 0.1–5% of the ticks in central Europe, up to 40% of the ticks in Syberia, and in 0.47–0.54% of the ticks in Slovenia (Pettersson et al. 2014, Bogovic and Strle 2015). The detection rate of TBEV was about 8.6% in adult ticks in Norway (Soleng et al. 2018). In Sweden, prevalence in ticks in the endemic areas ranged from 0.1% to 0.42%, which is comparable with other more established TBE endemic regions in Europe (Brinkley et al. 2008). In Russia, TBEV RNA was detected in up to 25% of I. persulcatus ticks (Pukhovskaya et al. 2018).
In Denmark, among 9321 I. ricinus, 2 pools of 34 tick pools were tested positive for TBEV-RNA resulting in a local estimated prevalence of 0.6% (Andersen et al. 2019). In Hungary, 2731 ticks (I. ricinus, Haemaphysalis concinna, Dermacentor marginatus) were analyzed for the presence of TBEV-RNA; TBEV prevalence in ticks was 0.08% for unfed nymphs and 0.78% for feeding larvae (Pintér et al. 2013). In Germany, the average TBEV prevalence in I. ricinus (nymphs, adults) ticks in Bavarian risk areas was between 0.5% and 2% (Süss et al. 2006). In a recent study conducted in France and Germany, minimal infection rates in ticks of 0.17% (1/595) for Schiltach (Germany) and 0.11% (1/944) for Foret de la Robertsau (France) were reported (Bestehorn et al. 2018).
In Bulgaria, a neighboring country of Turkey, 1542 ticks were analyzed, and 12 novel phleboviral sequences were found. The sequences formed five distinct groups and clustered with other tick-borne phleboviruses recently identified in Europe (Ohlendorf et al. 2019). In the present study, no TBEV-RNA was detected in 140 pools representing 563 ticks collected from cattle and sheep. Till now, there has been no study to investigate TBEV in ticks in Turkey. The ticks collected in this study were mainly ticks belonging to the Boophilus spp. (Rhipicephalus spp.) since they are more prevalent in the Thrace district than Ixodes spp., which represent the main reservoir of TBEV. Importantly, infection of Boophilus spp. ticks with TBEV was previously reported in GenBank Accession Number: JN936368.
We conclude from our study that genomic markers of TBEV are not present in ticks belonging to the Boophilus spp. in the Thrace district of Turkey. Although the results of this study indicate the absence of TBEV-RNA in ticks collected from the Thrace district of Marmara region in Turkey, further studies in different regions of Turkey are necessary.
CCHF was first reported in Russia and later in Bulgaria and Africa. It is presently found in Asia, Africa, and South Western Europe. Turkey had the highest prevalence in the last decade (Vatansever et al. 2007, Chinikar et al. 2010, Bente et al. 2013). A short duration of viremia occurs in ruminants after exposure to CCHFV, and the infection presents without clinical signs. In South Africa, antibodies to CCHFV were detected in up to 30% ruminants (Swanepoel et al. 1987). In the present study, no CCHFV-RNA was detected in buffy coat samples taken from Turkish cattle and sheep. However, in a study performed in the Black Sea region of Turkey, CCHFV RNA was found in nine of 105 (8.57%) leukocyte samples obtained from ruminants, as well as antibodies to CCHFV (Albayrak et al. 2012). Antibodies to CCHFV have not been investigated in this study since our goal for the study was to detect direct evidence of infection.
CCHFV is transmitted by 31 different species of ticks. Hyalomma ticks are the most common reservoir for CCHFV (Bente et al. 2013, Estrada-Peña and de la Fuente 2014). In Iran, among 90 tick pools, 5 Hyalomma pools were positive for CCHFV-RNA (Fakoorziba et al. 2012). In Bulgaria, CCHFV-RNA was detected in about 2% of 911 ticks (Gergova et al. 2012). In Spain, 1 of 12 Hyalomma lusitanicum pools was found to be positive for CCHFV-RNA (Estrada-Peña et al. 2010). In Greece, CCHFV was detected in 36 of 1290 (2.8%) tick pools (Papa et al. 2017a). In Albania, a total of 726 ticks (366 Hyalomma marginatum, 349 Rhipicephalus bursa, and 11 Rhipicephalus sanguineus) were collected, and 30 of 215 (13.9%) tick pools were positive for CCHFV-RNA (Papa et al. 2017a).
In Turkey, tick pools were investigated for the presence of CCHFV-RNA by various investigators. Depending on region and sample size, between 2% and 22.7% of the tick pools were found to be positive for CCHFV-RNA and the majority of the tick species positive for CCHFV-RNA belong to the Hyalomma, Dermacentor, and Rhipicephalus genera (Tonbak et al. 2006, Albayrak et al. 2012, Hekimoglu et al. 2012, Tekin et al. 2012, Yesilbag et al. 2013, Orkun et al. 2017). In a study performed in the Thrace district of Turkey, a total of 15 tick pools were collected from 9 localities. CCHFV-RNA was detected only in H. marginatum, Rhipicephalus (Boophilus) annulatus, and Rhipicephalus bursa with an overall maximum likelihood estimate of infection rates being 0.93% (95% CI = 0.35–2.05), 0.74% (95% CI = 0.24–1.78), and 1.67% (95% CI = 0.69–3.46), respectively (Gargili et al. 2011). In the present study, CCHFV-RNA was not detected in 140 tick pools collected from the same region, that is, the Thrace district of Turkey.
After first detection of WNF in Uganda, cases have been reported in South Africa, France, Israel, and Egypt between 1950 and 1980. It was then detected in Russia and Romania before it spread to other countries like the United States, Mexico, and Canada in 1999 (Daep et al. 2014, Chancey et al. 2015). WNF has been reported in humans and animals in Turkey, including cattle, sheep, horses, mules, and dogs (Ozkul et al. 2006, Ergunay et al. 2010, Hızel et al. 2010, Albayrak and Ozan 2013, Biceroglu et al. 2015, Yilmaz et al. 2019). Although WNV-RNA has been detected in horses (Ozkul et al. 2013, Toplu et al. 2015), it has not been detected in ruminants so far. Similar results were obtained in this study, which revealed the absence of WNV-RNA in buffy-coat samples taken from sheep and cattle.
It has been reported that about 45 mosquito species from 8 genera are susceptible to WNV and can transmit WNV. Among them Culex spp. and Aedes spp. are the most frequent species transmitting WNV to mammals (Chancey et al. 2015). In Italy, a total of 757,461 mosquitoes were collected and 9268 pools were formed. One hundred eighty (1.9%) of these pools were found to be positive for WNV-RNA (Calzolari et al. 2015). A prevalence rate of WNV-infected mosquitoes of 0.06% has been reported in Spain (Engler et al. 2013). Ninety-nine pools of 1303 mosquitoes were analyzed in Serbia. Two Aedes and 2 Culiseta annulata and 95 Culex pipiens pools were found to be positive for WNV-RNA (Petrić et al. 2017). In Israel, 7135 pools containing a total of 277,186 mosquitoes were investigated for the presence of WNV-RNA; WNV RNA was detected in 336 (4.71%) pools (Lustig et al. 2017).
In Iran, a neighboring country of Turkey, 45 pools representing 2143 mosquitoes were analyzed, and WNV-RNA was found in 2 pools (Bagheri et al. 2015). In the southeast region of Anatolia in Turkey, 6457 mosquitos were collected containing 56% of Culex spp. WNV-RNA was not detected in any of the samples (Ozer et al. 2007). In another study in Turkey, a total of 9261 mosquitoes were captured and divided in 232 pools representing the following mosquito species: Ochlerotatus caspius (90.9%), C. pipiens sensu lato (s.l.) (4.7%), Anopheles pseudopictus (3%), and Anopheles maculipennis s.l. (1.3%). Thirty-seven mosquito pools (15.9%) were positive in pan-flavivirus and WNV-specific assays. Viral sequences in positive pools were characterized as WNV lineage 1 clade 1a and demonstrated a 1–4% molecular divergence. WNV infection rates in Oc. caspius and C. pipiens s.l. pools were 15.6% and 36.3%, respectively (Ergunay et al. 2013).
Recently, 594 pools representing 12,711 mosquitos representing 15 different species were collected from the Thrace district of the Marmara region. Only one pool containing C. pipiens was found to be positive for the presence of WNV-RNA similar to sequences of WNV lineage-1 (Ergunay et al. 2017).
In the present study, among 245 pools representing 7390 mosquitoes, 1 mosquito pool was found to be positive for WNV-RNA. Phylogenetic analysis revealed that WNV lineage-2 (MN816439) sequences found in this study were similar to WNV lineage-2 viruses reported from Italy (KP789960), Czech republic (KM 203863), Serbia (KT 757318), Austria (KM 659876), and Greece (HQ 537483), but slightly different than WNV lineage-2 viruses detected in Romania (KJ 934710), Italy (KT 207791), and South Africa (JN 393308). This is the first study on the existence of WNV lineage-2 in mosquitoes in Thrace district, which borders European Union. In contrast, WNV lineage-1 was found in mosquitoes in the same area by others (Ergunay et al. 2017).
Although RVFV infection is widespread in humans and animals, mainly in Africa (Himeidan et al. 2014, Nanyingi et al. 2015), it is also endemic in the Middle East in Saudi Arabia and Yemen (Shoemaker et al. 2002, Madani et al. 2003, Mohamed et al. 2014, Memish et al. 2015). Given the increased cross-border travel and trade with countries within the Middle Eastern region, there is substantial risk of RVFV being introduced into Turkey.
In Saudi Arabia, 500 serum samples were taken from small ruminants, and 84% of them had antibodies to RVFV (Mohamed et al. 2014). In Tanzania, seropositivity in small ruminants was about 8.2% (Wensman et al. 2015). In South Africa, seropositivity was 7% in cattle and 1.4% in sheep. No RVFV-RNA was found in seropositive samples (Mapaco et al. 2012). In a study performed in Turkey, antibodies to RVFV were not detected in sera taken from cattle, horses, sheep, goats, and buffaloes (Albayrak and Ozan 2013). However, more recently, antibodies to RVFV were detected in camel, goitered gazelle, and Anatolian water buffaloes (Gur et al. 2017). Until now, the presence of RVFV-RNA has not been demonstrated in animals or humans in Turkey. Likewise, in the present study, no RVFV-RNA was detected in any of the samples collected from cattle and sheep.
Mosquitos of the Aedes and Culex species are mainly responsible for transmitting RVFV to mammals. In Kenya, among 3003 mosquito pools, 77 were found to be positive for RVFV-RNA and it was found in 10 different mosquito species (Himeidan et al. 2014). A total of 978 mosquitoes were collected during an outbreak in Sudan in 2007. The RVFV was detected in female, male, and larval stages of Culex and Anopheles species (Seufi and Galal 2010). In the present study, no RVFV-RNA was detected in any of the mosquito pools tested. Till now, there has been no study to investigate RVFV in mosquitos in the Thrace district of Marmara region in Turkey. Although the results of this study indicate the absence of RVFV-RNA in mosquitos collected from Thrace district of Turkey, further studies in different regions of Turkey are necessary.
Conclusions
The present study was conducted in the Thrace district of the Marmara region of Turkey to determine the presence of various arthropod-borne viruses (TBEV, CCHFV, WNV, RVFV) by real-time RT-PCR in ticks, mosquitoes, and blood samples from small ruminants and cattle. There is no evidence of TBEV, CCHFV, or RVFV in the area of Thrace, Turkey, where the study was conducted. However, WNV is present in this area, and further active surveillance should be carried out for these important zoonotic pathogens in their vector populations and ruminant hosts to protect human and animal health in this strategic region between Europe and Asia.
Footnotes
Acknowledgments
Thanks to Dr. Lanciotti for supplying positive WNV as PCR positive control and to Dr. Christine Klaus for supplying TBEV as PCR positive control.
Ethical Approval
Ethical approval was obtained from the Ethics Committee of the University of Istanbul.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was funded by The University of Istanbul (Project No. 13163).