
Abstract
Ljungan virus (LV), which belongs to the Parechovirus genus in the Picornaviridae family, was first isolated from bank voles (Myodes glareolus) in Sweden in 1998 and proposed as a zoonotic agent. To improve knowledge of the host association and geographical distribution of LV, tissues from 1685 animals belonging to multiple rodent and insectivore species from 12 European countries were screened for LV-RNA using reverse transcriptase (RT)-PCR. In addition, we investigated how the prevalence of LV-RNA in bank voles is associated with various intrinsic and extrinsic factors. We show that LV is widespread geographically, having been detected in at least one host species in nine European countries. Twelve out of 21 species screened were LV-RNA PCR positive, including, for the first time, the red vole (Myodes rutilus) and the root or tundra vole (Alexandromys formerly Microtus oeconomus), as well as in insectivores, including the bicolored white-toothed shrew (Crocidura leucodon) and the Valais shrew (Sorex antinorii). Results indicated that bank voles are the main rodent host for this virus (overall RT-PCR prevalence: 15.2%). Linear modeling of intrinsic and extrinsic factors that could impact LV prevalence showed a concave-down relationship between body mass and LV occurrence, so that subadults had the highest LV positivity, but LV in older animals was less prevalent. Also, LV prevalence was higher in autumn and lower in spring, and the amount of precipitation recorded during the 6 months preceding the trapping date was negatively correlated with the presence of the virus. Phylogenetic analysis on the 185 base pair species-specific sequence of the 5′ untranslated region identified high genetic diversity (46.5%) between 80 haplotypes, although no geographical or host-specific patterns of diversity were detected.
Introduction
Since its discovery in bank voles (Myodes glareolus) from Sweden in 1998, Ljungan virus (LV; also referred to as Parechovirus B, family Picornaviridae) has been studied for its zoonotic potential (Niklasson et al. 1998, 1999). Early studies suggested that LV-infected rodents and inoculated laboratory animals displayed human disease-like symptoms, and several publications implicated LV in human disease, including gestational disorders and those related to the central nervous system (Niklasson et al. 2007, 2009a, 2009b, Samsioe et al. 2009).
Although no specific disease symptoms have been associated with LV in humans (Fevola et al. 2016), high seroprevalences of anti-LV antibodies in Finnish residents (47.8%: Jääskeläinen et al. 2016, 33.8%: Fevola et al. 2017a) suggested that further studies on potential small mammal hosts and the genetic diversity of LV in nonhuman animals would be valuable. Although previous authors hypothesized as early as 2003 that LV may be widespread in wildlife (Johansson et al. 2003), very little is still known about its host range and geographical distribution, and previous research on potential hosts of LV has mainly focused on prevalence in a single country and/or potential host species/population.
To date, LV-RNA has been detected in wild rodents in Denmark (bank vole: Niklasson et al. 2003), Finland (bank vole: Jääskeläinen et al. 2013 and field vole Microtus agrestis: Forbes et al. 2014, Fevola et al. 2017b), Italy (bank vole and yellow-necked mouse Apodemus flavicollis: Hauffe et al. 2010 and red squirrel Sciurus vulgaris: Romeo et al. 2014), the United Kingdom (bank vole and field vole: Salisbury et al. 2014), and Sweden (bank vole, field vole, and gray-sided vole Craseomys, formerly Myodes rufocanus, Norway lemming Lemmus lemmus, and wood lemming Myopus schisticolor: Niklasson et al. 1999, 2006, Fevola et al. 2017b), as well as in voles in the United States (montane vole Microtus montanus: Johansson et al. 2003 and southern red-backed vole Myodes gapperi: Tolf et al. 2009). Reverse transcriptase (RT-)PCR prevalence has ranged from 8% to 50% per population, with the highest prevalence found in bank voles in Italy (Hauffe et al. 2010) and Sweden (Fevola et al. 2017b). As a result, this species is purported to be the main host of LV (Warvsten et al. 2017).
In this study, we used molecular screening of small mammals to investigate the presence of LV in 12 European countries, and in bank voles and other potential hosts. In addition, we investigated how the prevalence of LV in bank voles is associated with various intrinsic (e.g., sex and age) and extrinsic factors (e.g., weather and other environmental parameters).
Materials and Methods
Sample collection
As the aim of the study was to gain basic knowledge of the host and geographical distribution of LV, convenience sampling was considered appropriate; that is, liver samples from 21 small mammal species that had already been collected during various work packages of the EU FP7 project EDENext (Emerging Diseases in a Changing European Environment;
Number of Ljungan Virus-RNA Reverse Transcriptase-PCR-Positive/Total Individuals Screened (Prevalence %) in 15 Rodent Species and Representatives of 2 Insectivore Genera from 12 European Countries
20 individuals analyzed previously by Hauffe et al. (2010).
M. glareolus previously screened in Fevola et al. (2017b).
Previously screened in Romeo et al. (2014).
Crocidura leucodon and Crocidura suaveolens (one sample of C. leucodon from Italy was LV-RNA PCR positive).
Sorex alpinus, Sorex antinorii, Sorex coronatus, and Sorex minutus (two samples of S. antinorii from Italy were LV-RNA PCR positive).
Baltic states, Estonia, Latvia, and Lithuania; DE, Germany; FI, Finland; FR, France; HR, Croatia; IT, Italy; LV, Ljungan virus; NL, the Netherlands; SE, Sweden; SI, Slovenia; SK, Slovakia.
In the three Baltic States (Estonia, Latvia, and Lithuania), as well as Croatia, France, Germany, Italy, Slovakia, Slovenia, and the Netherlands, animals were live trapped and euthanized the same day, and liver samples were dissected out and stored at −20°C or lower. Additional animals were collected in Germany by snap trapping as described previously (Fischer et al. 2018). In Finland and Sweden, animals were snap trapped using Finnish traps (Etutuote Ky, Vaasa, Finland) or FOX metal traps (DeuFa Fallen Produktions- und Vertriebs GmbH, Neuburg, Germany), and either dissected immediately or frozen at −20°C and thawed and dissected at a later date. Liver samples from all countries were shipped to the Fondazione Edmund Mach, Italy, on dry ice for RT-PCR analyses.
The “age class” (functional groups related to maturation and breeding) of bank voles was estimated using body mass as a proxy (Escutenaire et al. 2002, Tersago et al. 2011), for example, individuals were classified as “juvenile” (<14 g), “subadult” (14–17.9 g). and “adult” (≥18 g), in combination with other indicators (Prévot-Julliard et al. 1999). Individuals with descended or undescended testes and penile shaft were classified as males, while those with mammary glands (with or without visible nipples) and a vagina (open or closed) were classified as females (Jackson 1912).
Field work and small animal trapping were carried out with permission from the ethics committees in the respective countries according to their national laws (Czech Republic: authorized in the protocol PP 27/2007) [institutional and state committees of the Czech Academy of Sciences in 2007]; France: authorized under French and European regulations on care and protection of laboratory animals: French Law 2001–486 issued on June 6, 2001, and Directive 2010/63/EU issued on September 22, 2010; all animal procedures (trapping and euthanasia) were preapproved by the Direction des Services Vétérinaires of the Herault Department under Agreement B 34-169-1; Finland: snap trapping does not require ethical permits under the Finnish Act on Animal Experimentation 62/2006 and by the decision of Finnish Animal Experiment Board 16 May 2007; however, permit n. 23/5713/2001 for capturing protected species (Sorex spp., M. rufocanus and M. schisticolor) was granted by the Finnish Ministry of the Environment; and Germany: small mammals were trapped using Sherman©live animal traps according to relevant legislation (H.B. Sherman Traps, Inc., Tallahassee, FL; official permit Site R1:Regierung der Oberpfalz 55.1–8646.4–140, Site T:Regierung von Schwaben 55.1-8646-2/30, Site S: AZ 36.11–36.45.12/4/12–001). Additional sample collection was authorized according to relevant legislation and by permission of the federal authorities (permits Regierungspräsidium Stuttgart 35–9185.82/0261, Landesamt für Natur, Umwelt und Verbraucherschutz Nordrhein-Westfalen 8.87–51.05.20.09.210, Landesamt für Landwirtschaft, Lebensmittelsicherheit und Fischerei Mecklenburg-Vorpommern 7221.3–030/09, Thüringer Landesamt für Lebensmittelsicherheit und Verbraucherschutz 22-2684-04-15-107/09); Italy: authorized by the “Comitato Faunistico Provinciale della Provincia di Trento,” protocol no. 595, issued on May 4, 2011; the Netherlands: authorized in compliance with Dutch laws on animal handling and welfare: RIVM/DEC permits 200700119, 200800053, 200800113, and 20100139; Slovakia: authorized according to current laws of the Slovak Republic, approved by the Ministry of Environment of the Slovak Republic, licence numbers 297/108/06–3.1, 6743/2008–2.1, and ZPO-594/2012-SAB; Slovenia: authorized by the Ministry of Culture of the Republic of Croatia (No. 532–08–01-01/1–11-03) and the Veterinary Administration of the Republic of Slovenia (No. 34401–36/2012/9; Sweden: authorized under the Animal Ethics Committees of Umeå: A 44–08, A 61–11, and A 121–11, the Swedish Board of Agriculture: A 135–12 and Dnr A78–08, and the Swedish Environmental Protection Agency: Dnr 412-2635-05, Dnr 412-4009-10, Nv 02939–11).
Molecular analysis
RNA extraction was performed using the RNeasy Lipid Tissue Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer's instructions. Using an LV-specific one-step RT-PCR method targeting a 185 nucleotide (nt) sequence of the 5′ untranslated region (UTR) (Donoso-Mantke et al. 2007), liver samples were screened for LV-RNA and confirmed by sequencing as previously described in Fevola et al. (2017b). RT-PCR products were purified either directly or from agarose gel using the PCR Purification Combo Kit (Invitrogen, Carlsbad, CA) in combination with the PureLinkt Quick Gel Extraction kit (Invitrogen). Purified DNA was sequenced in both directions using a BigDye Terminator cycle sequencing kit (Perkin Elmer, Applied Biosystems Division, Foster City, CA) following manufacturer's instructions, with a final volume of 10 μL. The dye-labeled sequencing products were run on an ABI PRISM 3100 Genetic Analyzer (Applied Biosystems).
Results were edited using BioEdit (version 7.2.5) (Hall 1999), and then assembled using Sequencher DNA sequence analysis software (version 4.7; Gene Codes, Ann Arbor, MI). Sequences were confirmed as LV-specific using BLASTn (Basic Alignment Search Tool 2017; National Center for Biotechnology Information, Bethesda, MD). Unique sequences were identified using DNAcollapser (
A cartogram of LV-RNA presence and distribution for all small mammal species combined with sampling effort (Fig. 1) was generated using ScapeToad (
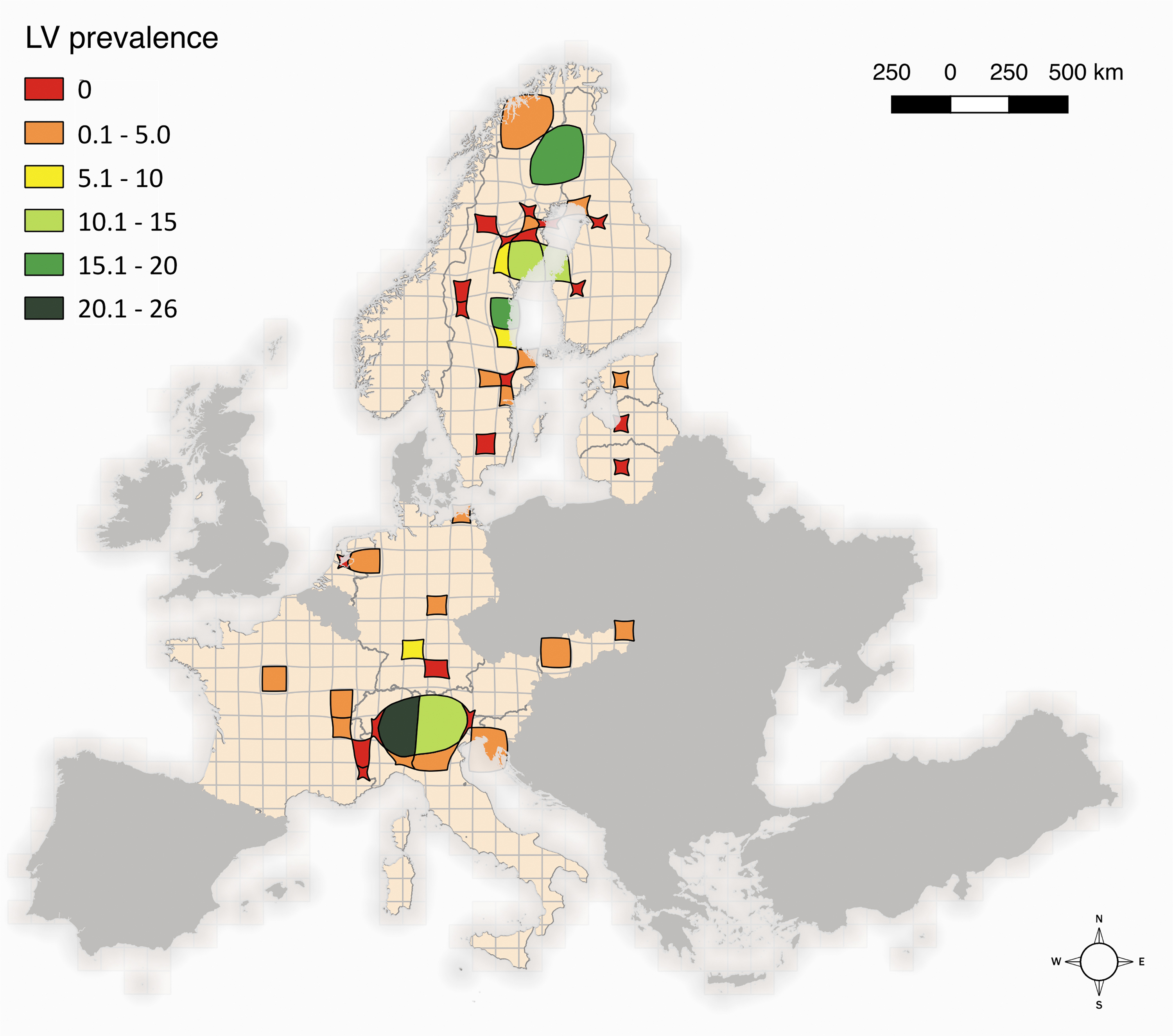
Cartogram of Ljungan virus RNA prevalence (%) in small mammals in Europe, showing how this varies with sampling effort. Sampling effort is given by the size of the cell, with the original size (10 km2) varying by the proportion of sampling occurring there. Sampling effort was calculated using the coordinates of each trapping site: the more samples from each cell, the greater was sampling effort for that cell.
Statistical analysis
All statistical analyses were performed using the R package stats (R Development Core Team 2016).
Geographic distances were calculated between each pair of LV-RNA PCR-positive individuals, taking into account the earth's curvature, using the R package raster (R Development Core Team 2016). To test the correlation between genetic and geographical pairwise distances, simple Mantel tests (Mantel 1967) were used in the R package ape.
Land cover information was obtained from the European Corine Land Cover (CLC) map (CLC 2006, version 17,
Mean land surface temperature and precipitation (daily mm of precipitation and number of days with precipitation) were estimated for the 6 months preceding each trapping date. Mean temperatures were obtained from the EuroLST database of gap-free daily MODIS land surface temperature (LST) maps (available at the Fondazione Edmund Mach) (Metz et al. 2014), with a spatial resolution of 250 meters, while precipitation data were extracted from the ECA&D dataset available at
To assess the association of individual (intrinsic) and environmental (extrinsic) factors with LV occurrence in bank voles in Europe, all available data on infection status of each bank vole from Finland, France, Germany, Italy, and Sweden were analyzed in a Generalized Linear Mixed Model with a binomial error distribution. The binary response variable was the presence or absence of LV-RNA, while explanatory variables were sex, “age class,” body mass, quadratic body mass, trapping season, country, trapping site (latitude/longitude/altitude of trapping location), land cover, mean daily temperature, and mean daily precipitation of the 6 months preceding the trapping day. Preliminary analysis (Pearson's correlation and variance inflation factor) indicated collinearity among some explanatory variables; therefore, the following were removed: country, longitude, temperature, and “age class” of individual voles. In addition, we considered only agricultural and forest land cover categories, as other categories were undersampled. As voles in different countries were not sampled in the same years (Table 1), year was treated as a random effect. Starting from the maximum model, which included all noncollinear explanatory variables, we carried out a multimodel selection approach (Burnham and Anderson 2002) based on Akaike Information Criterion correction (AICc) (Akaike 1974); the model with fewer parameters within the range ΔAICc <2 was selected as the best parsimonious model.
We also applied a generalized linear model (GLM) to determine differences in the LV prevalence in bank vole samples collected in Italy and Sweden, from which we had animals trapped for 3 consecutive years (2010–2012). In this case, year was treated as a quantitative fixed effect. The explanatory variables and model selection procedure for this analysis were the same as those listed above. In this case following collinearity analysis the following variables were removed: longitude, altitude, temperature, land cover, and “age class” of individual voles. All quantitative explanatory variables in both models were standardized before including them in the models.
Results
A total of 1685 rodents and insectivores belonging to 21 species from 12 European countries were collected and screened using LV-specific RT-PCR (Table 1). In total, LV-RNA was detected in samples from nine out of 12 countries, and in 12/21 species tested, including 1 of 2 commensal species: the house mouse (but not the black rat Rattus rattus) (Table 1). The only invasive species (the gray squirrel Sciurus carolinensis) was also LV-RNA PCR negative, as were the striped field mouse (Apodemus agrarius), wood mouse (Apodemus sylvaticus), and common vole (Microtus arvalis), although it should be noted that these latter three species all had small sample sizes, and/or were collected from very few countries (Table 1). In this study, LV-RNA was detected for the first time in two vole species, the red vole (Myodes rutilus), and the root vole (Alexandromys oeconomus), as well as in two insectivore species, the bicolored white-toothed shrew (Crocidura leucodon) and the Valais shrew (Sorex antinorii) (Table 1).
Overall, for all countries and species, LV-RNA PCR prevalence was 9.8% (166/1685), while overall LV prevalence in bank voles was 15.2% (135/885), ranging from a minimum of 3.7% (in Slovakia) to 25.9% (in Italy) (Table 1). Figure 1 shows that the highest overall LV-RNA PCR prevalence in small mammals is found in the boreal area in northern Fennoscandia and in the alpine area in northern Italy, with ranges of 15–26%. In other areas, LV prevalence was lower (0.1–5.0%), with only three sites with a prevalence ranging from 5% to 10%; in several areas, LV was not detected at all. However, as Fig. 1 indicates, the highest values of LV prevalence were detected in trapping areas with a more intense sampling effort.
Out of the 185 nt LV-specific sequences, 99 sites (53.5%) were conserved, especially at the 5′ and 3′ ends of this UTR fragment. Our sequences represented 80 haplotypes, which displayed no geographical or host-specific patterns (data available on request). In some cases, the same LV haplotypes were found in different animal species; for example, individuals of the bank vole, house mouse, and yellow-necked mouse from different trapping sites occasionally carried the same haplotype. On the other hand, certain haplotypes were also found present in several countries, for example Finland and Italy, or Sweden and Italy. Seventy-three out of the 80 haplotypes included at least one LV sequence obtained from bank voles (data not shown). Genetic distance (sequence variation) showed a small, but significant increase with increasing geographical distance when the whole dataset was considered (Mantel test, p < 0.01, although this is based on a low R 2 value (0.093).
Land cover analysis (R code available on request) identified 3 CLC level 1 classes (agricultural areas, forests, and seminatural areas, and artificial surface), 9 CLC level 2 classes (arable land, artificial, forest, heterogeneous agricultural areas, industrial, pastures, permanent crops, shrub and/or herbaceous vegetation associations, and urban fabric), and 15 CLC level 3 classes (annual crops associated with permanent crops, broad-leaved forest, complex cultivated patterns, coniferous forest, discontinuous urban fabric, fruit trees and berry plantation, land principally occupied by agriculture with significant areas of natural vegetation, mixed forest, moors and heathland, natural grasslands, nonirrigated arable land, pastures, sport and leisure facilities, and transitional woodland shrubs) in the study area. Overall, LV prevalence was higher in small mammals trapped in forested areas, that is, coniferous forests (48.8%) and woodland shrubs (18.7%).
The best parsimonious model for assessing which factors influenced LV-RNA occurrence in bank voles trapped in five European countries between 2006 and 2012 included the following explanatory variables: body mass (and its quadratic term), trapping season, altitude, and mean precipitation in the 6 months preceding the trapping date (Table 2). The effect of the quadratic body mass was significant (p = 0.02), meaning that LV prevalence was higher in animals with an intermediate body mass. Compared to the voles trapped in autumn, LV occurrence was significantly lower in spring (p = 0.02), but not in summer (p = 0.07). The occurrence of LV-RNA in bank voles was higher in trapping sites located at higher altitudes (Table 2). Interestingly, the amount of precipitation was negatively correlated with LV occurrence (p < 0.05), that is, high precipitation in the 6 months preceding the trapping date was significantly correlated with a decrease of LV occurrence in our study.
Coefficient Estimates and Statistics of Explanatory Variables Remaining in the Best Parsimonious Selected Models for Predicting the Occurrence of Ljungan Virus RNA in Bank Voles in Five European Countries from 2006 to 2012
All estimates refer to standardized variables. Reference value was autumn for “Season.”
Estimate to standard error ratio.
Two-tailed p values for Z statistics; significance level: * p < 0.05; ** p < 0.01; *** p < 0.001.
Mean precipitation for the previous 6 months.
The best model for assessing the occurrence of LV RNA in bank voles trapped in Italy and Sweden in the period 2010–2012 included body mass (again with a positive linear term and a negative quadratic term), the year, the season and country of capture, and the mean precipitation in the 6 months preceding the date of capture (Table 3). The GLM results showed once again a significant and negative association of individual body mass with LV occurrence (p < 0.001). In particular, the effect of the year is negative (p < 0.001), indicating a downward trend in LV prevalence from 2010 to 2012; LV occurrence was significantly lower both in spring and in summer with respect to autumn (p < 0.001); and LV-RNA PCR prevalence in bank voles detected in Italy was higher than in Sweden (p < 0.001). Finally, also in this model, the amount of precipitation recorded during the 6 months preceding the trapping date was negatively correlated with LV occurrence (p < 0.05) (Table 3).
Coefficient Estimates and Statistics of Explanatory Variables Remaining in the Best Selected Models For Predicting the Occurrence of Ljungan Virus RNA in Bank Voles in Italy and Sweden only, from 2010 to 2012
All estimates refer to standardized variables. Reference values were Italy for “Country” and autumn for “Season.”
Estimate to standard error ratio.
Two-tailed p values for Z statistics; significance level: * p < 0.05; ** p < 0.01; *** p < 0.001.
Mean precipitation for the previous 6 months.
Discussion
The prevalence of LV-RNA in 21 species of rodent and insectivore was assessed to document the geographical and host distribution of LV in 12 European countries, and to better understand the association of LV prevalence in bank voles with various intrinsic and extrinsic factors.
In this study, we showed that LV is widespread, both geographically (9 out of 12 countries) and among potential rodent and insectivore host species (12 out of 21 species screened in this study), although not in all tested commensal species, such as rats (see also Cuong et al. 2015). This brings the total number of LV-positive countries to 12 (those listed in Table 1 plus Denmark, United Kingdom, and the United States), and potential small mammal host species to 14 (those listed in Table 1 plus the montane vole and southern red-backed vole), including 8 vole species.
The overall LV-RNA PCR prevalence of 9.8% (all hosts and countries; standard error: 1.7% per country, 1.9% per species) is within the range of our previous screening of bank voles in Sweden and Finland only (16.2%) (Fevola et al. 2017b), and bank vole screenings in other countries by other authors (United Kingdom: 27.0%; Salisbury et al. 2014; Finland: 18%; Jääskeläinen et al. 2013; Germany: 8.4%; Kallies 2010; Italy: 50.0%; Hauffe et al. 2010). Prevalence in the yellow-necked mouse and the house mouse is lower compared to previous reports (yellow-necked mouse: 10%, Hauffe et al. 2010; house mouse: 25%, Kallies 2010; 26.1%, Salisbury et al. 2014), but within the previously reported range for field vole (17%, Kallies 2010, 0.6%, Forbes et al. 2014).
In Finland and Italy, where samples from several species were available, we found LV-RNA PCR-positive individuals from almost all the species. In particular, in Finnish Lapland (2 sites) 7 out of 8 rodent species screened were LV-RNA PCR positive, and in northern Italy (4 sites), 6 out of 12 were positive (Table 1). For both these examples, bank voles had the highest prevalence of all potential host species, implying they are the main host for LV, as has been suggested previously (Tapia et al. 2010, Fevola et al. 2017b). Nonetheless, susceptibility of potential host species to LV should be tested by experimental infection before they can be considered true hosts. The presence of LV-RNA PCR-positive animals of other rodent and insectivore species with a lower or much lower prevalence may be a consequence of a spillover from bank voles to other susceptible species. This “spillover” phenomenon has been described for Puumala orthohantavirus (PUUV) between bank voles and wood mice, for Dobrava-Belgrade virus (strain Dobrava) between striped field mice (A. agrarius) and yellow-necked mice, and for Tula orthohantavirus between common and field voles sharing the same habitat (Klingström et al. 2002, Schlegel et al. 2009, Schmidt et al. 2016). However, this is not always the case, since LV-RNA was clearly present in rodent communities at Kilpisjärvi, Finland (northern-most polygon in Fig. 1; although at lower prevalence than that found in the bank vole elsewhere), where bank voles do not occur.
The distribution of LV-RNA PCR-positive individuals was heterogeneous across study sites. If the transmission of LV is direct and density dependent, the spread and persistence of the virus would depend on the number and type of contacts between hosts (Swinton 1998, Heyman et al. 2012). Because the density of small mammal species is different in various habitats (Escutenaire et al. 2002), various biotic and abiotic factors could influence the spatial heterogeneity of pathogen transmission (Sauvage et al. 2003). In our study, the highest LV prevalence was detected in trapping areas with a more intense sampling effort, as expected, since a higher sampling effort translates into a larger number of samples collected in that area, with a consequent higher probability of detecting the virus.
Our results showed a correlation between body mass and LV prevalence, in that bank voles with an intermediate body mass were the most infected both overall, and when only Italy and Sweden were considered. Since body mass is correlated with functional group, these results support those of Fevola et al. (2017b), which showed a correlation between the “age class” of bank voles and LV prevalence, in that nonbreeding subadults with intermediate body mass were more frequently infected than (lighter) juveniles and (heavier) breeding adults. The results of these two studies together suggest that dispersal might be an important factor contributing to transmission, since in bank voles, it is primarily subadults (individuals trapped in autumn that did not attain maturity during their first year) that show density-dependent dispersal (e.g., Hansson 1987, Ecke et al. 2002). However, unlike PUUV infection, which is chronic and results in the oldest animals having the highest prevalence (Vapalahti et al. 2003, Kallio et al. 2007, Reusken and Heyman 2013, Voutilainen et al. 2016), this study suggests that LV is not persistent, and bank voles might clear the infection later in life. Interestingly, in human LV serological studies, waning immunity with age has also been observed (Jääskeläinen et al. 2016).
Overall LV prevalence was higher in small mammals trapped at higher altitudes and in forested areas, particularly in woodland, broad-leaved forest, and coniferous forests compared to arable land and heterogeneous agricultural areas. Since forest environments provide more favorable bank vole habitat (higher quality food resources and more stable climate) than fragmented, heterogeneous landscape (Guivier et al. 2011, Heyman et al. 2012), the higher prevalence in woodland may be due to bank vole density and behavior in this habitat type. Forest fragmentation also seems to decrease the prevalence of PUUV in Belgium for the same reasons (Linard et al. 2007, Vaheri et al. 2013, but see Magnusson et al. 2015 for Sweden). However, our results were heavily influenced by the high number of forest-dwelling bank voles screened in these habitats in Italy and Sweden compared to species living in arable land or other habitats. Favorable habitat might influence population dynamics in other ways that influence pathogen transmission, for example, higher turnover or population growth rates result in a higher proportion of younger nonbreeding subadult hosts, which the results of this study indicate are more likely to be infected with LV.
Our study showed that high precipitation in the 6 months before the trapping date was significantly correlated with a decrease of LV prevalence in bank voles. This correlation could be due to the effect of climatic conditions on the survival of viruses in the environment outside their hosts (Hurst et al. 1980, Kallio et al. 2006), or the survival of infected host. The survival of LV in the environment is not known (but see Ekström et al. 2007 for laboratory studies), nor is the survival of LV-infected bank voles. However, our results are consistent with previous studies that observed the effect of weather conditions on host activity, which in turn affected their exposure to viral infection. For example, precipitation during the autumn before sampling was shown to negatively influence PUUV prevalence in bank voles (Linard et al. 2007). As rainfall tends to diminish the activity of bank voles and thus the contact between individuals (Wróbel and Bogdziewicz 2015), it follows that the potential to transmit the virus may also decrease with increased rainfall. In addition, environmental regulators such as climate and habitat quality affect both direct and indirect transmission, through their effect on rodent reproductive success, host population abundance, and dispersal (Kozakiewicz 2000, Davis et al. 2005, Mills 2005, Linard et al. 2007). For PUUV, it is known that viral transmission between bank voles is driven by viral excretion into the environment and the survival of the virus outside the host (Kallio et al. 2006, Heyman et al. 2012, Voutilainen et al. 2015), LV transmission, and the factors affecting it still need to be elucidated.
Temporal variation was also noted in LV prevalence, that is, overall prevalence was significantly lower in spring compared to autumn. Because vole population density is usually the highest at the beginning of autumn just before the end of the breeding season (Bujalska 1996, Tadin et al. 2014) and includes a higher proportion of subadult animals, a higher prevalence at this time of year might be due to closer contact among nonterritorial and nonaggressive subadult infected and susceptible individuals, thus increasing the possibility of virus transmission between them. Moreover, since animals' immune statuses can be compromised during the breeding season, due to demanding physiological activities (Martin et al. 2008), this may increase the probability of breeding adults getting infected by increasing susceptibility.
The lower prevalence of LV in bank voles in spring is coherent with the hypothesis of the clearing of the infection from bank voles as they mature, as already proposed by Fevola et al. (2017b) and as already mentioned, but also with high winter mortality of this species, which may be exacerbated by the virus (Niklasson et al. 2006). There was no association of LV prevalence with sex; however, if vole activity is at the core of LV transmission, we would expect males to be more infected, as their mobility in the reproductive season is markedly higher compared with females, and they frequently leave their home range and visit shelters of other animals (Bernshtein et al. 1999).
We found a high level of variation among LV sequences (46.5% of variant sites), even though the 185 nt LV-specific fragment is from the UTR, a purportedly “conserved” part of the genome (Johansson et al. 2002, Donoso-Mantke et al. 2007). The 5′ UTR is the most highly conserved region in many other virus genomes, such as those of enteroviruses, the foot-and-mouth disease virus (FMDV) in the Picornaviridae family, and the hepatitis C virus (HCV), a member of the Flaviviridae family. In Smith et al. (1995), 77% of nucleotide positions in the 5′ UTR were invariant among all the sequences of HCV surveyed, and between the enteroviruses and FMDV strains, the average sequence similarities were 80–95% (Rivera et al. 1988, Santti et al. 1999, Carrillo et al. 2005).
The high number of haplotypes found in this study indicates that LV maintains a high level of variation compared to other viruses, possibly as a result of a wide host range, necessitating adaptation to different host genomes and immunities. However, in contrast to PUUV and Tula orthohantavirus virus (see Saxenhofer et al. 2017), only weak isolation by distance was revealed for LV. This result could be explained by the relatively short length of the fragment analyzed and needs to be confirmed. In contrast, whole genome studies of Pounder et al. (2015) suggested that LV was evolving slowly. However, that study used laboratory strains of the LV virus. Further phylogenetic analyses of LV are ongoing to better characterize the various LV strains and understand its evolutionary history, especially because, due to the high strain diversity in Parechoviruses, it is not clear if rodent and humans strains of LV are the same (Jääskeläinen et al. 2016).
Our results show that LV is widespread across Europe and its micromammal communities, and provides further support for the bank voles as the main wild host (Fevola et al. 2017b), particularly subadults. In addition, we noted a temporal pattern of prevalence (higher in autumn), and we show for the first time that higher prevalence is associated with lower precipitation, woodland habitats, and higher altitude; both these results are likely associated with subadult bank vole behavior affecting transmission. Although the zoonotic potential of LV is now considered unlikely (Fevola 2019) and high human seroprevalence has not yet been linked to transmission from rodents (in fact, LV prevalence in rodent hosts in Finland—0–23%—was lower than that in humans—34–48%; Jääskeläinen et al. 2016, Fevola et al. 2017a), these variables (lower precipitation, woodland habitats, and higher altitude, and higher densities of subadults) may represent potential risk factors for human contact with the virus. Research on the zoonotic potential of LV and association of human LV seroprevalence with these environmental variables is ongoing.
Footnotes
Acknowledgments
This research was made possible by a FIRST PhD School scholarship from the Fondazione Edmund Mach to C.F. We also thank Edvinas Rommel for help with the European CLC map. The article is cataloged by the EDENext Steering Committee as EDENext385 (
Author Disclosure Statement
No conflicting interests exist.
Funding Information
Laboratory work was funded by the European Union grant FP7–261504 EDENext - Biology and control of vector-borne infections in Europe to A.R., H.H., R.G.U., and H.C.H. and by Sigrid Jusélius Foundation. Sample collection in Sweden was financed by grants from the Swedish Environmental Protection Agency (by the National Environmental Monitoring Programme for small rodents and Alvins fond) to B.H.; the Stiftelsen Oscar och Lili Lamms minne to B.H. and F.E.; VINNOVA - Swedish Governmental Agency for Innovation Systems (Verket För Innovations system) (P32060–1) to F.E.; the Swedish Research Council Formas (221-2012-1562) to F.E., B.H., and G.O.; the National Environmental and Wildlife Monitoring and Assessment program (FoMA,