
Abstract
Japanese encephalitis virus (JEV) and West Nile virus (WNV) are arboviruses primarily transmitted by Culex spp. mosquitoes. Birds are the primary hosts for JEV and WNV. Recent WNV outbreaks in Europe and United States and their association with migratory birds highlight the importance of understanding the feeding host preference of potential vectors for outbreak preparedness, especially in nonendemic settings. Singapore is nonendemic to JEV and WNV, but is a stopover site for migratory birds of the East Asian–Australasian Flyway. Therefore, we elucidated the feeding host range of Culex spp. mosquitoes captured in four natural (bird) habitats in Singapore from January 2011 to December 2012. We characterized feeding host DNA in field-caught mosquitoes using a PCR sequencing-based assay targeting the mitochondrial gene regions. Of 22,648 mosquitoes captured, 21,287 belonged to the Culex vishnui subgroup. The host DNA analysis showed that mosquitoes from the Cx. vishnui subgroup are opportunistic biters, feeding on a range of birds and mammals. Cx. vishnui subgroup, Culex sitiens and Culex bitaeniorhynchus, was primarily ornithophagic, although they fed opportunistically on mammals, including humans. Culex gelidus and Culex quinquefasciatus, in contrast, fed mainly on mammals. The presence of ornitho- and anthropophilic mosquito vectors and susceptible avian and mammalian hosts poses a risk spill-over transmission of JEV and WNV among humans, should these viruses be introduced through migratory birds and establish persistent transmission in resident birds and animal hosts in Singapore.
Introduction
The emergence of zoonotic pathogens and their subsequent spread have caused a significant impact on global health and economies (Jones et al. 2008, Rajarethinam et al. 2018). Changes in animal habitats and their ecosystems as part of urbanization, deforestation, and climate change have resulted in close interactions between humans and animals, leading to the emergence of zoonotic infections.
Being a tropical country with ubiquitous presence of mosquitoes, arthropod-borne pathogens pose a potential threat to public health in Singapore. Dengue, which is hyperendemic in the country, is a good example (Hapuarachchi et al. 2016, Rajarethinam et al. 2018). Even though not yet endemic in Singapore, Japanese encephalitis and West Nile fever are potential zoonotic diseases.
Japanese encephalitis is endemic in many Asian countries and contributes to an annual disease burden of ∼30,000–50,000 reported cases and 10,000 deaths (Solomon 2006, Erlanger et al. 2009, Tiwari et al. 2012). Subsequent to abolishment of pig farms in 1992, the incidence of Japanese encephalitis in Singapore dropped sharply (See et al. 2002, Ting et al. 2004, Koh et al. 2006, Wang and Liang 2015, Yap et al. 2019). However, serological surveys revealed exposure to Japanese encephalitis virus (JEV) in wild boars, dogs, chickens, cows, and goats, suggesting the likelihood of an enzootic transmission cycle in local animal populations (See et al. 2002, Ting et al. 2004, Yap et al. 2019). From 1991 to 2005, six Japanese encephalitis cases have been reported in Singapore. Of them, two cases reported in 2001 and a single case in 2005 were believed to be locally acquired (Koh et al. 2006).
In contrast, local cases of West Nile fever have never been reported in Singapore. However, given the role of migratory birds in West Nile virus (WNV) transmission (Lopez et al. 2008, Dusek et al. 2009, Reisen et al. 2010, Petrovic et al. 2013) and widespread global distribution of the virus (Ciota 2017), the risk of WNV introduction into Singapore is not null. Singapore is one of the stopover sites for migratory birds along the East Asian–Australasian Flyway (EAAF).
EAAF brings in thousands of migratory shorebirds from their breeding grounds above the Arctic Circle in Alaska and Siberia (
Despite various possibilities of pathogen introductions, the risk of sustained transmission of arboviruses and their subsequent spread in new areas are dependent on the presence of vectors, their feeding behavior, and vectorial capacity for respective pathogens.
The feeding behavior of mosquitoes is dependent on host preference and host selection (Boreham and Garrett-Jones 1973, Lardeux et al. 2007). Host preference is defined as the trait to preferentially select a particular vertebrate host as a food source, over the other species that are equally available (Boreham and Garrett-Jones 1973). Host selection, in contrast, is defined as the feeding pattern in nature, represented by the relative frequency of different bloodmeal sources of a mosquito population in time and space. Although the host preference is determined by numerous intrinsic physiological characteristics of the vector (Walker and Edman 1985, Harrington et al. 2001, Lyimo and Ferguson 2009), host selection is primarily influenced by ecological factors (Lardeux et al. 2007). Prior knowledge on these aspects informs valuable information for the designing and implementation of appropriate vector control strategies to mitigate pathogen transmission in the event of an introduction (Tempelis 1975).
JEV and WNV (genus Flaviviridae) are arthropod-borne viruses (arboviruses) primarily transmitted by Culex spp. mosquitoes (Vythilingam et al. 1994, Hamer et al. 2008, Golding et al. 2012, Kuwata et al. 2013, Kim et al. 2015). Mosquitoes of the Culex vishnui subgroup (consisting of Culex tritaeniorhynchus, Cx. Vishnui, and Culex pseudovishnui), as well as Culex gelidus, Culex sitiens, Culex quinquefasciatus, and Culex bitaeniorhynchus, have been implicated as the major vectors of JEV (Vythilingam et al. 1994, Van den Hurk et al. 2001, Takhampunya et al. 2011, Thenmozhi et al. 2013).
Similarly, WNV has been isolated from >40 species of mosquitoes, primarily of the genus Culex (Hubalek and Halouzka 1999, Jupp 2001, Andreadis et al. 2004, Turell et al. 2005, Godsey et al. 2013). In addition, several other laboratory studies have also proven Culex spp. mosquitoes to be competent vectors of JEV (Huang et al. 2015, 2016) and WNV (Sardelis et al. 2001, Jansen et al. 2008, Fall et al. 2014). Even though all these potential vectors of JEV and WNV inhabit Singapore (Chan et al. 2014), there is limited empirical data on their host preference patterns in the country.
In this study, we, therefore, investigated the feeding host range of local species of Culex spp. mosquitoes collected at four natural habitats over a 2-year period. We also aimed at understanding the fluctuations in Culex spp. abundance at study sites. Our findings shed light on the differences in Culex spp. composition at different sites and variations in host feeding preference in each ecological setting.
Materials and Methods
Field sites and mosquito surveys
Four mosquito sentinel sites, which are known bird habitats, were set up in 2011 as part of the JEV and WNV risk assessment program in Singapore. The sites included a wetland reserve, an avian sanctuary, and Murai Farmway in the western part of Singapore, and Pulau Ubin, an offshore island in the north east of Singapore (Fig. 1). The wetland reserve is an annual stopover site along the EAAF, where migratory birds nest from September to March. The reserve is home to numerous resident birds as well. The bird sanctuary houses both free-ranging and captive birds and also attracts migratory birds. Murai Farmway is a rural area, where many poultry farms are located. Pulau Ubin forms part of the Ubin–Khatib Important Bird Area and is known to have a diversity of visiting and resident birds as well as other animal hosts. Adult mosquito population was monitored either monthly (wetland reserve and Pulau Ubin) or quarterly (avian sanctuary and Murai Farmway) from January 2011 to December 2012. Five trap locations were set up at each sentinel site. Adult mosquitoes were trapped using the Centers for Disease Control (CDC) light traps baited with dry ice. Trapping sessions comprised two consecutive overnight trappings. Each session lasted for 16 h (including 2 h of dusk and 2 h of dawn).
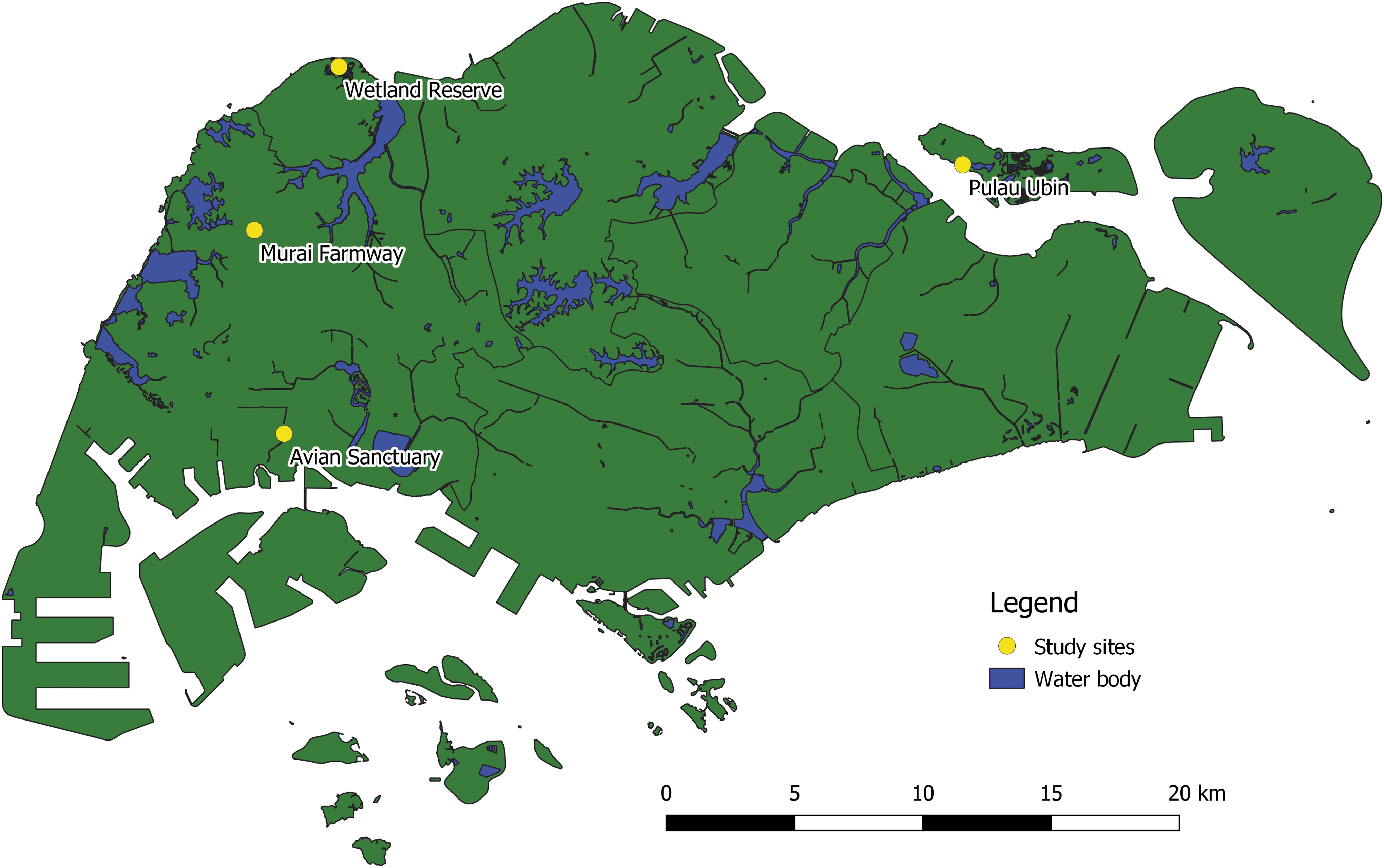
Map of Singapore with four study sites. These sites were set up as mosquito sentinel sites in 2011 as part of the JEV and WNV surveillance program in Singapore. JEV, Japanese encephalitis virus; WNV, West Nile virus.
Mosquito processing
Trapped mosquitoes were transported to the laboratory on ice. All female Culex spp. mosquitoes were identified to the species level whenever possible by using standard taxonomic keys (Bram 1967, Rattanarithikul et al. 2006). Morphologically similar species were identified to the species group level. Damaged specimens were identified to the genus level. Samples of the same species or group were pooled to include a maximum of either 50 mosquitoes (range 1–50) (Cx. vishnui subgroup excluding Cx. tritaeniorhynchus) or 10 mosquitoes (range 1–10) (Cx. tritaeniorhynchus, Cx. sitiens, Cx. gelidus, Cx. Quinquefasciatus, and Cx. bitaeniorhynchus) according to the collection site and session. All pools were stored at −80°C until further analysis.
DNA extraction
Each mosquito pool was homogenized in 500 μL of universal transport medium (Copan Diagnostics, USA) using stainless steel beads in the Mixer Mill MM 400 (Retsch Technology GmbH, Germany) at 30 Hz for 1 min. The homogenate was centrifuged at 4000 g for 1 min. Total DNA was extracted from the supernatant by using AllPrep DNA/RNA Mini Kit (Qiagen, Germany) according to manufacturer's instructions. Extracted DNA was stored at −20°C until further use.
Bloodmeal identification
Two pairs of primers described elsewhere (Kek et al. 2014) were used to amplify avian and mammalian DNA in separate PCR experiments. Amplicons generated by avian and mammalian PCR on each pool were subjected to sequencing to determine the host species/genus. These primers have been designed to avoid/minimize annealing to mosquito DNA and have been validated in numerous avian and mammalian species (Kek et al. 2014, Yeo et al. 2019). Mosquito pools were assayed in a reaction mixture (20 μL) containing 1 × Phusion™ Flash High-Fidelity PCR Master Mix (Thermo Scientific, Inc., USA), 0.5 μM of forward and reverse primers, and 2 μL of template DNA. The thermal profile for the amplification of avian DNA consisted of an initial denaturation at 98°C for 10 s, followed by 35 cycles of denaturation at 98°C for 5 s, annealing at 68°C for 8 s, extension at 72°C for 10 s, and a final extension step at 72°C for 1 min. The amplification profile of mammalian DNA was similar, except for the annealing temperature at 65°C (Kek et al. 2014). Sequencing was performed at a commercial facility using the BigDye Terminator v3.1 Cycle Sequencing Kit protocol as recommended by the manufacturer (Applied Biosystems, USA).
Consensus sequences were obtained by assembling raw contiguous reads in Lasergene version 15.01 (DNASTAR, Inc., USA). The bloodmeal identification was conducted at two different levels. First, PCR-positive mosquito pools were categorized into those containing avian and mammalian blood meals based on PCR results. Second, the avian and mammalian DNA-positive pools were subcategorized into classified and unclassified groups for the identification of genus/species of respective hosts based on sequencing results. Mosquito pools that generated individual peaks in chromatograms were considered to include DNA of a distinguishable host genus/species (categorized as classified for the genus/species identification). These consensus sequences were searched against the GenBank database using the BLASTn tool in BLAST (
Statistical analysis
Descriptive statistical parameters were calculated for the number of mosquitoes collected at respective study sites. Correlation between the variables was performed using the Pearson's correlation test. Probability values of p < 0.05 were considered significant.
Results
Mosquito species and their abundance at study sites
Trappings at four study sites collected 22,648 mosquitoes for the 2-year period. The Cx. vishnui subgroup yielded the highest number with 21,287 mosquitoes, whereof 709 were Cx. tritaeniorhynchus mosquitoes. The rest included Cx. sitiens (n = 1,037), Cx. quinquefasciatus (n = 162), Cx. gelidus (n = 119), and Cx. bitaeniorhynchus (n = 43). The number of mosquitoes trapped varied over time and between study sites (Fig. 2 and Table 1). The wetland reserve recorded the highest number of captured mosquitoes. The number of host DNA-positive mosquito pools correlated positively with that of Cx. vishnui subgroup adults captured (Pearson's correlation coefficient r = 0.84, p < 0.001), suggesting that the probability of detecting host DNA-positive pools is directly related to the number of mosquitoes screened.
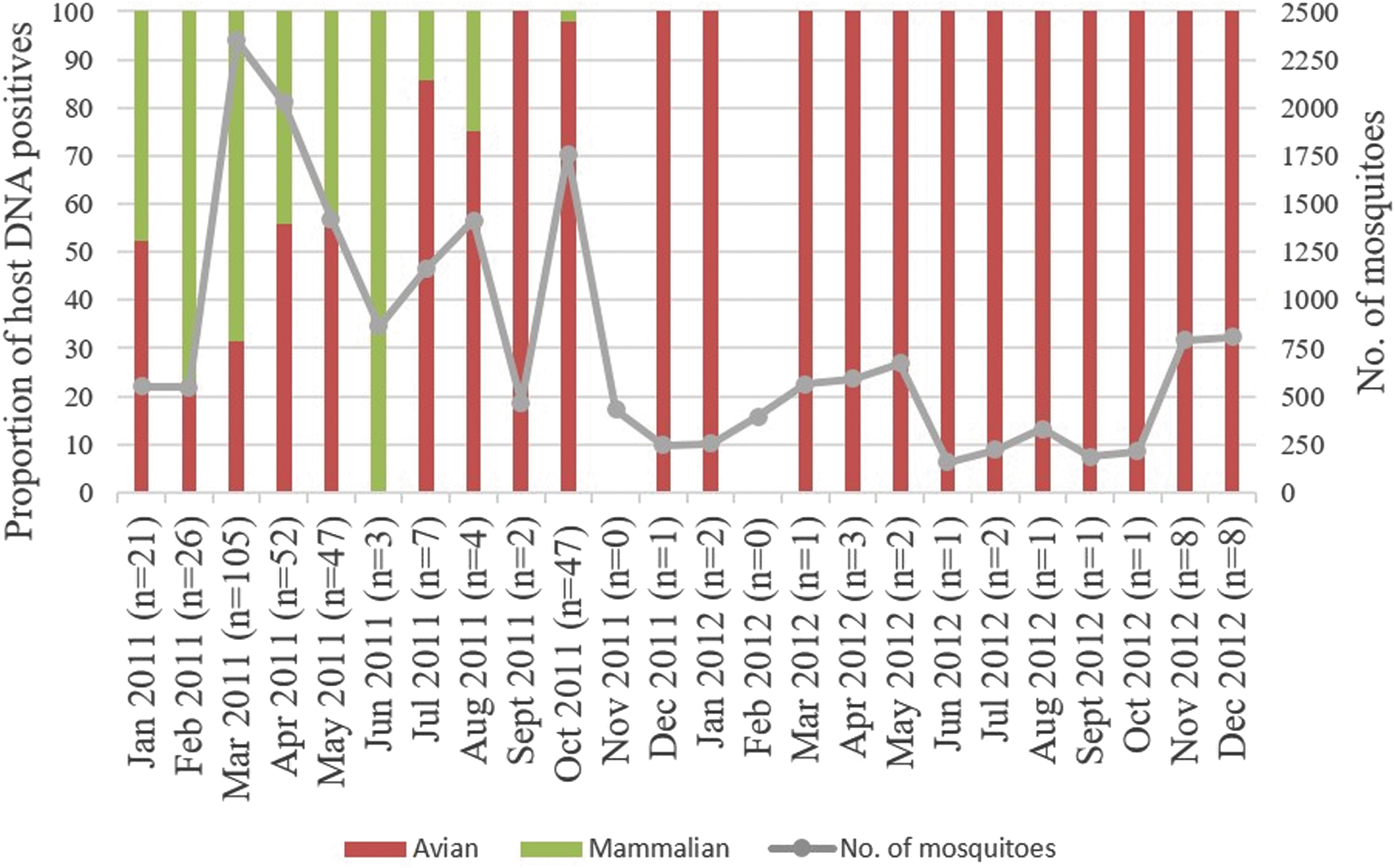
Temporal distribution of the abundance of Culex vishnui subgroup (excluding Culex tritaeniorhynchus) and the proportion of mosquito pools positive for host DNA at the wetland reserve from January 2011 to December 2012. Numbers within brackets in the x-axis show the total number of pools positive for host DNA in each month.
Abundance of Culex spp. Mosquitoes and the Positivity for Host DNA at Four Study sites
Each pool of the Cx. vishnui subgroup consisted of a maximum of 50 adult mosquitoes (range 1–50). The data exclude Cx. tritaeniorhynchus, which belongs to the Cx. vishnui subgroup. Cx. tritaeniorhynchus has previously been implicated as a primary vector of JEV in Singapore. Hence, the data for Cx. tritaeniorhynchus are given separately. The pools of all species, including Cx. tritaeniorhynchus, consisted of a maximum of 10 adult mosquitoes (range 1–10).
JEV, Japanese encephalitis virus; NA, not available.
Bloodmeal positivity among trapped mosquitoes
We screened 1739 mosquito pools, consisting of 22,648 individual Culex spp. mosquitoes (Table 1). Overall, 501 pools (29%) were positive for host DNA. Of them, 288 (57%) pools were positive only for avian DNA and 200 (40%) pools were positive only for mammalian DNA (single host category given in Table 1). The remaining 13 pools (3%) were positive for both avian and mammalian host DNA (dual host category given in Table 1). Of 501 positive pools, host genus/species could be identified in 465 pools (classified; Table 2). Of these, 452 pools were positive for the avian (n = 281) or mammalian (n = 171) host DNA (single host category given in Table 2). The remaining 13 pools were positive for the avian and mammalian host DNA (dual host category given in Table 2). Of these 13 pools, the genus/species could be classified in all avian samples. However, the genus/species could be identified only in 10 mammalian DNA-positive pools. There were 39 pools (7.7%) that were positive for either avian or mammalian host DNA (36 in the single host category and 3 in the dual host category), in which species/genus could not be identified due to multipeak chromatograms and thus were categorized as unclassified for host identification (Table 2).
Summary of Genus/Species Identification Among Host DNA-Positive Mosquito Pools from Four Study Sites
Unclassified are pools that generated multipeak chromatograms, thus the host genus/species could not be distinguished. These pools were considered to include avian and/or mammalian DNA of multiple host species.
The data exclude Cx. tritaeniorhynchus, which belongs to the Cx. vishnui subgroup. Cx. tritaeniorhynchus has previously been implicated as a primary vector of JEV in Singapore. Hence, the data for Cx. tritaeniorhynchus are presented separately.
Host preference patterns of Culex spp. mosquitoes at study sites
Cx. vishnui subgroup Cx. sitiens and Cx. bitaeniorhynchus were opportunistic feeders. Even though they fed on both mammals (humans and wild boars) and birds, they were more likely to feed on avian hosts, particularly in the wetland reserve and the avian sanctuary (Tables 2 and 3). Of 385 classified (avian/mammalian) pools of Cx. vishnui subgroup, 62% (n = 240) were positive for avian DNA (Table 2). This included 34 pools of Cx. tritaeniorhynchus. The respective proportions for Cx. sitiens and Cx. bitaeniorhynchus were 84% (47/56) and 57% (4/7), respectively. The number of Cx. vishnui subgroup pools positive for avian DNA at the wetland reserve was significantly higher (p < 0.001) than those positive for mammalian DNA. Similarly, at the wetland reserve, Cx. sitiens was six times more likely to feed on birds than mammals. These observations testified that the mentioned Culex spp. were largely ornithophilic, especially at locations where birds are abundant throughout the year. In contrast, Cx. gelidus and Cx. quinquefasciatus preferably fed on mammalian hosts (81%, 13/16 and 71%, 5/7, respectively). However, the number of positive pools for Cx. gelidus, Cx. Quinquefasciatus, and Cx. bitaeniorhynchus was relatively small to make any strong conclusion on their host preference patterns.
Biting Ratio of Culex spp. Mosquitoes at the Four Sentinel Sites
The feeding host preference of mosquito species was variable among the study sites. At the wetland reserve where the highest number of mosquitoes was caught, host DNA was detected in all species (Tables 1 and 2). As illustrated in Figs. 3 and 4, chickens, owls, herons, doves, and humans were the preferred avian and mammalian hosts at the wetland reserve. The majority of Cx. vishnui subgroup (80%, 41/51) caught at the avian sanctuary were positive for avian DNA, notably flamingos, herons, and owls, which are resident birds at the site (Table 2 and Fig. 3). In contrast, 63% (10/19) of mosquito pools collected at Murai Farmway were positive for mammalian host DNA (Table 2—including “unclassified”). Mosquito pools from Pulau Ubin contained avian and mammalian DNA in almost equal proportions, although the preference was variable among mosquito species. The biting ratio of humans was higher than that of wild boars at the wetland reserve and avian sanctuary (Table 3). The opposite was observed at the Murai Farmway and Pulau Ubin, where human settlements are either sparse or absent.
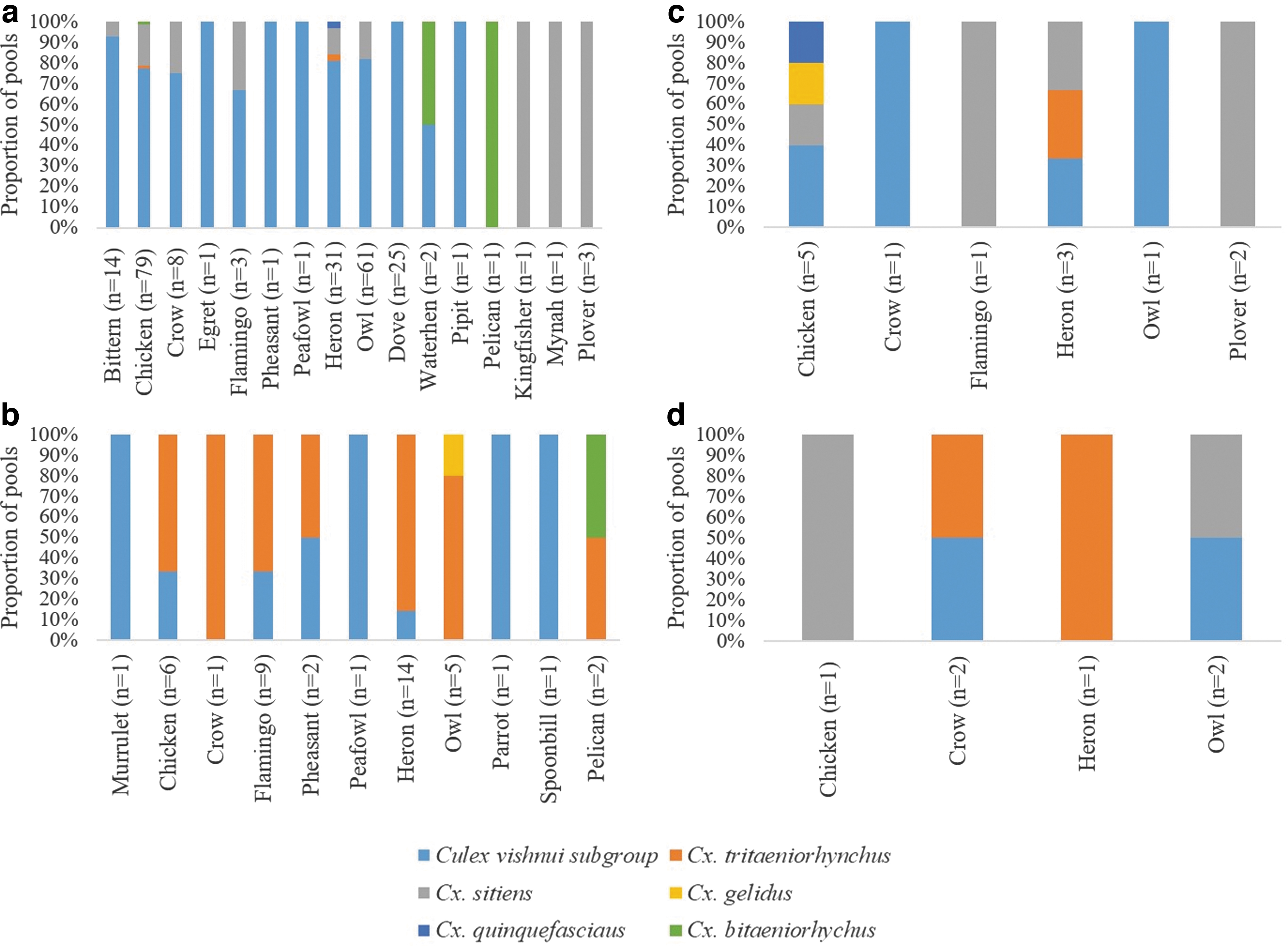
Genus/species identification of avian DNA in bloodmeals of Culex spp. mosquitoes at four sentinel sites from January 2011 to December 2012.
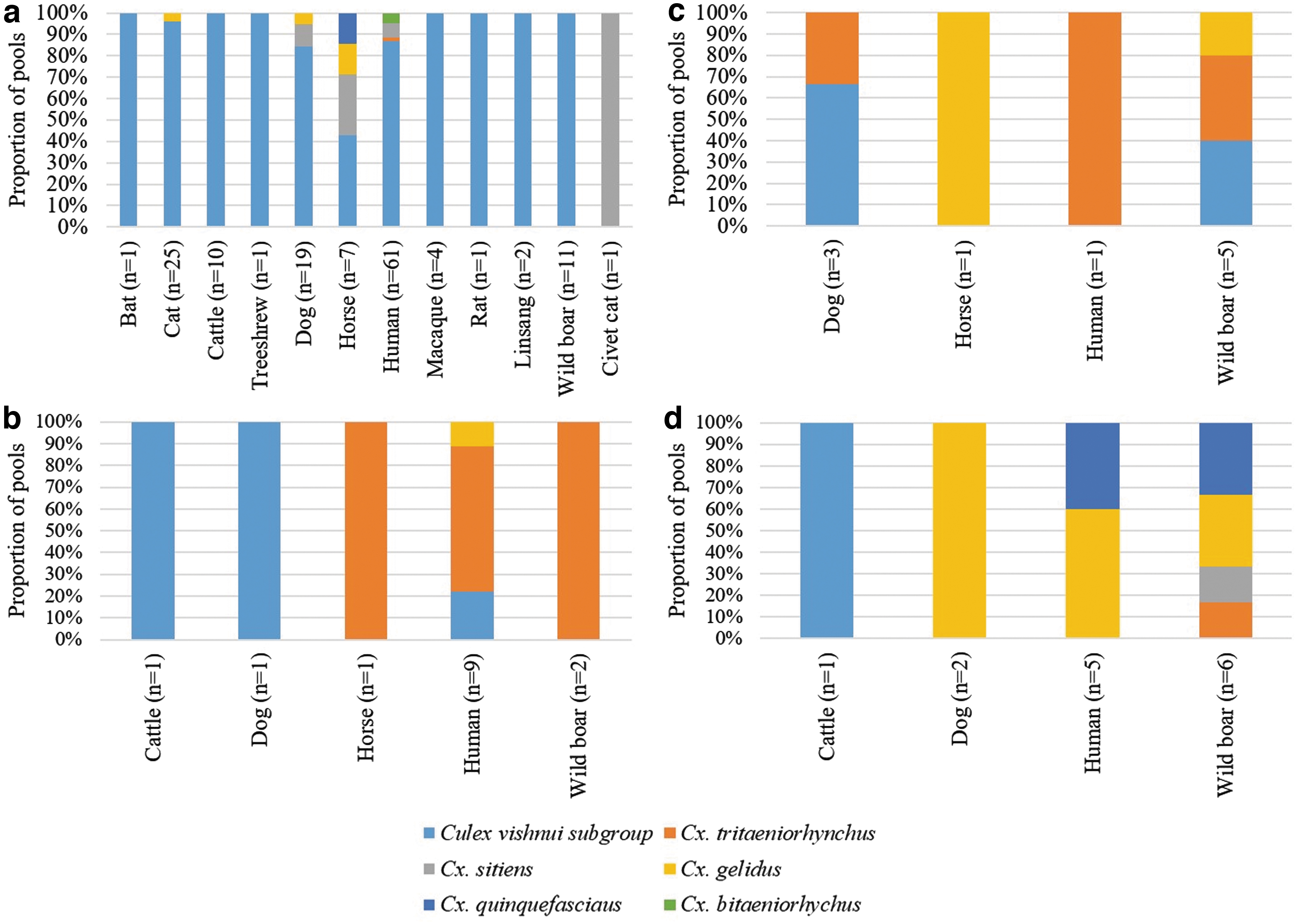
Genus/species identification of mammalian DNA in bloodmeals of Culex spp. mosquitoes at four sentinel sites from January 2011 to December 2012.
Discussion
The transmission cycle of JEV and WNV involves multiple hosts that act as reservoir hosts (birds for JEV), amplifying hosts (swine and birds for JEV and WNV, respectively), and dead end hosts (humans, horse, and cattle). Both viruses circulate in an enzootic cycle driven by Culex spp. mosquitoes (Turell et al. 2002, Savage et al. 2007, Go et al. 2014). Occasionally, these mosquitoes act as bridging vectors to infect humans, thereby to initiate an epizootic cycle. Therefore, for a particular mosquito species to be an effective vector of these viruses, it should ideally have a catholic host preference that allows feeding on multiple host species, including humans (Tuno et al. 2017). Having an equal propensity to feed on humans, as well as other animals, provides those species an edge for efficient zoonotic pathogen transmission. Overall, our findings showed that Culex spp. mosquitoes demonstrate a catholic host preference, although variable patterns among different species and locations. Cx. vishnui subgroup Cx. sitiens and Cx. bitaeniorhynchus were primarily ornithophagic, whereas Cx. gelidus and Cx. quinquefasciatus were mammalophagic. Their host selection appeared to be opportunistic and was likely dependent on host abundance and availability. For example, even though Cx. vishnui subgroup Cx. sitiens and Cx. bitaeniorhynchus preferred to feed on avian, they were opportunistic when mammals, including humans, were available. Interestingly, where humans are abundant, such as the wetland reserve and the avian sanctuary, these mosquito species preferably fed on humans. In contrast, the reverse was observed at Murai Farmway and Pulau Ubin, where more mosquito pools were positive for wild boar DNA than human DNA. This could be due to the primary difference in the abundance and availability of humans and wild boars in respective locations.
Birds play an important role in the maintenance and transmission of JEV and WNV. Members of the avian family Ardeidae (waders such as herons, egrets, and bittern) are known reservoir hosts of JEV, whereas passerine birds of the family Corvidae (crows) and house sparrows are considered the most important amplifying hosts of WNV (Nemeth et al. 2012). We detected wader bird DNA (egrets, herons, bitterns, spoonbills, and plovers) among mosquitoes from all study locations. However, sandpiper DNA was not detected in mosquitoes from any location, despite sandpipers being one of the most abundant migratory birds in Singapore. Nevertheless, chickens were the most common source of avian DNA in our mosquito pools from the wetland reserve, but were also detected from the other three locations to a lesser extent. Adult chickens develop low magnitude viremia (Banerjee and Deshmukh 1987) and, therefore, are thought to play a minor role in JEV transmission in comparison with waders (Adames et al. 1993). However, chickens at a young age develop viremia that is high enough to transmit the virus to mosquitoes (Turell et al. 2006, Cleton et al. 2014). JEV-specific antibodies had previously been detected in young chickens in JEV-endemic regions (Ogata et al. 1970). Likewise, an epidemiological link for WNV transmission had also been established between juvenile domestic chickens and mosquitoes (Maquart et al. 2016). In addition, their role as sentinels for seroconversion to WNV had been established (Chaintoutis et al. 2015, 2016, Pastiu et al. 2016, Khan et al. 2017). DNA from crows, a well-documented host for WNV (Caffrey et al. 2003, Komar et al. 2003, Yaremych et al. 2004), has also been detected in small number of mosquito pools from all four locations. Other than JEV and WNV, birds have been implicated as the primary reservoir hosts of many other mosquito-borne pathogens such as St. Louis encephalitis virus (Adames et al. 1993), eastern equine encephalitis virus (Estep et al. 2011), and Murray Valley encephalitis virus (Gubler 2002).
Whimbrels, common redshanks, common greenshanks, marsh sandpiper, Mongolian plovers, and the pacific golden plovers are the most abundant migratory birds in Singapore. They nest, especially in the wetland reserves, during each migratory season. Herons, egrets, and bitterns are common resident birds. A previous study has reported the presence of JEV neutralizing antibodies in plovers and common redshanks in Singapore (Yap et al. 2019), suggesting the exposure of common migratory birds to JEV. WNV-specific antibodies have previously been detected in migratory birds such as the turtle doves in Slovakia (Ernek et al. 1977), common coots, purple herons, and ruffs in India (Mishra et al. 2012), great cormorant and white stork in Germany (Linke et al. 2007, Seidowski et al. 2010), warblers and blackcaps in Czech Republic (Hubalek et al. 2008), and Eurasian coots in Netherlands (Lim et al. 2018). Moreover, WNV viremia has been detected in resident birds in the United States mainly during the migratory period (Dusek et al. 2009). Therefore, there is a persistent risk of JEV and WNV being imported into Singapore through migratory birds. Both the amplifying/reservoir hosts and competent vectors are present in the country. Our data showed the presence of wild boars and Culex spp. vectors of both JEV and WNV in all study sites. Wild boars are one of the major amplifying hosts of JEV (Hamano et al. 2007, Komiya et al. 2019). The findings also showed that all different species/subgroups of Culex spp. mosquitoes captured in this study fed on wild boars, although at different preferential patterns in four study sites. Moreover, we showed that these vectors feed on known hosts of JEV and WNV, such as heron, bittern, egret, and crow. These observations indicated that local ecoenvironmental conditions are favorable for the persistent enzootic transmission of JEV and WNV. In fact, serological evidence has indicated JEV transmission in animal hosts (resident birds, migratory birds, and wild boars) in Singapore (Yap et al. 2019). Our findings also demonstrated that many of these vectors opportunistically feed on humans. The wetland reserve and avian sanctuary are visitor attractions and collectively record approximately one million visitors every year. Therefore, there is a risk of JEV and WNV spillover transmission among humans, should both viruses establish enzootic transmission in resident birds and animal hosts in Singapore.
One of the challenges of our study was the difficulty in quantifying host selection by mosquito vectors due to the limited knowledge on density and availability of various hosts in study locations. This is important because the feeding behavior of mosquitoes is dependent on host preference and selection (Boreham and Garrett-Jones 1973, Lardeux et al. 2007). Previous studies have alluded this to the technical difficulties in obtaining an accurate estimation of the biting preference of vectors (Takken and Verhulst 2013). The results of these studies are often biased in favor of the most abundant host species locally available, which may not necessarily be the most preferred host. For example, collections in houses are more likely to capture mosquitoes fed with human blood, and those collected at locations with the presence of animals are more likely to be fed with animal blood (Takken et al. 2002). Cx. gelidus is known to prefer cattle to pigs, when both hosts are equally available (Reuben et al. 1992, Mwandawiro et al. 2000). It was not surprising that our findings revealed more wild boar than cattle DNA among Cx. gelidus pools captured at all study sites, since wild boars are more widespread and abundant than cattle in Singapore (Colless 1959). To address these issues, behavioral studies of host choice need to be conducted. Another limitation of the study was the inconsistency in number of mosquitoes captured over time, creating an unrealistic biasness in host preference determined based on bloodmeal analyses. For example, the biased host feeding preference of Cx. vishnui subgroup for chickens, especially in the wetland reserve, could be due to the undersampling of mosquitoes, rather than low availability of waders. Thousands of birds that migrate along the EAAF flock in the wetland reserve during the annual migratory season from September to March. The EAAF is known to bring the highest number of waders from the Northern Hemisphere to as far south as New Zealand, before making their way back to their breeding grounds. However, as illustrated in Fig. 2, the number of captured mosquitoes dropped drastically before the migratory season and subsequently maintained at a minimal level, reducing the chances of detecting bloodmeals derived from these migratory birds. This also applies to Cx. quinquefasciatus and Cx. bitaeniorhynchus collection. Finally, analyzing pooled mosquitoes is not ideal for studies that aim to investigate their feeding patterns in different ecosystems/ecological niches using sequencing-based molecular tools. The challenge is specifically at the host genus/species level identification when more than one specimen in a pool contains host DNA. These pools generate mixed chromatogram patterns that are unclassifiable to any single genus/species, when universal primers are used to amplify different groups of hosts, such as Aves and mammals. Ideally, individual mosquito specimens need to be analyzed to minimize “unclassified” bloodmeal results. However, given the large number of samples collected during this type of studies, it is not feasible to analyze individual specimens due to cost, time, and manpower considerations.
Conclusions
To the best of our knowledge, this is the first report on the host-feeding pattern of potential JEV and WNV vectors in Singapore. Taken together, these findings shed light on the biodiversity of vectors and their interactions with animal and human hosts that shape zoonotic disease transmission in different ecologies. The observations collectively suggested a potential risk of JEV and WNV introduction through migratory birds and that local ecoenvironmental conditions are favorable for the persistent enzootic transmission of both viruses in Singapore. This could eventually lead to an epizootic cycle through bridging vectors that opportunistically feed on humans.
Footnotes
Acknowledgments
We thank colleagues at NParks and the Environmental Health Institute (EHI), NEA, Singapore, for their assistance in carrying out this study.
Authors' Contributions
L.C.N. and G.Y.A.P. conceptualized and designed the study. G.L.Y., M.H., X.F.L., D.M., S.C., and C.B.H. assisted in field collection of mosquitoes and G.L.Y., M.H., X.F.L., D.M., C.S.C., S.G.P.L., and R.L. performed morphological identification of mosquitoes. G.L.Y., M.H., X.F.L., D.M., and C.S.C. were involved in the data collection, generation, processing, and analyses. G.L.Y., M.H., X.F.L., and D.M. performed DNA extraction and PCR. G.L.Y., M.H., X.F.L., and D.M. handled mosquito identification database. G.L.Y., M.H., X.F.L., and D.M. carried out data analysis and interpretation. G.L.Y., H.C.H., and G.Y.A.P. drafted the article. H.C.H., L.C.N., and G.Y.A.P. revised the article critically for important intellectual content. All authors are accountable for every aspect of the work, especially the accuracy and integrity, and have read and approved the article for publication.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The study was carried out under the zoonotic ecoepidemiology program, which is a multiagency collaboration between National Environment Agency (NEA) and National Parks (NParks) Singapore. The program was jointly funded by the Ministry of Finance (through a Reinvestment Fund) and NEA, Singapore. Funding sources had no role in the study design, data collection, analysis, interpretation, and the decision to submit the article for publication.