
Abstract
Transmission of vector-borne pathogens can vary in complexity from single-vector, single-host systems through to multivector, multihost vertebrate systems. Understanding the dynamics of transmission is important for disease prevention efforts, but is dependent on disentangling complex interactions within coupled natural systems. Ross River virus (RRV) is a multivector multihost pathogen responsible for the greatest number of notified vector-borne pathogen infections in humans in Australia. Current evidence suggests that nonhuman vertebrates are critical for the maintenance and spillover of RRV into mosquito populations. Yet, there is a limited knowledge of which mosquito vector species and amplifying vertebrate host species are most important for transmission of RRV to humans. We conducted field surveys of nonhuman vertebrates and mosquitoes in the RRV endemic city of Brisbane, Australia, to assess the effect of vector and host community structure on human RRV notifications. Six suburbs were selected across a gradient of human disease notification rates. Differences in vertebrate and mosquito compositions were observed across all suburbs. Suburbs with higher RRV notification rates contained greater vertebrate biomass (dominated by the presence of horses) and higher mosquito abundances. This study suggests that horse–mosquito interactions should be considered in more detail and that vertebrate biomass and mosquito abundance be incorporated into future RRV modeling studies and considered in public health strategies for RRV management.
Introduction
Obtaining a detailed understanding of pathogen-host-vector dynamics within a given environment is particularly challenging for multivector multihost pathogens. The relative role of different vectors and/or vertebrate reservoir species in supporting pathogen circulation is often largely unknown under natural conditions. However, an understanding of transmission is essential for the design of effective prevention and control strategies (Weinstein, 1997, Janousek et al., 2014, Hoffmann et al., 2015, Gardner et al., 2017) and for predicting net effects of zoonotic mosquito-borne diseases on human populations.
Mosquito-borne pathogens contribute significantly to the global burden of infectious disease in both human and animal populations (Hill et al., 2005). Widely distributed across Australia, Ross River virus (RRV) is Australia's most frequently notified mosquito-borne disease in humans (Australian Government, 2018). RRV is a multihost multivector pathogen with a complex ecology: it persists in all Australian bioregions and climates and has been isolated from more than 40 species of mosquito under field conditions (Russell, 2002). Humans are generally not considered important amplifying or maintenance hosts of RRV, but may play a role in outbreaks (Aaskov et al., 1981, Harley et al., 2001). However specific epidemiological studies quantifying the role of humans in transmitting RRV are limited. Similarly, the relative roles of various nonhuman vertebrate host species in maintaining transmission of RRV are poorly understood.
Strong experimental evidence supports marsupials as competent reservoirs (based on viremia profiles which are sufficient to infect susceptible vectors), but amplification and transmission competence has also been demonstrated in other species groups, including placental mammals and birds (Kay et al., 1986, Stephenson et al., 2018). Due to a myriad of factors, experimental infection studies are infrequently undertaken, and further, it is unclear how transmission competence of hosts in an experimental setting reflects their relative role in transmission in natural settings (Claflin and Webb, 2015). As such, many potential reservoir species have not yet been investigated.
Although at least 10 mosquito species have been demonstrated to have some level of vector competence of RRV in the laboratory, it is unclear what the relative contributions of each species are to both enzootic transmission and/or human disease outbreaks (Russell, 2002, van den Hurk et al., 2010, Jansen et al., 2019). It is likely that the vectors associated with human outbreaks vary across different bioregions of Australia, and this may also be the case with reservoir hosts (Claflin and Webb, 2015). Simply, the key vectors and hosts associated with the maintenance of RRV, and its spillover into humans, in different regions of Australia remain unclear.
Of the ∼5000 human RRV notifications reported in Australia each year, the majority are reported from the state of Queensland (Australian Government, 2018). Residents of Queensland's capital city, Brisbane (27.4698° S, 153.0251° E), consistently represent the highest proportion of the state's cases. In 2014–2015, Queensland experienced the largest outbreak of RRV ever recorded in Australia, totaling more than 6000 human notifications (Jansen et al., 2019, Queensland Government, 2019). Brisbane has a subtropical climate, which accommodates a diversity of habitats for both vector and reservoir species. This diversity, in combination with the persistent high human notification rates of RRV, makes the city an ideal location to study RRV disease ecology.
This study aims to assess mosquito and vertebrate host community ecology across suburbs of Brisbane encompassing locations with a range of RRV notification rates. Specifically, we aim to identify relationships between abundance, diversity, and biomass of potential nonhuman vertebrate reservoirs, mosquito vectors, and human notification rates of RRV.
Materials and Methods
Human notification data
Long-term human notification data for Brisbane suburbs were obtained for previous years (2001–2016, inclusive) from the Queensland Department of Health (ethics application: ref no. P2238). Notifications were based on laboratory evidence of probable or confirmed RRV infection, determined through (1) isolation of RRV from patients, (2) detection of RRV by nucleic acid testing, or (3) IgG seroconversion or a significant increase in IgG antibody level in previously tested patients (Australian Government, 2019), and the suburb of residence of notified cases was recorded. To account for high levels of interannual variability in RRV notifications (Supplementary Table S1), mean annual notification rates from 2001 to 2016 were calculated and used to identify areas with persistently high or low rates of RRV. At the time of this investigation, this was the most current dataset available to estimate long-term trends and select field sites. Suburbs included had a minimum population number of 1000, and rates were expressed as per 100,000 population, relative to the population size of the suburb. Suburbs were represented spatially using the “State Suburb Codes (SSCs)” geographical unit. Notifications for each of Brisbane's SSCs were matched to population data from the 2016 census, available from the Australian Bureau of Statistics (1999).
Site selection
Six SSCs were chosen across Brisbane to be included in our study (Fig. 1). SSCs were selected to represent: (1) a diverse range of human RRV notification rates, (2) varied geographical and ecological environments (urban, suburban, coastal, inland), and preferentially (3) locations included in Brisbane City Council (BCC) mosquito monitoring scheme. Study sites were identified within each of these SSCs, as described in Table 1.
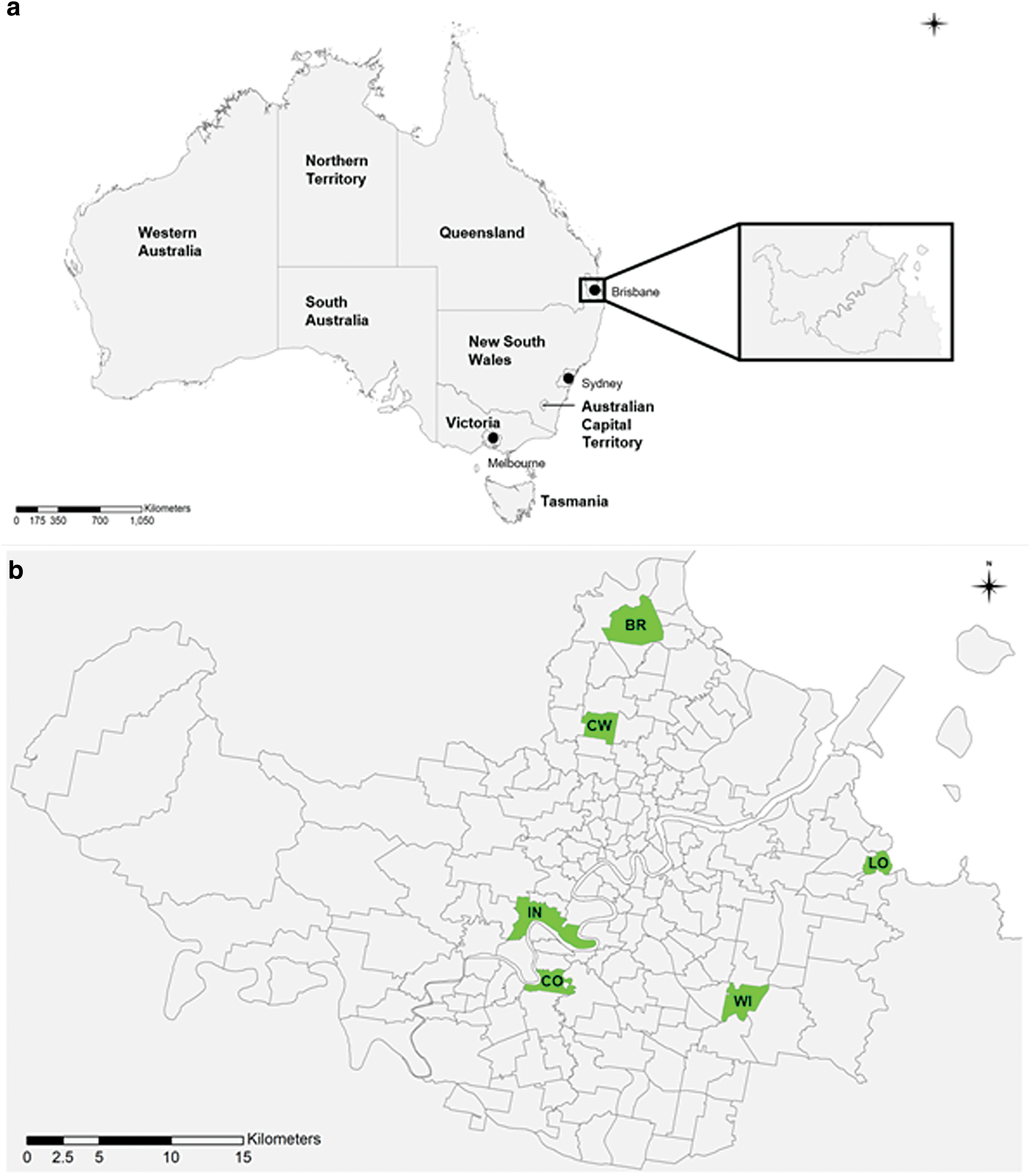
Ecological, Geographical, and Social Characteristics of Suburbs as Containing Study Sites
Human population size and density are derived from the aAustralian Bureau of Statistics 2016 census and the RRV notification rate is averaged from 2001 to 2016, from bQueensland Department of Health data.
RRV, Ross River virus; SSC, state suburb code.
Vertebrate surveys
Within each SSC, three study sites were selected to satisfy two criteria: (1) sites were required to be an accessible public park or nature reserve where it was feasible to establish transects of adequate size, and (2) at least one site within each SSC had to include the location of the mosquito trap (selected by Brisbane City Council). Each study site comprised a 100 × 100 m transect (marked using handheld GPS devices; Garmin-62S) between 50 and 1000 m apart from other sites. Dawn and dusk surveys of nonhuman vertebrates were carried out monthly in each site over a 6-month period that encompassed the peak RRV transmission season, between October 2017 and March 2018.
Methods were adapted from the “Standard Early Bird Search” and “Arboreal Spotlight Search” detailed in the Queensland Fauna Survey Guidelines (Eyre et al., 2014). In brief, dawn surveys commenced 30 min before sunrise and ran for 30 min in each site, sites within a SSC were sampled sequentially, and the order was mixed at each visit. The following data were recorded for all nonhuman vertebrate species identified: species, number of individuals, record type (i.e., seen, heard, and seen and heard), microhabitat (i.e., on the ground, in the canopy, and above the canopy), and whether the observation was in, near (<50 m), or off (>50 m) the transect. The same data were also recorded for evening surveys, which commenced after sunset and were carried out in each transect for 30 person minutes. Equipment used to identify vertebrates included binoculars, handheld torches, field guides, and the Birds of Australia mobile device application (Morcombe, 2003, Knight and Menkhorst, 2010).
Mosquito surveys
Mosquitoes were collected across the six SSCs in Brisbane using CDC-stylelight traps (Pacific Biologics, Scarborough, Australia) baited with carbon dioxide (CO2; as ∼1 kg dry ice) and 1-octen-3-ol. Trapping was undertaken by BCC as part of its ongoing mosquito surveillance and control program. Traps were set once per week before dusk (starting the 5th of September 2017, ending the 26th of March 2018) and collected the following morning. A single trap was located within one of the three sites in each SSC used for vertebrate surveys. Traps were set 1.5 m off the ground in proximity to a productive larval mosquito habitat. Mosquitoes were identified to species level with the aid of morphological keys (Lee et al., 1989, Webb et al., 2016).
Data analysis
Variable analysis
Community ecology variables, including biomass (vertebrates only), abundance, and species diversity, were calculated for vertebrate and mosquito communities within each site. Species diversity was calculated using the Shannon–Weiner index (Boyle et al., 1990), which accounts for both the abundance and evenness of a given species in a community. Biomass was calculated by multiplying the total number of observed individuals for a given species by the average adult body mass (kg) of that species and log transformed for analysis. Body mass was derived from the PANTHERIA dataset for mammals (Jones et al., 2009), and study of Garnett et al. (2015) was used for birds. Generalized linear models (GLM) were used to test whether human notification rate for RRV correlated with a given community ecology variable.
Community analysis
Vertebrate and mosquito communities within each SSC were analyzed in two ways. First, stacked column charts were constructed to visually represent community composition. Second, to examine the effect of community compositions on human notification rates, nonmetric multidimensional scaling (NMDS) analyses were performed using the vegan R package (Oksanen et al., 2010) and BiodiversityR (Kindt, 2019). These ordinations analyzed community composition of vertebrates and mosquitoes (separately) for each month at each SSC.
The data were auto-transformed and scaled using Bray Curtis dissimilarity indices using the “metaMDS” function. The degree of stress for each NMDS plot was calculated, which indicates the reliability of the outcome, that is, lower stress corresponds with a higher reliability. The ordination of elements was considered arbitrary for stress values of 0.3 or above. The dissimilarity matrices were based on abundances for both vertebrates and mosquitoes, and an additional biomass ordination was generated for vertebrates.
Results
Human cases of RRV within each SSC
The mean annual notification rate across all Brisbane suburbs was 31 cases/100,000 people between 2001 and 2016. The six SSCs selected for our study exhibited wide variation in the rate of reported RRV (Fig. 2). Of these, Corinda reported the highest mean annual notification rate, followed by Bracken Ridge, Chermside West, Indooroopilly, Wishart, and Lota, respectively.
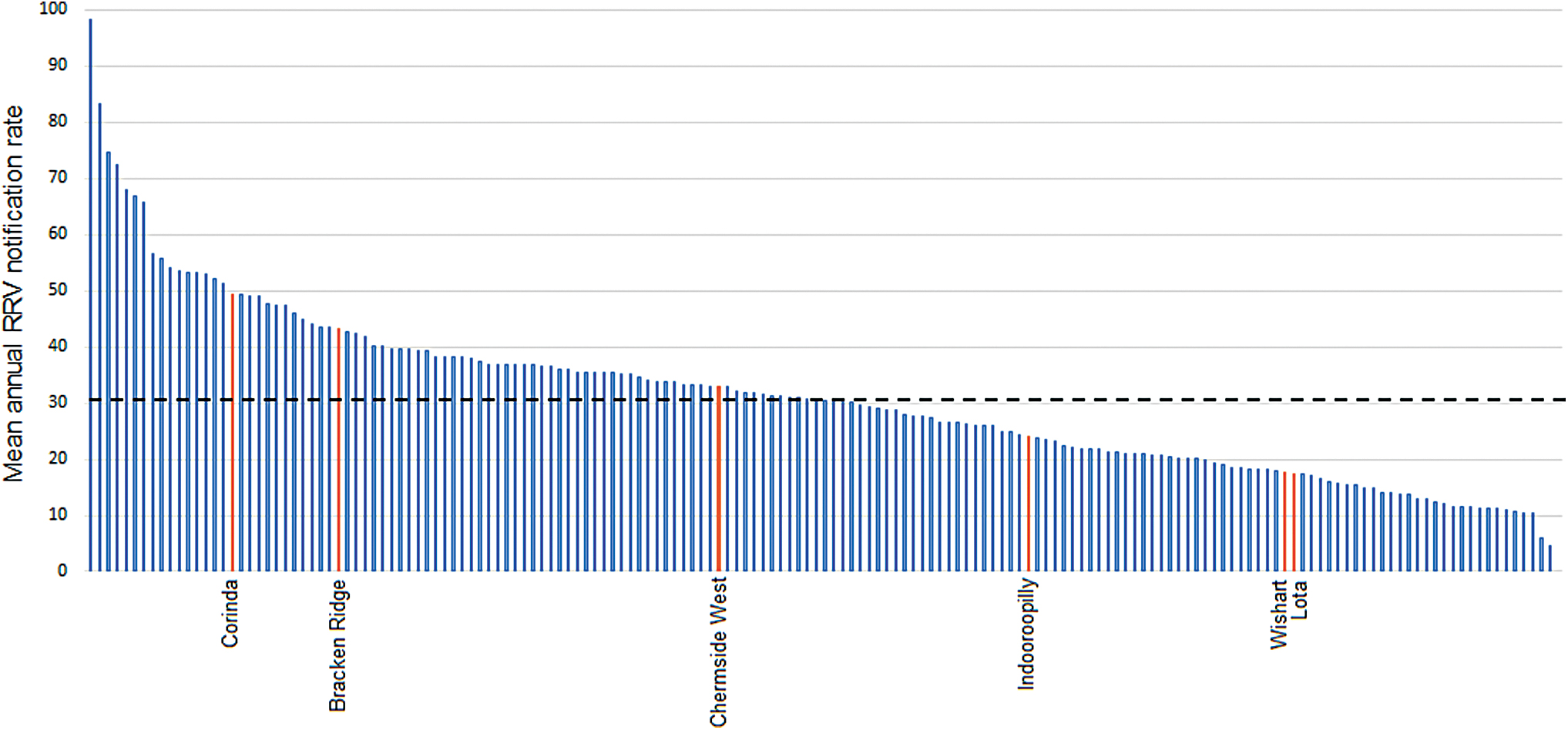
Mean annual notification rate (per 100,000 population) for SSCs in Brisbane with a population greater than 1000. SSCs shown in orange are those within which study sites were located, and the dashed black line represents the average notification rate across all SSCs (31/100,000). Color images are available online.
Vertebrate and mosquito survey summary statistics
Full species lists and counts of individuals observed over the sampling period are summarized in Supplementary Tables S2 and S3. Given the limited number of sites, the power to detect statistically significant results overall is limited. However, of the variables tested, logged vertebrate biomass had a significant association with human notification rate (GLM: R-squared = 0.11, p = 0.03, d.f. = 31). The suburbs reporting the highest human notification rates, Corinda and Bracken Ridge, had the greatest nonhuman vertebrate biomass, which was followed by a decline in biomass in suburbs with lower human notification rates (Fig. 3a).
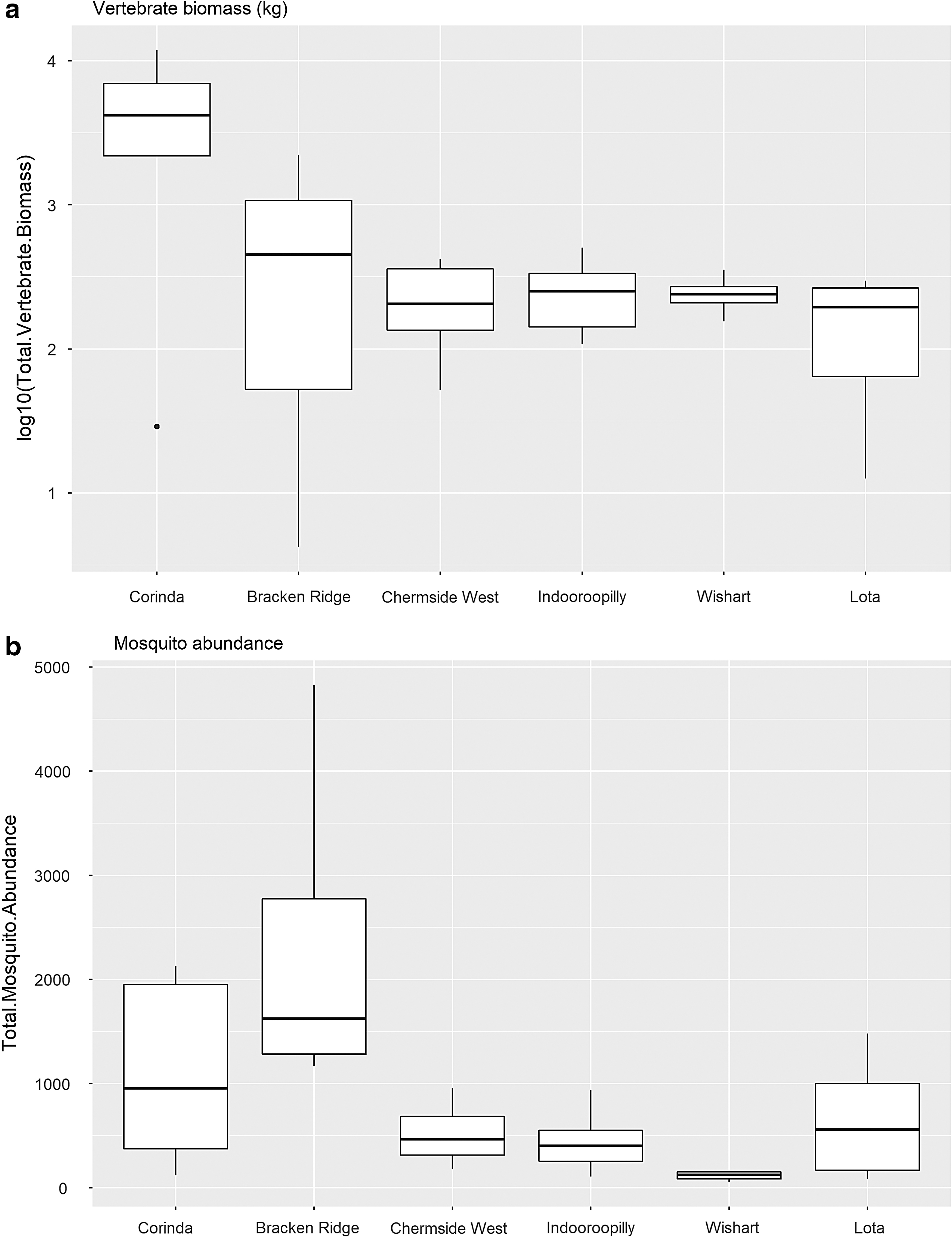
Boxplots of the two variables with a significant association with human cases
Total mosquito abundance was also significantly correlated with human notification rate (GLM: R-squared = 0.2271, p = 0.003, d.f. = 31). However, this trend was not linear, with the Bracken Ridge SSC reporting the highest mosquito abundance and the Wishart SSC reporting the lowest (Fig. 3b). There were statistically significant differences in mosquito diversity, placental mammal abundance, marsupial diversity, and marsupial abundance across suburbs, but these significant differences did not correlate with human notification rates (Supplementary Fig. S1c, d, f, g). No significant differences between SSCs nor associations with human notification rates were observed for vertebrate abundance and diversity, bird abundance and diversity, or placental mammal diversity (Supplementary Table S4, Supplementary Fig. S1a, b, e, h, i).
Community composition
Across all SSCs mosquito collections comprised mostly of the following species: Culex annulirostris, Aedes procax, Aedes notoscriptus, Aedes vigilax, Anopheles annulipes, Coquillettidia linealis, and Culex sitiens (Fig. 4). There was a 29-fold difference in mosquito abundance between the SSC with the highest (Bracken Ridge, n = 13,468) to that of the lowest mosquito abundance (Wishart, n = 456) (Fig. 4b, Supplementary Table S2).
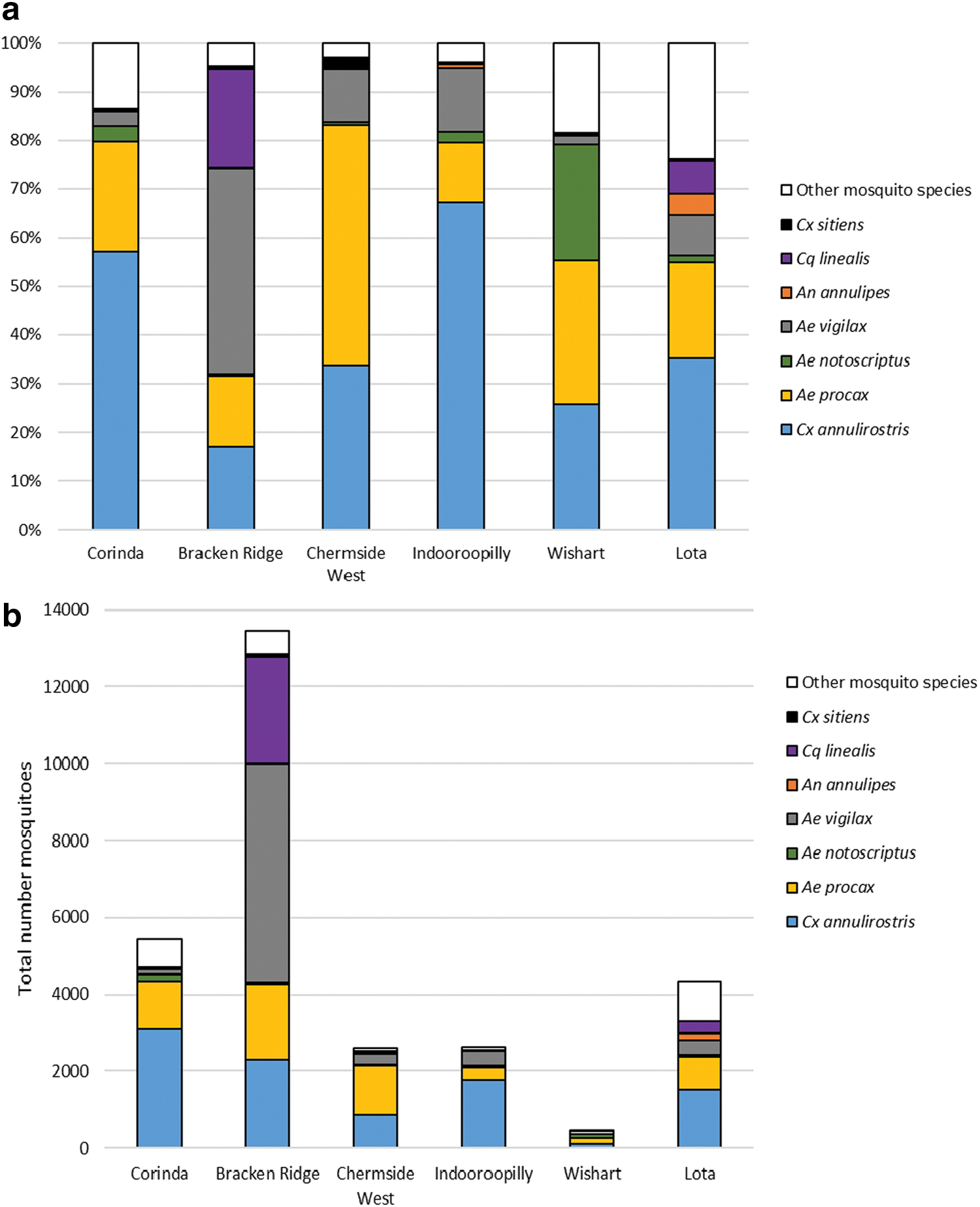
RRV mosquito community composition within each SSC, represented as
SSCs with the highest RRV notification rates were not distinguishable from other SSCs in terms of mosquito species composition. In all SSCs, except Bracken Ridge, both Cx. annulirostris and Ae. procax contributed >55% of the species composition. Compared with other SSCs, Bracken Ridge had a unique species composition where Cx. annulirostris and Ae. procax comprised less than one third, while Ae. vigilax and Cq. linealis dominated (>62% combined). At least 20% of the species composition in Wishart and Lota (SSCs with the lowest RRV notification rates) were species that have not previously been considered as candidate vectors of RRV and, thus, their vector status is unknown (such as Cx. orbostiensis and Ae. vittiger (Harley et al., 2001, Jansen et al., 2019)).
The two SSCs with the highest notification rates were distinguishable from all other SSCs based on their nonhuman vertebrate biomass composition (Fig. 5). Specifically, Corinda and Bracken Ridge had high total vertebrate biomass, almost entirely dominated by Perissodactyla (horses), which was driven by the proximity of horses to a single sampling site. Otherwise, the composition patterns were relatively similar across the remaining four SSCs. Minor differences included a larger proportion of marsupials (Diprotodonts) in Chermside West and Indooroopilly compared to other SSCs and a greater proportion of avian species (including Psittaciformes, Passeriformes, and Pelecaniformes) in SSCs with the lowest human notification rates. Overall, the relative composition of Carnivores was generally consistent across SSCs with mid-range or low human RRV notification rates (Chermside West, Indooroopilly, Wishart, and Lota) ranging from 33.8% in Indooroopilly to 45.8% in Wishart.
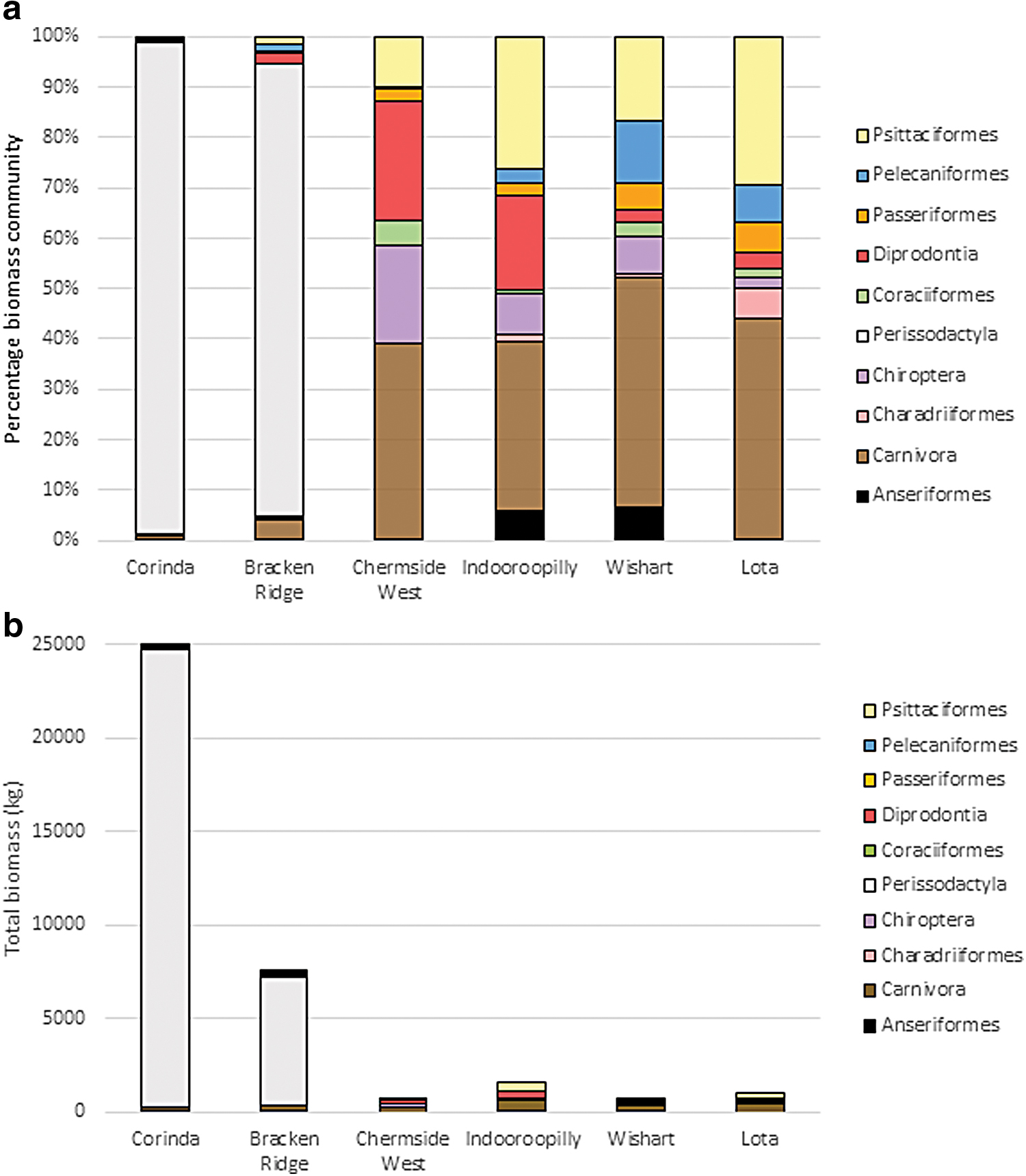
Nonhuman vertebrate community biomass (kg) categorized by taxonomic class within each SSC, represented as
Ordinations
Dissimilarity matrices resulting from NMDS analyses had moderately high stress and demonstrated no clear groupings of suburbs with higher or lower RRV notification rates based on vertebrate biomass or mosquito abundance. In general, community composition values for each survey were grouped most closely together by the SSC they were undertaken in (i.e., Bracken Ridge or Indooroopilly), but showed no trend in grouping between the highest to lowest human notification rate (Fig. 6).
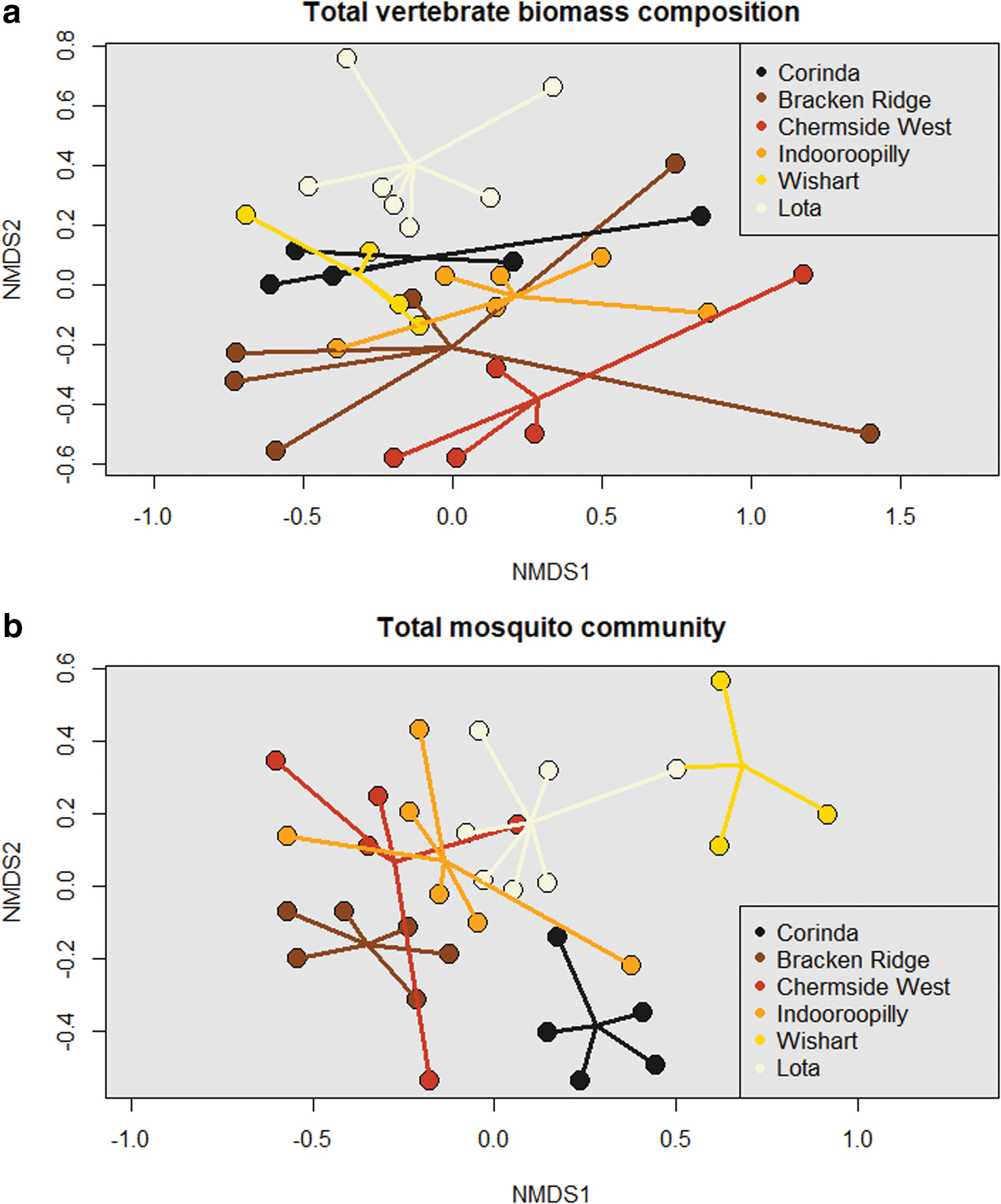
Results of NMDS analysis for the
Discussion
By integrating data on the abundance, diversity, and biomass of potential RRV vertebrate reservoirs and mosquito vectors, we identified associations between high RRV notification rates in humans, total mosquito abundance, and total nonhuman vertebrate biomass (particularly due to the presence of horses).
While variability existed between study sites, the overall positive relationship observed between total mosquito abundance and long-term human notification rates of RRV is as expected for a mosquito-borne virus. This result is also consistent with previous investigations, which found that a greater number of mosquitoes (Lindsay et al., 1996, 1997) or closer proximity to mosquito habitats (Jardine et al., 2015) corresponded with higher human notification rates (Ryan et al., 1999). Importantly, however, this measure of overall mosquito abundance does not necessarily take the vector competence or vectorial capacity of specific species into account. Highly competent species at low abundances could be important for ongoing transmission.
Although RRV has been isolated from more than 40 species of mosquitoes, each differing in general biology, ecology and, most notably, host feeding patterns (Claflin and Webb, 2015, Stephenson et al., 2019b), it is likely that some species are more important than others as enzootic and/or bridge vectors. Further investigations are required to implicate specific mosquitoes as the most important vectors of RRV in Brisbane; however, we observed notable differences in mosquito communities between suburbs. For example, Bracken Ridge had a high dominance of Ae. vigilax and Cq. linealis which is of interest for RRV as these two species have demonstrated relatively high vector competence under laboratory investigations (Ryan et al., 2000, Jeffery et al., 2002) and frequently feed on both humans and horses (which were abundant in this suburb) (Stephenson et al., 2019b).
The dominance of Cq. linealis in Bracken Ridge is also interesting as this species is associated with constructed wetlands in urban environments, which has relevance for authorities managing the increased abundance of this species (Russell, 1999, Crocker et al., 2017, Hanford et al., 2019). The dominance of competent vectors at a given site did not always correspond to high notification rates, for example, more than 50% of mosquitoes trapped at Corinda and Indooroopilly were Cx. annulirostris (a species frequently cited as a vector of RRV), but the two sites have markedly different notification rates. In contrast, Lota, which reported low RRV notification rates, trapped several mosquito species, which have not previously been investigated as vectors of RRV (no virus isolations) or transmission studies; for example, Cx. orbostiensis and Ae. vittiger (Harley et al., 2001). While these species are present in relatively low proportions in these sites, the lack of data on their competence and feeding patterns makes it difficult to determine their contribution to RRV transmission.
The significant positive relationship between vertebrate biomass and RRV notification rate is novel and of interest when mosquito host seeking behavior is considered. Studies on mosquito bloodmeals have found that mosquitoes can feed preferentially on particular vertebrate species, independent of their relative abundance (Kilpatrick et al., 2006, Lyimo and Ferguson, 2009, Janousek et al., 2014). When seeking a bloodmeal, mosquitoes detect hosts through CO2 and body heat (Takken and Verhulst, 2013), in addition to numerous olfactory factors. Both CO2 and heat emission increase with vertebrate body size (Franz et al., 2010), and thus, larger animals may be more attractive to host-seeking mosquitoes.
Biomass accounts for the size of vertebrates, as well as their total abundance. As such, although marsupials are considered potential reservoirs of RRV, their biomass contributed <5% of the community composition in four of the six SSCs. In SSCs with relatively low RRV notification rates, there was a moderate biomass of species that have an unknown or limited reservoir potential, including Carnivores (namely cats and dogs) and Psittaciformes (namely rainbow lorikeets Trichoglossus moluccanus and little corellas Cacatua sanguinea) (Boyd et al., 2001, Stephenson et al., 2018). By comparison, sites with higher notification rates were dominated by a relatively large biomass of horses. Various studies have suggested that horses are competent amplifiers when experimentally infected (developing sufficient viremia to infect vectors) (Kay et al., 1987, Stephenson et al., 2018), that they experience high exposure rates under natural conditions (Gummow et al., 2018, Stephenson et al., 2019a), and they have yielded multiple RRV isolates (Azuolas, 1998). The potential role of horses in the maintenance and transmission of RRV should be explored further, as their high biomass and interactions with vectors may be important for spillover of RRV to human populations.
Overall, our findings suggest that vertebrate biomass rather than abundance alone should be assessed in greater detail for RRV ecology, but it is most informative when combined with existing knowledge on the amplification potential (such as viremic profile or interactions with vectors) of vertebrate species.
The ordinations offered little insight into the disease ecology of RRV. Specifically, no relevant groupings of sites or human notification were ascertained from either the vertebrate or mosquito communities. One possible explanation may be that all sites are situated within the same bioregion (“South Eastern Queensland”), characterized by similar climatic conditions, with the furthest distance between sites ∼30 km. Although it is assumed that most vertebrates and mosquitoes did not disperse widely outside of the sites, flying foxes (Tidemann and Nelson, 2004, Roberts et al., 2012), some bird species (Smith and Smith, 2012), and Ae. vigilax (Chapman et al., 1999, Webb and Russell, 2019) readily travel distances greater than 10 km. In general, while the total community did not vary significantly by SSC within this study, it is unlikely that this would be the case if sites with a greater geographic separation were considered, particularly since RRV has a nationwide distribution, which supports diverse mosquito and vertebrate communities.
Whether a dilution effect exists is an important question when dealing with zoonotic pathogens with multiple reservoir hosts (Rohr et al., 2019). The dilution effect applies to situations in which species diversity reduces measures of disease risk because not all hosts are susceptible and, therefore, act as decoys for vector-borne pathogens (Keesing et al., 2006). This study was not explicitly designed to detect a dilution effect, but if a dilution effect was present, vertebrate diversity would have been greatest in areas with low human notification rates. We did not observe this. Given the complex coupled interactions occurring within this natural system, detecting a dilution effect for RRV would require a large-scale study targeting this specific issue.
Confounding factors within this study include the number of replicates, inherent survey method bias, and the choice of sites based on human notifications. First, the limited number and purposeful selection of sites and survey months mean that our findings may not be applicable across Brisbane or to other cities. Despite confounding factors within this study, this research offers a novel approach to studying zoonotic arboviruses in Australia and provides a framework for other arboviruses such as urban flaviviruses, which may be reliant on urban bird populations for transmission (Maute et al., 2019).
The nature of the data collection, both for mosquito and vertebrate communities, is time consuming and costly. We found that the use of a single mosquito trap in each SSC successfully captured a diversity of mosquito communities that largely corresponded to the habitat conditions (for example, frequent trapping of Ae. vigilax at Bracken Ridge, a saltwater habitat; or frequent trapping of Cx. annulirostris at Corinda and Indooroopilly, freshwater habitats). Future studies, however, would benefit from additional traps. There would be great value for future investigations to expand data collection with additional SSCs across the gradient of human notifications, as well as additional mosquito traps and host surveys.
A caveat of selecting study sites on the basis of human notifications is that it may not accurately represent locations with high or low infection rates of RRV in humans. First, the place of residence of an infected person may not have been the place of infection. In addition, not all cases of RRV cause clinical manifestations, and the proportion of clinical cases that result in notifications is unknown. Finally, it is unknown whether asymptomatic cases in humans contribute to RRV transmission. These factors could result in an underestimate in RRV transmission and notified cases in humans. With these uncertainties in mind, and given the small number of annual notifications within each SSC (n = 0–26), we determined that a long-term notification rate is most suitable to identify and categorize those areas with tendencies for high or low RRV cases.
Future studies on RRV community ecology would benefit from including broader vertebrate survey methods (such as active trapping or passive camera trapping) to incorporate a wider diversity of vertebrates, including day-active species. For example, small and/or cryptic vertebrates, such as murids, were infrequently detected in this study. However, some serological evidence suggests that these species are exposed to RRV (Vale et al., 1991). Although limited in their application, open-access biodiversity datasets (such as the Global Biodiversity Information Facility [Flemons et al., 2007] and the Atlas of Living Australia [Belbin and Williams, 2016]) could be used to cross-check species lists or identify vertebrates in the absence of formal vertebrate surveys.
In addition, inclusion of mosquito bloodmeal analyses would further inform the potential role of the vertebrates considered in this study, by demonstrating which mosquito vectors readily interact with which vertebrate hosts. Of three published blood-feeding studies conducted in Brisbane between 1995 and 2008, birds, humans, placental mammals, and marsupials were all variously dominant among the vertebrate species fed on by mosquitoes, but this was likely influenced by differing mosquito collection sites between the studies (Ryan et al., 1997, Kay et al., 2007, Jansen et al., 2009). Only one of these three studies attempted to quantify the relative availability of host species in trapping sites, finding that, overall, dogs were the most dominant bloodmeal relative to their abundance, but this was based largely on subjective householder survey (rather than vertebrate observation surveys) and was undertaken in SSCs where animals with higher biomass such as horses were not abundant (Kay et al., 2007). This also supports the inclusion of vertebrate and bloodmeal surveys plus consideration of the effect of biomass in future studies.
Conclusion
Improved understanding of both mosquito and vertebrate communities can lead to better prediction and management of transmission for zoonotic mosquito-borne diseases. In this study, we find that RRV vertebrate biomass (particularly horses) and mosquito abundance had the strongest association with human notification rates of RRV in selected suburbs of Brisbane. Further investigations, including ongoing vertebrate surveillance and modeling studies, are needed to quantify the importance of vector competence and mosquito feeding patterns in association with the novel finding of vertebrate biomass from this study.
Footnotes
Acknowledgments
The collection of these data would not have been possible without the following field volunteers: Georgia Braun, Cara Parsons, Ian Parsons, Hannah Thomas, Jack Dodd, Scout Fisher, Stevie Tozer, Stuart MacLeod, Robin Rowland, Jaylan Schabrod, Michael Johnson, and Trina Kateifides. The authors acknowledge Lara Herrero for supervising the PhD study of which this research is a part and more broadly HM laboratory members and Lara Herrero laboratory members for feedback on data collection and analysis.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This project was supported by an Ecological Society of Australia Graduate Grant. E.B.S. was supported by an Australian Government Research Training Program and a Griffith Graduate Research School Publication Assistance Scholarship. A.J.P. was supported by a Queensland Government Accelerate Postdoctoral Research Fellowship.
Supplementary Material
Supplementary Figure S1
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.