
Abstract
Rodents are a large group of mammals that can be carriers of zoonotic pathogens such as Yersinia strains that cause yersiniosis. The prevalence of Yersinia enterocolitica and Yersinia pseudotuberculosis was determined in 214 small wild rodents from south-eastern Poland. Samples were analyzed by precultivation and PCR. Nine (4.2%) Y. enterocolitica and one (0.5%) Y. pseudotuberculosis isolates were received. Most of them (n = 5) were obtained from the common vole (Microtus arvalis). All Y. enterocolitica strains were classified as biotype (BT) 1A. A PCR analysis of virulence markers revealed that all Y. enterocolitica isolates contained the ystB gene and five isolates harbored a rare genetic combination of ail/ystB. Three of the four ail/ystB-positive isolates belonged to serotype O:5.27. The Y. pseudotuberculosis inv-positive isolate was classified as BT 1. A genetic analysis of Y. enterocolitica harboring the ystB gene revealed 100% similarity between the analyzed sequences and the sequences from diarrhea patients in India and the United Kingdom as well as high similarity with the sequences from different species of wild animals from Poland. The Y. pseudotuberculosis inv sequence was 100% identical to the sequence isolated from fully virulent clinical strain from France and Australia. The results of our study suggest that small wild rodents, especially voles and yellow-necked mice, may act as carriers of Yersinia strains. The high similarity of the tested gene sequences between our isolates and the isolates from other free-living animals indicates that small wild rodents can play a role in the epidemiology of yersiniosis and can shed Yersinia spp. into the environment.
Introduction
R
According to the World Health Organization (1982, EFSA 2007), yersiniosis is a zoonotic disease caused by Yersinia enterocolitica and Yersinia pseudotuberculosis. It is the third most prevalent foodborne disease in Europe after campylobacteriosis and salmonellosis. Symptoms of yersiniosis include diarrhea, abdominal pain, fever, and various symptoms of parenteral forms of the disease. According to the European Food Safety Authority (EFSA), 99.1% of yersiniosis cases are caused by Y. enterocolitica, but the most severe form of the disease is caused by Y. pseudotuberculosis (EFSA/ECDC 2018). Symptomatic yersiniosis is caused by serotypes belonging to biotype (BT) 1B, 2, 3, 4, and 5 of Y. enterocolitica, whereas all Y. pseudotuberculosis strains are considered pathogenic for humans and animals (Amphlett 2015).
The pathogenicity of Yersinia is determined by the presence of chromosomal and plasmid genes responsible for disease symptoms and the production of proteins that enable the pathogen to survive in the host body. Yersinia virulence plasmid is unstable; therefore, chromosomal virulence markers are often searched during bacterial detection by PCR. The presence of Y. enterocolitica is most often confirmed based on yst gene, which encodes the production of heat-stable Yst enterotoxins as well as the ail gene (attachment invasion locus) (Fredriksson-Ahomaa et al. 2006, Syczyło et al. 2016). Y. pseudotuberculosis is most frequently identified based on the inv (invasin) gene, which encodes the Inv outer membrane protein—the most important virulence factor promoting adherence to eukaryotic cells and invasion into host cells (Niskanen et al. 2009, Reinhardt et al. 2018).
Y. enterocolitica infections most often result from the consumption of raw or semiraw pork or food products contaminated with the feces of infected animals (Fredriksson-Ahomaa et al. 2001, 2006, EFSA/ECDC, 2018). In humans, yersiniosis caused by Y. pseudotuberculosis is usually as a consequence of consumption of raw vegetables or unboiled water contaminated by infected wild animals (Okwori et al. 2009).
Pigs are the main reservoir and source of Y. enterocolitica strains pathogenic for humans. The prevalence of Y. enterocolitica is high in pig farms due to high levels of contamination originating from the feces of infected animals (Fredriksson-Ahomaa et al. 2001). The experimental attempts to eliminate Yersinia from pigs and breeding facilities were unsuccessful because the pathogen is disseminated via various routes (Gurtler et al. 2005). According to some researchers, field and forest rodents may contribute to the spread of the pathogen in pig farms (Backhans et al. 2011, Backhans and Fellstrom 2012). Field mice were capable of spreading pathogenic microorganisms within a radius of 3 km from an infected farm (Han et al. 2015). The abovementioned implies that rodents can spread Y. enterocolitica to other sites on the farm, to neighboring farms, as well as to the surrounding environment inhabited by free-living animals.
Free-living animals, wild birds, and rodents are regarded as reservoirs of Y. pseudotuberculosis (Fukushima and Gomyoda 1991, Okwori et al. 2009). However, many animal species, including companion animals, livestock, and wild animals, are susceptible to infection.
Yersinia is a psychrophilic microorganism that persists in the environment for a long time. These bacteria were isolated from soil, stagnant water, well water, river water, and from plants contaminated with the feces of infected animals (Terech-Majewska et al. 2016). There is substantial evidence indicating that these microorganisms can circulate between farm animals and free-living animals (Fukushima and Gomyoda 1991, Backhans et al. 2011). The same strains of Y. enterocolitica were found in livestock and game animals (Jakubczak et al. 1993, Platt-Samoraj et al. 2017, Syczyło et al. 2018).
According to various sources, Yersinia pathogens are highly prevalent in field and forest rodents (Kapperud 1975, Backhans et al. 2011). The prevalence of these microorganisms in small rodents has never been investigated in Poland. However, our previous studies have confirmed the widespread occurrence of Y. enterocolitica in wild ruminants, wild boars, and beavers, but the presence of Y. pseudotuberculosis has not been analyzed in the above studies (Jakubczak et al. 1993, Bancerz-Kisiel et al. 2014, 2015, Platt-Samoraj et al. 2015, 2017, Syczyło et al. 2016, 2018).
The prevalence of Y. pseudotuberculosis has been rarely studied in humans, animals, and environmental sources in Europe; therefore, our understanding of the epidemiological mechanisms underpinning yersiniosis is limited. There is also a general scarcity of published data on the role of different mouse species in the epidemiology of yersiniosis. This is the first study to investigate the prevalence of two Yersinia spp. in small wild rodents in Poland.
The aim of this study was to determine the prevalence of and to characterize Y. enterocolitica and Y. pseudotuberculosis in the intestines of small wild rodents in south-eastern Poland with the use of bacteriological and molecular methods.
Materials and Methods
A total of 214 rodents were examined, including the yellow-necked mouse (Apodemus flavicollis), 66 striped field mouse (Apodemus agrarius), 33 common vole (Microtus arvalis), and 1 bank vole (Myodes glareolus), sampled in 6 Polish voivodeships during a 2-year study. The animals were caught with the use of live mouse traps in their habitats and natural foraging areas in the vicinity of farm buildings. The animals were not trapped in protected areas, and protected species were released into the wild. The number of the collected samples and sampling locations are presented in Fig. 1.
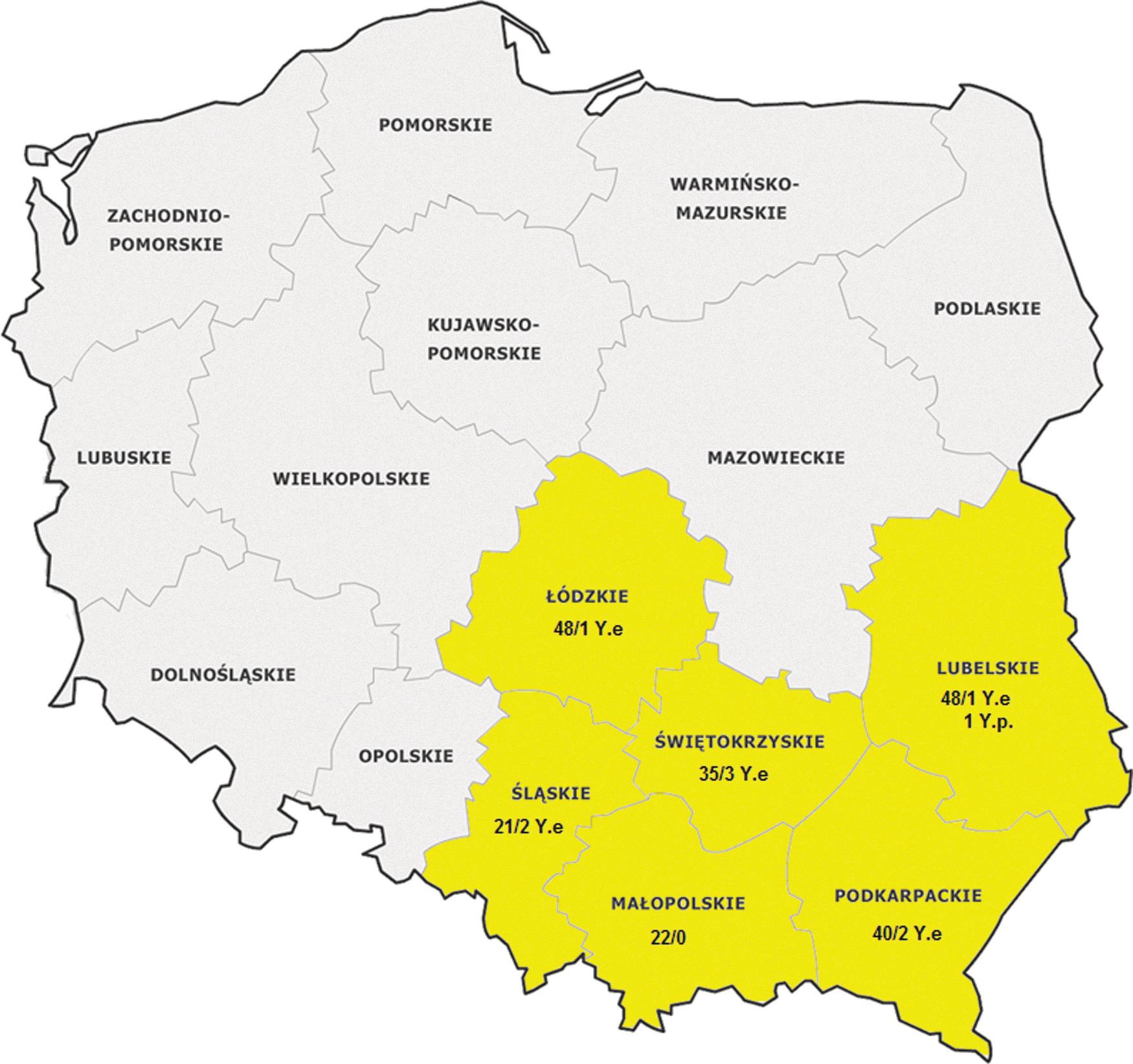
Geographic location, number of Yersinia isolates, and the number of the examined small wild rodents in Poland. The voivodeships where rodents were sampled are highlighted in yellow. Y.e, Yersinia enterocolitica; Y.p, Yersinia pseudotuberculosis.
Rodents were collected as part of a research project on the occurrence of leptospirosis in wild animals, including wild boars, cervids, and red foxes. The animals were euthanized upon the approval of the Local Ethics Committee for Animal Experimentation in Lublin, protocol no. 30/2016. Necropsies were performed on the day of capture. The intestines were aseptically removed and transported to the laboratory in separate sterile containers on ice. In the laboratory, the samples were crushed and vortexed for 5 min with 0.5 mL of phosphate-buffered saline (PBS) (peptone, sorbitol, and bile salts medium prepared according to PN-EN ISO 10273). Then, 100 μL of the supernatant from each sample was enriched with 10 mL PBS and incubated for 21 days at 4°C. Bacteriological and molecular methods were performed as previously described with few modifications (Bancerz-Kisiel et al. 2014). The BT of inv-positive Y. pseudotuberculosis was confirmed in the API 20E test (bioMerieux, raffinose fermentation, melibiose fermentation, and metabolic conversion of citrate). The test strip was incubated at 25°C instead of 37°C for 20–24 h.
Multiplex PCR included the amplification of four gene fragments: ail, ystA, and ystB in Y. enterocolitica with primers described previously (Syczyło et al. 2018), as well as the region of the inv gene specific to Y. pseudotuberculosis with primers described by Thoerner et al. (2003).
Multiplex PCR was carried out with HotStartTaqPlus DNA Polymerase (Qiagen) and the HotStartTaqPlus Master Mix Kit (Qiagen). The following PCR protocol was adopted: initial heating at 95°C for 5 min., followed by 30 cycles of DNA amplification, denaturation at 94°C for 0.75 min, annealing at 54°C for 0.5 min, polymerization at 72°C for 1 min, and final polymerization at 72°C for 10 min. The products were separated by electrophoresis in 2% agarose gel with the Midori Green Advanced DNA stain (Nippon Genetics Europe GmbH, Germany) in 1 × Tris-acetate-EDTA (TAE) buffer. PCR results were analyzed and archived with the use of the GelDoc gel documentation system (Quantity One analysis software; Bio-Rad). The specificity of the reaction was confirmed by sequencing the obtained amplicons.
Nucleotides were sequenced with longer ystB primers (YSTBF: 5′-GGA CAC CGC ACA GCT TAT ATT TT-3′, YSTBR: 5′-GCA CAG GCA GGA TTG CAA CA-3′). The amplicons were purified with the Clean-up Purification Kit (A&A Biotechnology, Poland) according to the manufacturer's recommendations. Purified amplicons were sequenced independently (Genomed S.A., Poland) in both directions. Multiple sequence alignment was carried out in ClustalW (Larkin et al. 2007). A phylogenetic analysis was carried out in Computational Evolutionary Biology MEGA v. 5.2.1. freeware (Tamura et al. 2004). Nucleotide and amino acid sequences were identified using BIOEDIT v.7.2.0 software. The analyzed nucleotide sequences are available in GenBank (MK734422–MK734430).
Results
Y. enterocolitica was detected in nine of the 214 examined rodents (4.2%), yellow-necked mouse (n = 4), and common voles (n = 5). Five isolates, including three isolates from yellow-necked mice and two isolates from common voles, harbored both ail and ystB genes. The Y. pseudotuberculosis isolate was confirmed by PCR in one striped field mouse (0.5%). The results are presented in Table 1.
Characteristics of the Identified Yersinia enterocolitica and Yersinia pseudotuberculosis Isolates
NI, not identified.
All Y. enterocolitica isolates were biotyped as BT1A. Three of the four ail/ystB-positive isolates were serotyped as O:5,27. Seven isolates did not react with any of the available diagnostic sera (not identified). The Y. pseudotuberculosis inv positive was classified as Y. pseudotuberculosis BT1 according to Niskanen et al. (2009) and Tsubokura and Aleksic (1995).
The DNA sequencing of amplicons confirmed the analyzed strains' affinity for Y. enterocolitica and, in one case, to Y. pseudotuberculosis.
Seven of the nine examined ystB sequences from three common voles and four sequences from yellow-necked mice (MK734422–MK734426, MK734428, and K734429) demonstrated 100% similarity to the ystB sequence in Y. enterocolitica isolated from beavers (Acc. No. KJ592624) in northern Poland. Six out of the seven examined ystB sequences (MK734422–MK734426 and MK734428) demonstrated 100% similarity to the partial coding sequences of ystB AY966880 isolated from diarrheic patients in India. Two sequences from common voles showed 100% similarity to the HF571988 strain from human fecal samples isolated in the United Kingdom. Only one ystB sequence (MK734429) from a common vole revealed 100% similarity to Y. enterocolitica DNA for Yersinia Heat-stable Enterotoxin Type B, complete coding DNA sequence (CDS) (acc. no. D88145). In addition, sequences MK734423–MK734426 and MK734428 from four yellow-necked mice and one common vole demonstrated 99.62% similarity to sequence KM253279 derived from deer (Cervus elaphus). ystB sequences MK734430 and MK734427 (from a common vole and a yellow-necked mouse) revealed 99.62% and 99.24% similarity to sequence KU198401 from a mallard duck (Anas platyrhynchos) isolate from Poland. All but one (MK734427) of the examined ystB sequences of Y. enterocolitica were characterized by high mutual similarity (Fig. 2).
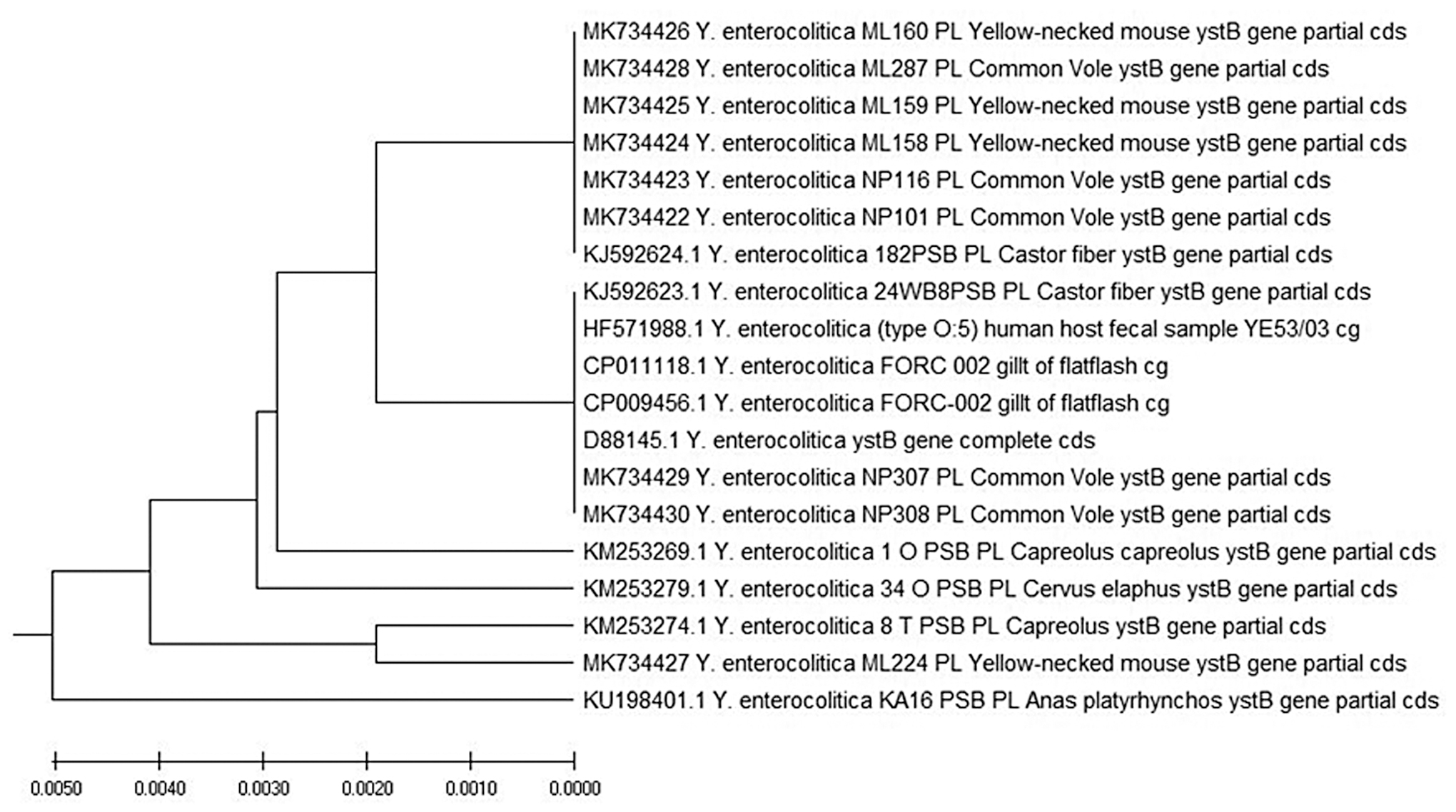
Evolutionary relationships of taxa. The evolutionary history of the examined taxa was inferred with the UPGMA method (Sneath and Sokal 1973). The optimal tree with a total branch length of 0.02387336 is shown. The tree was drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. The evolutionary distances were computed using the Maximum Composite Likelihood method (Tamura et al. 2004), and were presented in terms of the number of base substitutions per site. This analysis involved 19 nucleotide sequences. The analyzed codon positions were 1st +2nd +3rd+noncoding. All ambiguous positions were removed from each sequence pair (pairwise deletion option). There were a total of 263 positions in the final dataset. Evolutionary analyses were conducted in MEGA X (Kumar et al. 2018). cds, coding DNA sequence; cg, complete genome; MEGA X, Molecular Evolutionary Genetics Analysis; PL, Poland; UPGMA, unweighted pair group method with arithmetic mean.
The inv 183-bp sequence from the intestinal sample of yellow striped mouse was 100% identical to the inv sequence (CDS) in Y. pseudotuberculosis serotype I (BX936398.1) that was isolated in France and described as a fully virulent clinical isolate from a human patient. The isolate also was characterized by 100% homology to strain CP010067.1 isolated from a human patient in Japan and 100% identical to LT596221.1 isolated from human feces in Australia.
Discussion
The synanthropization of many rodent species plays a key role in the emergence of many diseases, and it can contribute to environmental pollution. The possibility that Yersinia spp. is spread by rodents has been suggested many years ago. Kaneko et al. (1978) isolated Y. enterocolitica 4/O:3 from a rat caught in a pig house. Kapperud (1975) isolated Y. enterocolitica from the feces of 72.5% of examined samples from rodents.
In our study, Yersinia spp. were detected in 4.7% of the tested animals (n = 9, Y. enterocolitica and n = 1, Y. pseudotuberculosis). Results, similar to ours, were also reported by other researchers. Just like in our study, the vast majority of the isolates were biotyped as BT1A and Y. pseudotuberculosis was isolated from only two shrews (Backhans et al. 2011, Joutsen et al. 2017).
Oda et al. (2015) identified Y. enterocolitica in 15.7% of rodents in Japan. However, their study relied only on bacteriological methods, and the results were not confirmed by molecular techniques. Most Y. enterocolitica strains isolated by Oda et al. (2015) were biotyped as BT1A, but four isolates were classified as BT1B/O:8. The above authors also isolated the same strains from human patients in the province of Aomori, where Y. enterocolitica was isolated from mice (Apodemus argentus and Apodemus speciosus). They concluded that Apodemus mice harboring and excreting Y. enterocolitica serotype O:8 may pollute the environment, including water, and act as a source of infection for humans.
Hayashidani et al. (2003) isolated highly pathogenic Y. enterocolitica bioserotype 1B/O:8 from rodents and pigs in Japan and found similar pulsotypes in rodent and pig isolates, which were indicative of a common source of contamination. These results postulate the presence of epidemiological relationships between Yersinia infections in various animal species and the potential risk to human health.
It should also be noted that the sequences analyzed in our study demonstrated 100% similarity to the sequences isolated from diarrhea patients in India as well as strain HF571988 from fecal samples isolated in the United Kingdom.
The prevalence of Y. enterocolitica and Y. pseudotuberculosis in rodents is closely related to microbial contamination of their habitats, which, among others, results from infection of game animals. In our study, an evolutionary analysis of 19 taxa of the ystB gene revealed high similarity between the sequences from the examined rodents and the previously examined species of wildlife animals from Poland (Fig. 2) (Syczyło et al. 2016, Platt-Samoraj et al. 2017). Sequence identity was determined at 100–99.4% in deer, roe deer, beavers, and wild ducks.
Mutations and natural selection of bacteria in the reservoirs, hosts, and vectors play an important role in the prevalence and incidence of various diseases. These processes can lead to the emergence of new pathogenic and antigenic features, and they can even enable pathogens to adapt to new hosts (Han et al. 2015, Gliński et al. 2017). In the present study, all isolates of Y. enterocolitica were classified as BT1A. However, five isolates harbored ystB as well as ail, which is an important marker of pathogenicity. These isolates resemble classic pathogenic strains that contain ail and ystA genes as well as nonpathogenic BT1A biotypes with only ystB. To date, such configurations have been regarded as extremely rare. Only 11 such strains have been described before 2017. Our previous studies confirmed the high prevalence of ail/ystB strains in game animals (Platt-Samoraj et al. 2017). A comparative analysis of the sequences of ystB fragments confirmed their genetic affinity with the sequences isolated from rodents (Fig. 2).
The low detectability of Y. pseudotuberculosis is surprising, both in our study and in the other researchers' work. The disease caused by Y. pseudotuberculosis is generally referred to as rodentiosis. Published data suggest that rodents are the main reservoir of this pathogen; therefore, a much higher number of isolates were expected in our study. The reasons for the low prevalence of Y. pseudotuberculosis in our experiment and in other studies are complex. According to some authors, Y. pseudotuberculosis is prevalent, but difficult to isolate with the use of the available laboratory techniques and can be easily overlooked. It also can persist in low numbers (Backhans et al. 2011, Psareva et al. 2018).
The epidemiology of infections caused by Y. pseudotuberculosis is still poorly understood. Y. pseudotuberculosis vectors have not been identified to date. Surface water, soil, and plants contaminated by shedding animals can be sources of infection for humans. However, many factors related to the epidemiology of Y. pseudotuberculosis, including sources and transmission routes, are still obscure. There is considerable evidence to indicate that rodents and wild birds act as reservoirs of this pathogen (EFSA 2007, Reinhardt et al. 2018). However, these observations were not confirmed in our study. Y. pseudotuberculosis was isolated from only one out of the 214 tested samples. Such a low isolation rate could be attributed to the complex procedure of culturing the analyzed microorganism. Y. pseudotuberculosis is relatively difficult to isolate. Effective direct and indirect isolation methods are still being sought (Reinhardt et al. 2018). Yersiniosis caused by Y. pseudotuberculosis is sporadically diagnosed in humans. According to EFSA/ECDC (2018), only 0.9% of new cases were reported in Europe in 2017. Such a low detection rate can be attributed to the variety of clinical manifestations of Y. pseudotuberculosis infections, which can obscure the diagnosis, or the fact that microbiological laboratories do not detect Y. pseudotuberculosis during routine testing with the involvement of standard microbiological techniques.
Conclusions
Our results suggest that small wild rodents in Poland, especially voles and yellow-necked mice, may act as carriers of Y. enterocolitica and Y. pseudotuberculosis strains. They can play a role in the epidemiology of yersiniosis and can shed Yersinia spp. into the environment. This conclusion is supported by the high similarity of the tested gene sequences between the obtained isolates and the isolates from other free-living animals. The identified isolates were also characterized by 100% genetic similarity to the clinical isolates deposited in GenBank. Additionally, five Y. enterocolitica isolates with rare ail/ystB genetic configuration have been isolated.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
Project financially cosupported by Minister of Science and Higher Education in the range of the program entitled “Regional Initiative of Excellence” for the years 2019–2022, project no. 010/RID/2018/19, amount of funding 12.000.000 PLN and partially supported by National Science Center, Poland, decision number DEC-2013/09/B/NZ7/02563.