
Abstract
Reservoir-targeted vaccines (RTVs) have the potential to be effective at breaking the transmission cycle of many tick-borne pathogens including, but not limited to, Borrelia burgdorferi, B. miyamotoi, B. mayonii, Babesia microti, and Anaplasma phagocytophilum. To determine what proportion of a wild reservoir species we could effectively target, we distributed an experimental non-RTV Rhodamine B (RhB)-coated pellet formulation devoid of nutrient supplementation using bait boxes with ad libitum access, in battery-operated time-release bait stations, and by hand broadcast. Regardless of distribution method, a total of 208 of 242 (86%) white-footed mouse (Peromyscus leucopus) captures were positive for RhB by either pelage staining or by detecting fluorescent expression in vibrissae under a microscope. In bait box locations, 91% of captured mice were RhB-positive, 89% in hand broadcast locations, and 80% in time-release station locations. Based on results, we are confident that the bait formulation was readily accepted regardless of distribution technique, reached a substantial proportion of the reservoir population, and provides an effective vehicle to deliver a range of RTVs to targeted, wild, pathogen reservoir populations.
Introduction
Zoonotic diseases, infections that exist in animals that can be transmitted to humans, accounted globally for over 60% of all emerging infectious diseases between 1940 and 2004, with over 70% originating in wildlife species (Jones et al. 2008). Zoonotic diseases can be transmitted directly from the animal reservoir to humans, however, 23% of emerging zoonoses are arthropod vector-borne (Jones et al. 2008) with ticks and tick-borne zoonoses becoming an increasingly significant public health concern. For instance, infection with the spirochete Borrelia burgdorferi results in an estimated 300,000 human cases of Lyme disease annually, making it the most common vector-borne disease in the United States (Centers for Disease Control and Prevention 2019a). Cases of human babesiosis and anaplasmosis (caused by infection with Babesia microti and Anaplasma phagocytophilum, respectively) are also increasing in frequency (Eisen et al. 2017). In the United States, the geographic distribution of babesiosis has followed that of Lyme disease (Vannier et al. 2015) and in Connecticut, there was a six-fold increase in the number of cases in the 10 year periods from 1991–2000 to 2001–2010 (Stafford et al. 2014). Reported nationwide cases of anaplasmosis increased 16-fold from 2000 to 2017 (Centers for Disease Control and Prevention 2019b). In addition, new and emerging pathogens are being discovered with greater frequency such as B. miyamotoi and B. mayonii (Gugliotta et al. 2013, Pritt et al. 2016). White-footed mice (Peromyscus leucopus) are a host for immature stages of blacklegged ticks (Ixodes scapularis) and are known competent reservoirs for A. phagocytophilum, B. microti, B. burgdorferi, B. mayonii, B. miyamotoi, and Powassan virus (Anderson et al. 1987, Magnarelli et al. 2006, Barbour et al. 2009, Eisen et al. 2017, Johnson et al. 2017). These disease-causing agents are vectored by I. scapularis in eastern regions of Canada and the United States (Adelson et al. 2004, Dibernardo et al. 2014, Dolan et al. 2016).
Disruption of transmission of vector-borne zoonotic pathogens can be achieved using reservoir-targeted vaccines (RTVs) that can block, or significantly reduce the chance/ability of arthropod vectors to become infected with and transmit disease-causing pathogens to uninfected reservoirs or humans (Bhattacharya et al. 2010, Gomes-Solecki 2014, Richer et al. 2014). Reservoir-targeted vaccines (RTVs) have been successful in preventing B. burgdorferi infection in rodent reservoirs and host-seeking ticks by disrupting transmission cycles (Tsao et al. 2004, Richer et al. 2011, 2014). Current tick-borne RTVs exist only for B. burgdorferi, but future formulations could target other pathogens as well. Borrelia burgdorferi outer surface protein A (OspA) is a proven effective vaccine immunogen (Schoen et al. 1995, Gern et al. 1997, Wormser et al. 1998). Past delivery approaches included hand injection (Tsao et al. 2004), but a highly pragmatic approach is an orally delivered RTV. Oral delivery of an OspA vaccine protected 89% of C3H/HeJ inbred Mus musculus mice against B. burgdorferi infection in a laboratory setting (Gomes-Solecki et al. 2006). A 5-year orally delivered OspA RTV study was field-tested on wild rodent reservoirs in a controlled manner, resulting in a 76% reduction of B. burgdorferi-infected host-seeking nymphal I. scapularis (Richer et al. 2014).
RTVs can be administered using an orally delivered substrate that may be based upon delivery platforms such as liquid sachets or bait pellets coated with encapsulated agents to be released in the digestive tract of the reservoir species (Cliquet et al. 2008, Darkaoui et al. 2014, Gibson et al. 2019). For any orally deliverable substrate to be practical, cost effective, and successful, it must be efficiently distributed across the desired landscape, maintain its structural integrity, withstand the elements, and be readily accepted by targeted reservoir species. Central to the success of any RTV is that the oral bait is required to be readily consumed to deliver a sufficient dosage to a significant proportion of the reservoir population.
A recent effort successfully reduced infection with B. burgdorferi in the wild P. leucopus reservoir and in parasitizing I. scapularis using an RTV (Stafford et al. 2020). However, due to background infection rates, it was difficult to determine the proportion of captured P. leucopus that had been effectively dosed. To determine bait acceptance and the proportion of P. leucopus that consumed bait to ensure effective future RTV delivery, we used Rhodamine B (RhB) dye coated on an experimental formulation of bait pellets that did not contain a vaccine formulation. RhB has been used as a biomarker additive in a variety of wildlife studies including rodents, raccoons (Procyon lotor), feral cats (Felis catus), and coyotes (Canis latrans) (Fisher 1999, Fisher et al. 1999, Fry and Dunbar 2007, Fry et al. 2010). Consumption of RhB ultimately results in a banding pattern in mammalian vibrissae (whiskers) in response to metabolic processing that fluoresces under ultraviolet (UV) light (Fisher 1999). Appearance of RhB bands in vibrissae occurs as the dye is incorporated postingestion and is retained for many weeks. In wild house mice (Mus musculus domesticus) vibrissae for example, RhB bands were readily detected 12 h postingestion and retained for 7–12 weeks (Jacob et al. 2002). Additionally, RhB was used successfully to mark P. lotor in an oral rabies vaccination program (Fry et al. 2009).
We used three different delivery methods to determine effective distribution of an experimental RhB-coated bait formulation at multiple sites in Connecticut, mimicking RTV distribution. Our objectives were to determine bait consumption/acceptance rates of the experimental bait formula and to document the percentage of captured wild P. leucopus that were positive for consumption of RhB comparing the three different distribution methods. Although this study does not report research on the efficacy of a specific vaccine, it does provide important information about an integral component of any oral RTV, delivery and acceptance of bait by the target reservoir species.
Materials and Methods
Study areas
In 2014, we experimented with RhB-coated bait pellets in plastic boxes accessible to small rodents on 22 noncontiguous residential properties in the town of Redding in southwestern Connecticut (41.308°N, 73.392°W). Redding is heavily forested, comprised of relatively large (>1.6 ha) private residential properties adjacent to relatively large, unbroken tracts of water supply lands that are heavily forested. In 2018, we experimented with two additional distribution methods that systematically compared stationary (an experimental time-release bait station) versus ambulatory (hand-distribution) bait broadcasting each on three adjacent 60 × 60 meter plots on a 1400 ha privately owned, forested, water supply property in the town of North Branford in south-central Connecticut (41.350°N, 72.762°W).
RhB-coated bait distribution
Baiting substrate for both 2014 and 2018 was a specialized mouse diet created by LabDiet (Purina, Inc., Largo, FL). Bait was formulated as a pellet devoid of attractants or nutritional supplementation. Powdered RhB (Arcos Organics; Thermo Fisher Scientific, Waltham, MA) was dissolved in 95% ethanol and sprayed onto bait pellets and allowed to dry. Each pellet was coated with 3.75 mg RhB within the context of US Biologic's chemical encapsulation matrix (Zatechka et al. 2017). Once ethanol evaporated and bait was dry, it was stored in a container at room temperature until it was to be distributed.
In 2014 in Redding, RhB-coated bait was distributed via PROTECTA Sidekick® boxes (Bell Laboratories, Inc., Madison, WI) at the 22 properties between August 25 and September 19. Boxes were distributed approximately every 9 meters along lawn-woodland edges (
In North Branford in 2018, we hand-delivered 1000 RhB-coated pellets/60 × 60 meter plot on three occasions at 10-day intervals. At each of the three 60 × 60 meter plots, we walked interior at a 45° angle ∼28 meters from each of the four plot corners and hand broadcast 250 pellets. We followed this procedure on July 9, July 19, and again on July 29, 2018. There was no attempt to quantify consumption of broadcast pellets.
Four time-release bait stations were deployed also in North Branford within each of the three plot interiors, at a 45° angle ∼28 meters from each of the four plot corners. Stations (∼30 cm × 30 cm × 13 cm) were a molded-plastic experimental prototype consisting of six interior chambers engineered to provide iterative feedings to rodent reservoirs to mimic dosing and boosting schedules (Fig. 1). A timed-application locking mechanism made available a predetermined volume of bait pellets via a small aperture located at the bottom of the station. The lid of the bait station locked in place with three screws to prevent nontarget animal consumption and an O-ring sealed out excess moisture from the elements (Luttrell et al. 2018).

Top down view of an opened battery-powered LymeShield bait station filled with uncoated LabDiet bait pellets. Color images are available online.
For this initial field trial, the first three wells contained 125 RhB-coated bait pellets. Stations were affixed to cement paving stones to prevent nontarget wildlife species from relocating them (Fig. 2). Time-release stations were activated on July 13, 2018 and were programmed to rotate on July 23 and again on August 2, 2018. We attempted to quantify bait consumption at all 12 feeding stations both 3 and 7 days into each 10-day rotation by opening time-release stations and counting remaining pellets. After day 7, it was clear there was heavy consumption by P. lotor reaching under devices and advancing carousels to access bait within and confirmed using trail camera imagery. On July 23, we manually filled wells two and three with 125 RhB-coated bait pellets. Thereafter, we permitted bait stations to function as programmed, which limited pellet availability for P. leucopus in the third distribution round due to nontarget consumption.

LymeShield bait station affixed to paver with concrete screws. Color images are available online.
Rodent capture and RhB detection
To determine whether P. leucopus had contacted RhB-coated pellets, they were routinely captured throughout the 2014 and 2018 trials using Sherman live traps (LFAHD folding trap; H.B. Sherman Traps, Inc., Tallahassee, FL) baited with peanut butter. For the 22 residential properties in 2014, rodent trapping occurred during week 2 and 4 between 1 and 3 days post RhB-coated bait distribution. Sherman traps were distributed along the lawn/woodland ecotone at ∼9-meter intervals (
Captured P. leucopus were temporarily sedated using the inhalant anesthetic isoflurane (Piramal Critical Care, Inc., Bethlehem, PA). While sedated, each captured P. leucopus was fitted with a uniquely numbered ear tag (National Band and Tag Co., Newport, KY). Captured and sedated P. leucopus were visually inspected for RhB staining. If no visual evidence was immediately apparent, we placed them in a viewing cabinet with rechargeable six-watt shortwave (254 nm)–longwave (365 nm) combination UV lamp for approximately one minute to detect fluorescing RhB. Additionally, four vibrissae from each sedated P. leucopus were removed and retained. Sedated P. leucopus were returned to their traps, allowed to recover, and then released to the location from which they were originally captured.
Capture and handling protocols for P. leucopus were approved by both the Wildlife Division of the Connecticut Department of Energy and Environmental Protection (#816005a) and the Connecticut Agricultural Experiment Station's Institutional Animal Care and Use Committee (#P18-13) in accordance with the American Society of Mammologists guidelines for the use of wild animals in research (Sikes 2016).
Vibrissa microscopy
All sampled vibrissae were individually mounted on slides and placed on the stage of a fluorescence stereomicroscope (SteREO Lumar.V12; Carl Zeiss, Gottingen, Germany) with a rhodamine filter cube (excitation wavelength = 540 nm; emission wavelength = 625 nm) and observed at 10 × magnification. In 2014, all P. leucopus vibrissae were documented to be either positive or negative for RhB. In 2018, fluorescing RhB bands in vibrissae were each measured to the nearest 0.5 mm from the base of the root to the furthest point where the band was first expressed, which included multiple bands/vibrissa (Fig. 3).
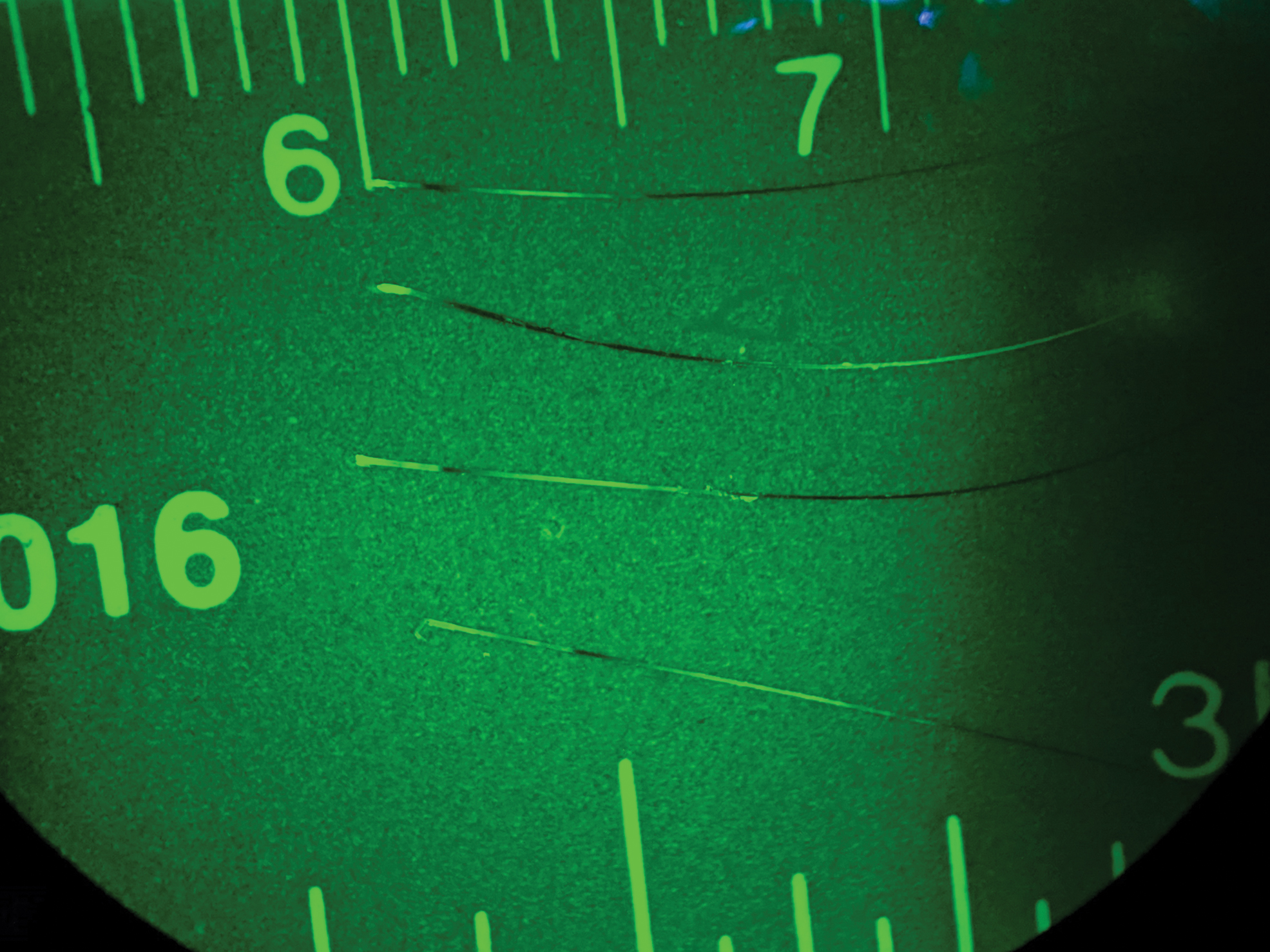
Example of fluorescent banding patterns in vibrissae. The base of each vibrissa (on the left) fluoresced. Fluorescence closer to the base would indicate a more recent encounter with RhB dye as seen in the top vibrissa and fluorescence further from the base would indicate an older contact as in the second vibrissa from the top. Distance between 6 and 7 in the image = 1 cm. RhB, Rhodamine B. Color images are available online.
Development of model for estimated time since bait consumption by P. leucopus
Times that RhB-coated bait was deployed and lengths of fluorescing bands in vibrissae were known. As such, we were able to approximate when individual P. leucopus made contact with bait in the hand broadcast and time-release stations during 2018. We devised a model that incorporated band length, growth rate, and metabolism as known variables to estimate time since consumption:
Length of banding was determined for individual vibrissa and averaged for each captured P. leucopus. Growth rate and metabolism were constants and determined as described below. No references were found for vibrissa growth rate in P. leucopus, however, in laboratory C3H mice growth was ∼0.3–1 mm/day and ∼0.6–1.5 mm/day for rats (Ibrahim and Wright 1975). To approximate vibrissa growth rate in P. leucopus, a linear model was created using all longest band lengths observed compared to the time since bait distribution and fit that to the shortest lengths observed. This model assumed that vibrissae grow at an equal rate which was determined to be 0.75 mm/day, within the range of published results for mice and rats.
Metabolism was defined as the time it took for RhB to first appear as a fluorescing band in vibrissae postingestion. Assuming the metabolism of RhB was equal across all captured wild P. leucopus, the different band lengths observed were fitted in the model using the time since distribution and established growth rate. Metabolism was determined to be 37 h, which is in line with published values in closely related species (Jacob et al. 2002).
Statistical analyses
All data interrogations and statistical analyses were performed using R Statistical Computing Software, version 3.4.4, packages “stats,” “pwr,” “nlme,” “lme4,” “mlbench,” “dplyr,” and “ggplot2” (Leisch and Dimitriadou 2010, Bates et al. 2015, Pinheiro et al. 2020, Wickham et al. 2018). Figures were created using R Statistical Computing Software Statistical Computing Software, version 3.4.4, and GraphPad Prism, version 8.1.2. Fisher's exact test was used to analyze the feeding patterns in P. leucopus and banding patterns in whiskers. Two-sided Fisher's exact test was used to analyze number of feedings in hand versus distribution stations. We used Pearson's chi-squared test for categorical data to determine multiple pairwise differences in the percent of mouse captures positive for RhB contact between the three distribution methods. We compared effectiveness of bait box to hand distribution, hand to time-release station distribution, and time-release station to bait box distribution for captured P. leucopus exposure to RhB for pelage staining, expression in vibrissae, and for captured P. leucopus positive for either pelage, vibrissa staining, or both.
Results
Small mammal capture and bait preference
In 2014, the 22 Redding properties were each trapped on 2 occasions (n = 418 trap nights) resulting in 36 captures in the first trapping round (17% success) and 31 in the second (15% success). Eight P. leucopus captured in the first round were also captured in the second. In 2018, the three hand broadcast plots were each trapped on three occasions (n = 441 trap nights) between 3 and 5 days post-RhB bait distribution. There were a total of 81 captures of 37 unique P. leucopus; 8 were captured once, 14 captured twice, and 15 captured thrice. The three time-release plots were each also trapped on three occasions (n = 441 trap nights) between 4 and 5 days after RhB bait became available. There were a total of 94 captures of 46 unique P. leucopus; 14 were captured once, 16 captured twice, and 16 captured thrice.
Within 2 days of non RhB-coated LabDiet pellet deployment in Redding in 2014, 70% was consumed and after 7 days, 99% was consumed. After 2 days, 85% of RhB-coated LabDiet pellets were consumed and 99% consumed after 7 days in each monitored distribution round. After 2 days of deployment of Rodent Laboratory Chow, 36% had been consumed and 85% consumed after 7 days. In North Branford in 2018, after 3 days of deployment, 64% of RhB-coated LabDiet pellets were consumed from the first available well within time-release stations and 7 days postdeployment, 94% of pellets in the available first well had been consumed, but also 75% of pellets from well two and 77% from well three that were supposed to remain unavailable to rodents. Ten days postdeployment, we refilled 125 RhB-coated pellets in wells two and three and advanced carousels as scheduled. Three days later, 89% of RhB-coated pellets had been consumed from the second available well and 35% consumed from the third unavailable well.
RhB contact
Throughout the three distribution treatments, not all P. leucopus captures positive for RhB pelage staining were positive in vibrissae (Fig. 3) and vice versa. A total of 91% of captures were positive for RhB contact through either pelage staining, vibrissae, or both using the Sidekick boxes in Redding in 2014; 89% were positive using hand distribution and 80% positive for contact using time-release stations in 2018. Further details and total contact with RhB by trapping round can be found in Table 1.
Total Number of Peromyscus leucopus Captures for Each Distribution Type for Each Trapping Round with Number and Percent of Captures Positive for Rhodamine B Exposure on Pelage (Pelage +), Vibrissae (vibrissae +), or Either (Either +)
Fluorescing vibrissae and estimated time since consumption of bait by P. leucopus
In 2018, a total of 135 of the 175 P. leucopus captures (77%) were positive for RhB in vibrissae regardless of distribution method. Fluorescing bands increased in length as days from first deployment of RhB-coated LabDiet pellets increased both in individual vibrissae (Fig. 4A, B) and for each captured P. leucopus (Fig. 4C, D) for both hand distribution and time-release bait stations in 2018. Based on banding patterns, we determined that for each distribution round P. leucopus in the hand distribution treatment had a median of two contacts with RhB while P. leucopus within time-release station plots median was three. In addition, mean length of fluorescing bands in individual vibrissa were significantly longer in time-release station plots than in the hand distribution plots (p = 0.0003; Table 2).
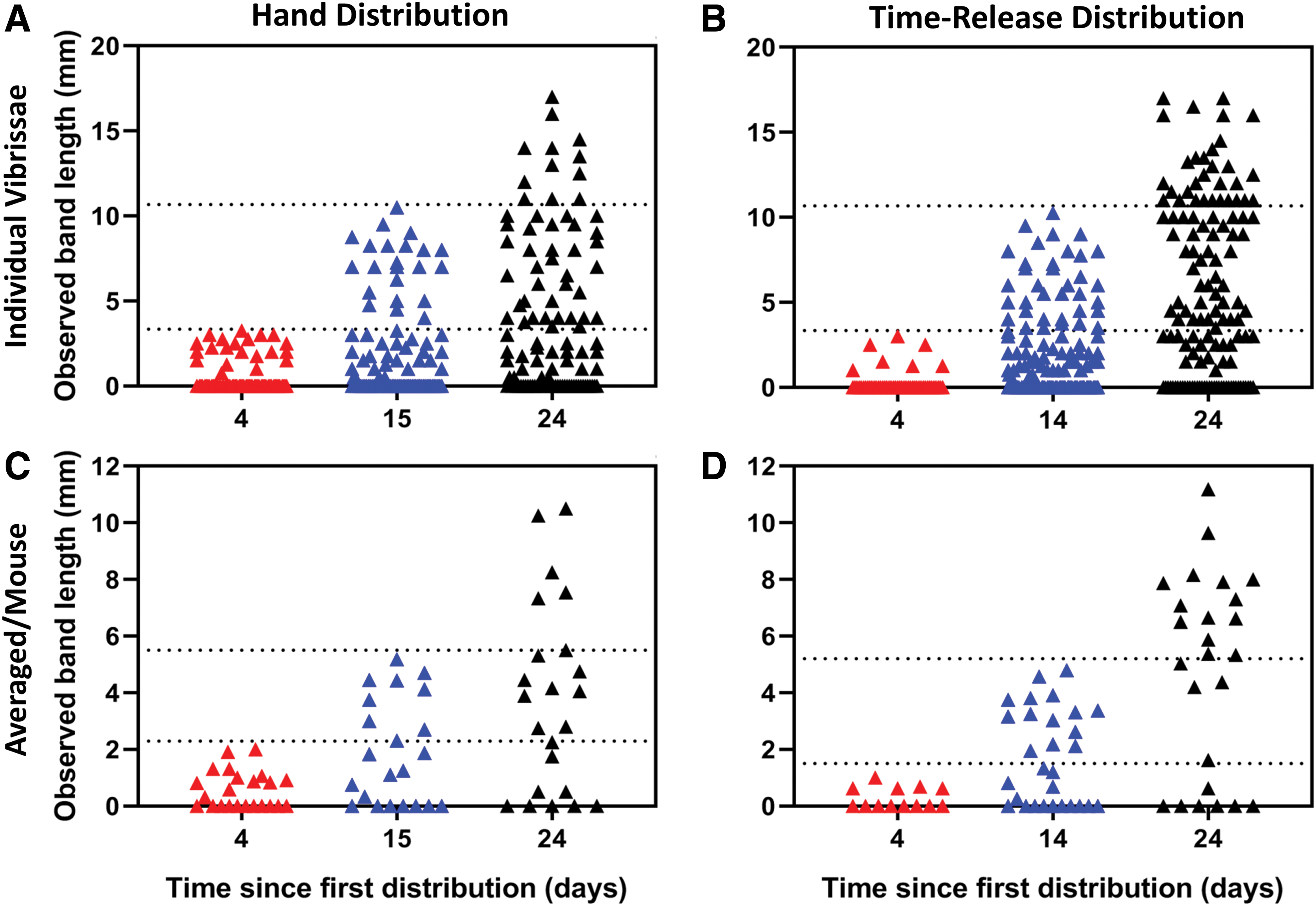
Length of Rhodamine B (RhB) bands observed in individual Peromyscus leucopus vibrissae for hand distribution
Mean Length, Range, and Coefficient of Variation of Fluorescing Bands in Peromyscus leucopus Vibrissae from Hand Distribution and Time-Release Bait Station Locations
Bold values represent statistically significant differences.
CV, coefficient of variation.
Using our model, we then approximated the time elapsed since consumption of bait in the hand broadcast and time-release station plots. At 4 days post time-release station deployment or hand broadcast, as expected, all bands were grouped together as P. leucopus would have had the opportunity to consume bait only within that short 4-day window (Fig. 5). The second trapping round (day 14 bait station, day 15 hand broadcast) revealed a similar distribution of days since consumption (Fig. 5). However, the third trapping round (day 24) showed even distribution in consumption/captured P. leucopus in the hand broadcast plots (Fig. 5C) but a lag in more recent encounters with RhB in the time-release stations (Fig. 5D).
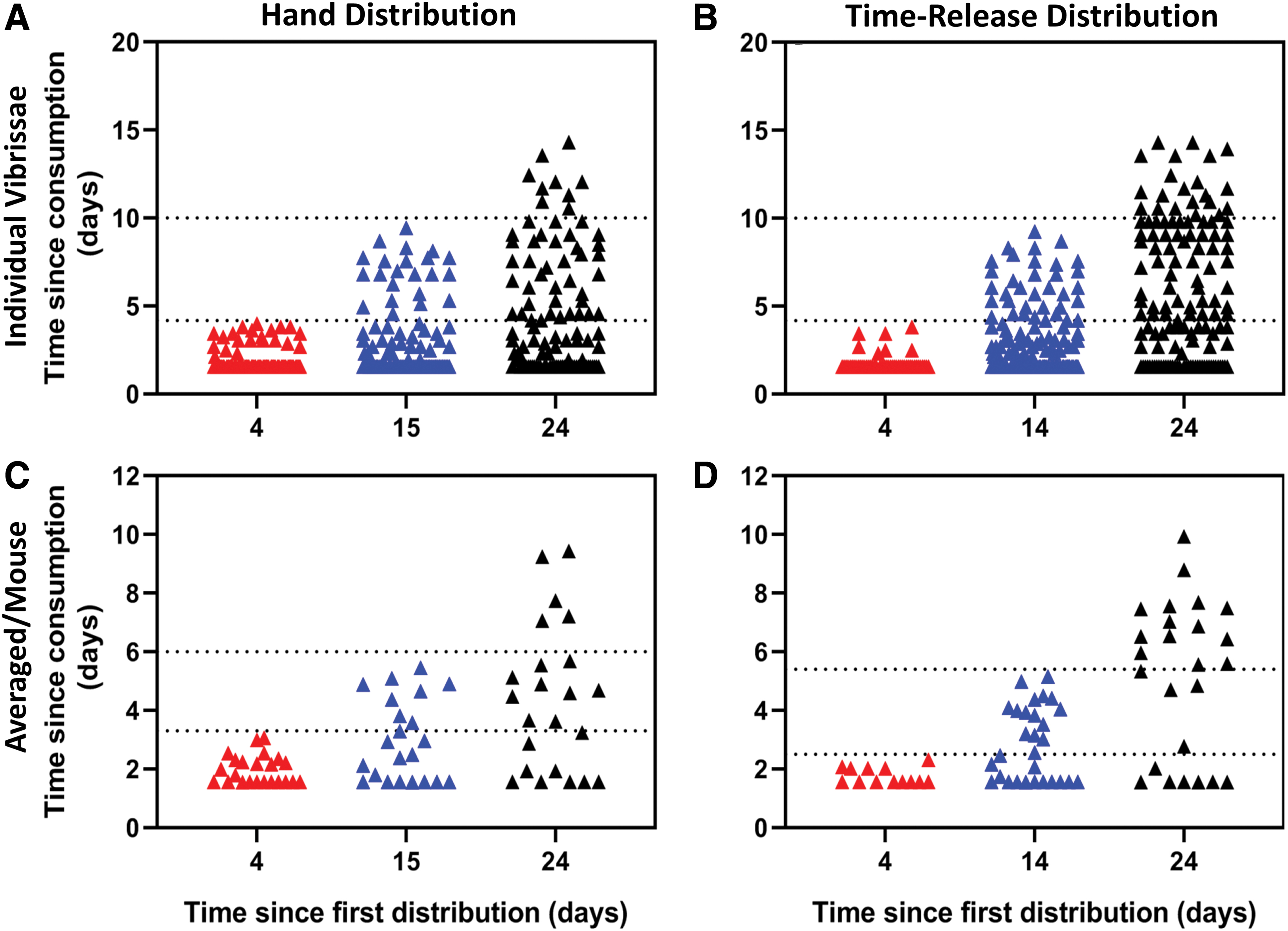
Estimated time since bait consumption by captured Peromyscus leucopus by individual vibrissae for hand distribution
Statistical analyses
Each captured P. leucopus was determined to be positive for contact with RhB through pelage staining, fluorescing bands in vibrissae, or both. There were no statistical differences in comparison of the bait box and hand broadcast distribution in pelage staining, vibrissae, or the proportion of P. leucopus positive for RhB presence by either means. However, a significantly higher proportion of P. leucopus were positive for contact with RhB using hand broadcast than time-release stations for both pelage staining (p < 0.001) and vibrissae (p < 0.03), but there was no significant difference for RhB contact with either. Similarly, a significantly higher proportion of P. leucopus were positive for RhB when comparing bait box to time-release station distribution for pelage staining (p < 0.001), vibrissae (p < 0.03), and either one (p = 0.03). Detailed results can be found in Table 3.
Multiple Comparison Analysis of Positive Rhodamine B Contact by Captured Peromyscus leucopus by Distribution Method
Bold values represent statistically significant differences.
Discussion
Over the past century, two trends have been observed in global infectious diseases; the first half experienced a decline in the spread and incidence of infectious diseases due to the advent of population immunization strategies and increased awareness and implementation of sanitary methods (Lederberg et al. 1992, Cunningham et al. 2017). During the second half, an increase was observed due to newly emerging pathogens as the result of increases in global travel, urbanization, and climate change (Jones et al. 2008). Of the emerging infectious diseases, most are classified as zoonoses, the majority of which are found in wildlife (as opposed to domesticated) reservoirs (Taylor et al. 2001, Woolhouse and Gowtage-Sequeria 2005, Jones et al. 2008). Because host reservoirs can be infected with multiple pathogens and their numbers are orders of magnitude smaller when compared to the abundance of arthropod vectors, RTVs have the potential to be effective at preventing infection, breaking pathogen transmission cycles, and ultimately, reducing diseases in humans.
Over the past three decades, western and central Europe have implemented oral vaccination strategies and successfully controlled the spread of rabies in red fox (Vulpes vulpes) populations (Freuling et al. 2013). A similar effort was recently implemented in the United States using manual and/or aerial distribution of rabies vaccine oral bait (Maki et al. 2017). As bait was broadcast unprotected, challenges were identified including bait consumption by nontarget animals and scarcity of bait in densely populated areas (Henning et al. 2017). RTV formulations targeting B. burgdorferi were shown to be effective in laboratory (Gomes-Solecki et al. 2006, Kern et al. 2016) and controlled field settings (Richer et al. 2014), but many variables exist that can limit effectiveness when broadcast across the landscape. However, Stafford et al. (2020) recently showed that this distribution technique can be effective as well. To complement the results by Stafford et al. (2020), we feel it essential to certify bait acceptance by the target reservoir despite alternate food availability.
An effective bait as an RTV delivery vehicle must meet several criteria including palatability, stability, be devoid of nutrients to avoid increasing reservoir population fecundity, and compatibility with the vaccine formulation to produce an appropriate immune response (Zatechka 2016). Our consumption data using RhB show that P. leucopus readily consumed the LabDiet bait, suggesting excellent palatability combined with the chemistry employed in the bait formulation, which ensured long-term stability, particularly in the hand distribution plots where pellets were exposed to the elements. Wild P. leucopus were not deterred by the RhB coating on the LabDiet pellets and had the same consumption results as uncoated LabDiet pellets, 99% consumption after 7 days deployed. These results are in accordance with previous studies, which demonstrated that rodent pellets with a surface coating of RhB were highly palatable to P. leucopus and did not affect bait acceptance (Fisher 1999, Jacob et al. 2002).
Because of the inevitable consumption of bait by nontarget wildlife (Haley et al. 2019), we assumed that rodent reservoirs would have a greater likelihood of encountering pellets that were hand distributed on the forest floor than learning the location of and having to compete for pellets at a point source like a bait box or time-release station. We confirmed our assumptions by the fact that 72 of the 81 mouse captures (89%) over the three trapping rounds at the hand distribution plots were positive for RhB contact. Additionally, the percentage of captures positive for RhB in capture round 1 were 100% in the hand distribution plots as opposed to 65% in the time-release station plots. This was to be expected and is a function of the learned behavior of P. leucopus, which slightly delayed their initial contact with coated bait (Pyter et al. 2006). Individuals need to encounter the bait station and learn to return to it to access bait within; a targeted or nontargeted reservoir host is much more likely to have a chance encounter with bait broadcast over a relatively large area. This learned behavior was evidenced further in the second round of trapping at the time-release bait station plots where 31 of 32 captured P. leucopus (97%) were RhB positive. Clearly, P. leucopus (and P. lotor) in the vicinity learned of the presence of bait in the stations through trapping round one, continued to check periodically, and when bait became available again, readily exploited it. Additionally, the fact that mean lengths of RhB bands in vibrissae were significantly longer in P. leucopus from the time-release bait station plots is further evidence of this learned behavior and resulting in long-term contact with coated bait. Time-release station manipulation and resulting bait consumption by the nontarget P. lotor reservoir significantly reduced the amount of bait available for the third distribution round, leading to the lag of new RhB contact that can be seen in Figs. 4D and 5D.
Iterative feeding behavior is especially important when considering the distribution of RTVs, where often a primary and at least one booster immunogen exposure are required to establish lasting immunity (Ramshaw and Ramsay 2000). The time-release bait stations were engineered to deliver such a feeding schedule to the rodent reservoir on the landscape while also sheltering remaining bait from the elements. Additionally, the time-release stations were designed to be accessible only to wild P. leucopus and not to larger, nontarget reservoirs such as coyotes, wild turkeys (Meleagris gallopavo), American crows (Corvus brachyrhynchos), and foxes that could consume multiple pellets encountered on the forest floor. However, it continually proves difficult to thwart the strength, dexterity, and intelligence of P. lotor. Hand broadcast bait would be subject to weather and consumption by an entire array of nontarget species that could limit its availability to target reservoir species in a matter of days postdeployment. The fact that bait density in time-release stations was half what it was at hand distribution plots coupled with heavy interference from P. lotor explains the reduction in contact with RhB in the third distribution round, though still 80% of captured P. leucopus were RhB positive.
For RhB to be detectable in vibrissae using fluorescence microscopy, P. leucopus require a minimum dose of 73.3 mg/kg, and for a 25 g individual, which equates to ∼1.8 mg RhB (Weerakoon et al. 2013). Because each pellet was coated with 3.75 mg RhB, consumption of only half a pellet would have resulted in successful detection of fluorescent banding in vibrissae. The vast majority of captures encountered RhB multiple times over each of the three rounds, which, in an actual vaccination program, could result in successful priming and boosting exposure to the applied RTV.
Conclusions
Regardless of which distribution technique was used, we documented the vast majority of P. leucopus captures (86%) were positive for contact with RhB on at least one occasion. Vibrissa staining was a more reliable source of contact with RhB and through the presence of multiple fluorescing bands could determine multiple contacts, but may have missed more recent encounters that had not yet been metabolized at the time of capture. We also determined that the learned behavior of P. leucopus readily returning to a point food source such as the time-release stations would likely result in an effective boosting and priming sequence when distributing an RTV. We have demonstrated that the new oral bait formulation is stable, well accepted by P. leucopus, and even suitable for broadcast in the environment, an essential step in the future distribution of an RTV bait formulated with an OspA or other types of vaccines.
Footnotes
Acknowledgments
We would like to thank the property owners in Redding, CT, and the South Central Connecticut Regional Water Authority for allowing us access to their properties. We would also like to acknowledge Michael Short, Heidi Stuber, Laura Estep Hayes, Steph Shea, Heather Wiles, Sarah Hemstock, and Jamie Cantoni for their assistance with bait distribution and rodent trapping and Lars Eisen for reviewing an earlier version of this article.
Disclaimer
Its contents are solely the responsibility of the authors and do not necessarily represent the official views of the Centers for Disease Control and Prevention or the Department of Health and Human Services.
Author Disclosure Statement
S.Z., J.G.v.O., L.M.R., S.K.W., and C.P. are employed by U.S. Biologic, Inc. The authors declare this affiliation did not create a conflict of interest in preparation of this document. The other authors declare no conflicts of interest.
Funding Information
This research was supported by U.S. Biologic, Inc., Hatch Act funds, and Centers for Disease Control and Prevention Cooperative agreement number U01CK000509.