
Abstract
Although rodents are well-known reservoirs and vectors for a number of zoonoses, the functional role that peridomestic rodents serve in the amplification and transmission of foodborne pathogens is likely underappreciated. Clear links have been identified between commensal rodents and outbreaks of foodborne pathogens throughout Europe and Asia; however, comparatively little research has been devoted to studying this relationship in the United States. In particular, regional studies focused on specific rodent species and their foodborne pathogen reservoir status across the diverse agricultural landscapes of the United States are lacking. We posit that both native and invasive species of rodents associated with food-production pipelines are likely sources of seasonal outbreaks of foodborne pathogens throughout the United States. In this study, we review the evidence that identifies peridomestic rodents as reservoirs for foodborne pathogens, and we call for novel research focused on the metagenomic communities residing at the rodent-agriculture interface. Such data will likely result in the identification of new reservoirs for foodborne pathogens and species-specific demographic traits that might underlie seasonal enteric disease outbreaks. Moreover, we anticipate that a One Health metagenomic research approach will result in the discovery of new strains of zoonotic pathogens circulating in peridomestic rodents. Data resulting from such research efforts would directly inform and improve upon biosecurity efforts, ultimately serving to protect our food supply.
Introduction
Foodborne illnesses are a major threat to human health, and it is estimated that foodborne pathogens sicken at least 48 million in the United States annually, causing upwards of $70 billion USD in health-related costs (Scallan et al. 2011, Scharff 2012, Hoffman et al. 2015, Hoffmann and Scallan 2017). Moreover, ∼64% of hospitalization and deaths associated with foodborne pathogens in the United States are caused by zoonotic bacteria, including nontyphoidal Salmonella enterica, Campylobacter spp., Clostridium spp., Shiga-toxin producing Escherichia coli (STEC), and Listeria monocytogenes (Scallan et al. 2011).
The direct environmental point source for many of the zoonotic enteric pathogens that enter our food supply is typically difficult to determine; however, wildlife distributed throughout local environments associated with a given outbreak are frequently implicated in spreading zoonosis (Atwill et al. 2012, Himsworth et al. 2013, Greig et al. 2015, Sellers et al. 2018, Ayyal et al. 2019). When considering putative reservoirs and vectors for the major zoonotic foodborne pathogens that underlie human disease, a critically important and uniting feature is that each is associated with peridomestic rodent hosts (Backhans et al. 2011, Himsworth et al. 2015, Ayyal et al. 2019, Bondo et al. 2019, Tan et al. 2019) (Table 1).
Zoonotic Foodborne Pathogens Associated with Rodent Reservoirs
AMR, antimicrobial resistance; STEC, Shiga-toxin producing E. coli.
Rodents are the most specious group of mammals in the world with over 2000 species recognized, and they are well known for harboring a plethora of zoonotic pathogens of human health concern (e.g., Hantaviruses, Lassa fever virus, Giardia, Borrelia, and so on) (Han et al. 2015). Commensal and peridomestic rodent species are of special interest to the global One Health initiative, as regionally invasive and native species of mice and rats have benefitted from human activities, especially agricultural systems. Commensal rodents are a common component of the agricultural landscape, and these animals are known to transmit foodborne pathogens to livestock, poultry, and raw produce by contaminating the overall farm environment (Rodriguez et al. 2006, Meerburg 2010, Backhans and Fellström 2012, Kilonzo et al. 2013). This transmission is largely due to the amplification of foodborne pathogens through the daily deposition of urine and fecal pellets into the production environment, and a clear example of this rodent-based amplification is found with Salmonella (Fig. 1).

Salmonella amplification by a rodent host.
Fifteen Salmonella Enteritidis cells can successfully infect a mouse (Mus musculus), and the infection can be maintained for over 10 months (Trampel et al. 2014). During active shedding, one mouse fecal pellet can contain up to 230,000 Salmonella cells, and the rodent can output more than 100 fecal pellets in a day (Fig. 1). Thus, a single rodent within a barn or food-production facility can introduce upwards of 23 million Salmonella bacteria into production pipelines within 24 h (Davies and Wray 1995, Trampel et al. 2014). In particular, the poultry industry is especially susceptible to rodent-driven Salmonella outbreaks as poultry preferentially consume rodent fecal pellets (see Salmonella section below), and as little as 100 Salmonella bacteria are required to successfully infect young chicks (sustained doses of 106 colony-forming units [CFUs] required for maintaining infections [Van Immerseel et al. 2004]).
Despite a growing list of examples that document strong links between rodents on farms and outbreaks of foodborne pathogens in Europe and Asia (Berndtson et al. 1996, Burt et al. 2012, Lapuz et al. 2012, Espinosa et al. 2018, Camba et al. 2020), comparatively little research on the rodent-agriculture interface has occurred in the United States (Henzler and Opitz 1992, Kilonzo et al. 2013). Moreover, studies investigating this interface frequently lack specific knowledge of the rodent species involved and refer to rodents simply as “mice” or “rats,” therefore preventing the identification of species-specific patterns with respect to reservoir status, demographic trends, etc. (Hancock et al. 1998, Hiett et al. 2002). For these reasons, the role of peridomestic rodents as reservoirs or vectors of zoonotic foodborne pathogens in the United States is not well defined, and there is a great need to fill this epidemiological knowledge gap.
We review select rodent species and bacterial foodborne pathogens that are routinely linked to rodent reservoirs. Our intent is to bring awareness to the rodent species diversity that must be considered when examining environmental sources of foodborne pathogens; these data can ultimately inform public health policies and species-specific biosecurity measures in food production operations.
Primary Species of the Rodent-Agriculture Interface Across the Midwestern United States
In this study, we summarize some of the rodent species that are likely at the heart of the rodent-agriculture interface in the Midwestern United States (i.e., Iowa, Michigan, Minnesota, Missouri, Nebraska, North Dakota, Ohio, South Dakota, and Wisconsin), one of the most agriculturally productive regions of the world. Importantly, the species we highlight represent a regional example and are not exhaustive; these are species that are commonly observed on farms throughout the Midwest and they include both North American native species and invasive species.
Native Species
White-footed mouse
The White-footed Mouse (Peromyscus leucopus) is a relatively small rodent in the genus Peromyscus that has a distribution extending from Canada to Mexico, including the eastern and midwestern United States, as far west as Arizona (Fig. 2) (Lackey et al. 1985). White-footed mice are semiarboreal, but also occur in agricultural areas (Drickamer 1970, Cummings and Vessey 1994); and their diet consists of seeds, green vegetation, insects, and arthropods (Lackey et al. 1985, Wolff et al. 1985). Cummings and Vessey (1994) reported that P. leucopus could be found in nearly all areas trapped across the farmsteads of Ohio and was second in abundance only to the invasive house mouse (M. musculus; see below). White-footed mice are of special concern to One Health because they have been positively identified as vectors for not only hantavirus (Cummings and Vessey 1994) but also for other pathogens causing Lyme disease, human babesiosis, and human granulocytic ehrlichiosis (Stafford et al. 1999). When considering foodborne zoonoses, multiple pathogens have been identified within Peromyscus hosts (Table 1).
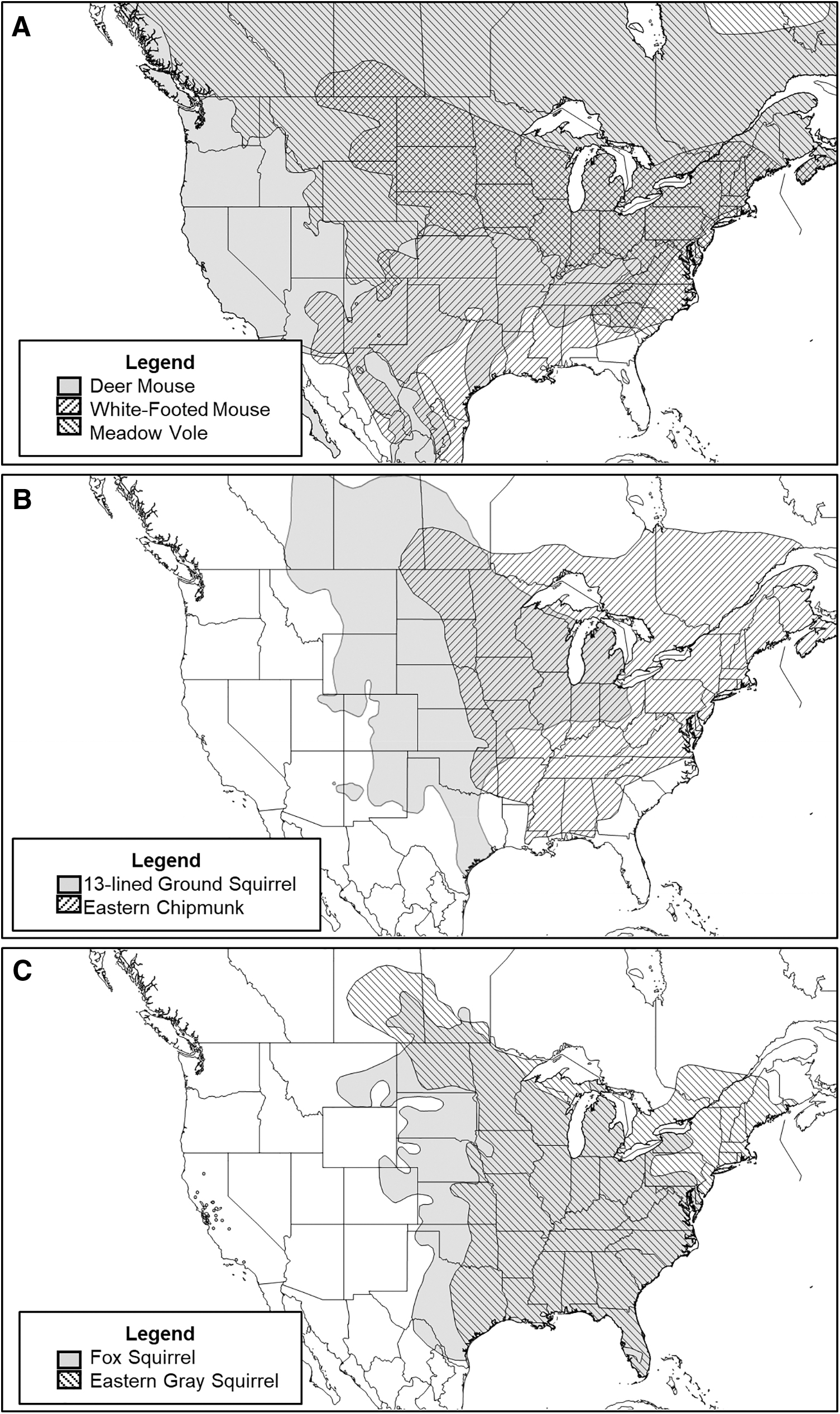
Geographic distributions of native rodent species highlighted herein.
Deer mouse
A close relative to the White-footed Mouse is the Deer Mouse (Peromyscus maniculatus) from the same genus, Peromyscus. The Deer Mouse is a common rodent species that is distributed throughout North America, including Canada, nearly all the United States, and southward into Mexico (American Society of Mammalogists and King 1968). Similar to the White-footed mouse, deer mice feed on seeds, vegetation, and arthropods (Jameson 1952, Wolff et al. 1985). Deer mice are well adapted to different environments, including grasslands, prairies, and agricultural lands (Beckwith 1954), and have been reported to prefer indoor granaries when living in a farm setting, especially during winter months (Bovet 1970). Peromyscus maniculatus is a known reservoir for many zoonotic pathogens, including Sin Nombre Virus (Madhav et al. 2007), Cryptosporidium and Giardia (Kilonzo et al. 2017), and Coxiella burnetii and Pasteurella pestis (Yersinia pestis) (Orsborn et al. 1959). This species is also associated with several foodborne pathogens (Table 1).
Meadow vole
The Meadow Vole (Microtus pennsylvanicus) can be distinguished from Peromyscus species by a darker pelage and shorter tail (Reich 1981). It has a large range compared to other voles in the genus Microtus. The Meadow Vole occurs throughout Canada, the north and eastern parts of the United States, and extending southward into Mexico (Reich 1981). This species has been known to frequent farm settings in the Midwest and is associated with dense grassland habitats (American Society of Mammalogists and Tamarin 1985). Meadow voles are mainly herbivores, although they have been found to eat insects and scavenge on animal remains (Zimmerman 1965, Reich 1981). Microtus pennsylvanicus has been implicated as a reservoir for Borrelia burgdorferi (causative agent of Lyme disease) (Markowski et al. 1998) and Rickettsia responsible for Rocky Mountain Spotted fever (Gould and Miesse 1954). Multiple species of voles (Microtus) have been implicated as reservoirs for Campylobacter sp., E. coli, and Listeria sp. (Table 1).
Thirteen-lined ground squirrel
The Thirteen-lined Ground Squirrel (Ictidomys tridecemlineatus) is recognizable by its slender body and distinctly alternating light-colored longitudinal stripes and rows of spots (Lawlor 1982). The distribution for the thirteen-lined ground squirrel extends from Canada to Texas, east of the Rocky Mountains to the Great Lakes region (Streubel and Fitzgerald 1978). This species is considered carnivorous, mainly feeding on grasshoppers, although seeds and other vegetation have been found in their stomach contents (Fitzpatrick 1925, Whitaker 1972). Thirteen-lined ground squirrels have adapted well to farms and urban areas (Streubel and Fitzgerald 1978), thus increasing the risk of zoonotic disease transfer. For instance, Cloud-Hansen et al. (2007) reported multidrug resistant bacteria from wild-caught thirteen-lined ground squirrels (Morganella morganii and Stenotrophomonas maltophilia). Furthermore, thirteen-lined ground squirrels have been implicated as potential transmitters of avian influenza viruses (Vandalen et al. 2009). Very little research has focused on the putative pathogens that thirteen-lined ground squirrels carry despite the species being a common resident on farms and in urban areas throughout the Midwest.
Eastern chipmunk
The Eastern Chipmunk (Tamias striatus) is heavier-set than the thirteen-lined ground squirrel and has prominent white dorsal stripes flanked by darker stripes and no spots (as seen in the thirteen-lined ground squirrel). The Eastern Chipmunk is distributed throughout eastern North America, including Canada and southward to the Gulf states, but is absent from the coastal plain (Snyder 1982). Eastern chipmunks are omnivorous, and their diet includes seeds, nuts, insects, fungi, frogs, snakes, birds, and small mammals (Elliott 1978, Snyder 1982). Tamias striatus has been associated with West Nile Virus (Gómez et al. 2008) and was implicated as an amplifier host for La Crosse Virus (Hanson et al. 1975). Furthermore, isolates of E. coli have been found in chipmunks (Maldonado et al. 2005). Other datasets showing putative links to foodborne pathogens are lacking.
Eastern gray and fox squirrels
The Eastern Gray Squirrel (Sciurus carolinensis) is distributed throughout the eastern United States and northward into Canada. Several introductions have occurred in the western United States and Europe (Koprowski 1994a). The Fox Squirrel's (Sciurus niger) range overlaps with much of the Eastern Gray Squirrel's range, but extends further west in the United States and Canada (Koprowski 1994b). The Eastern Gray Squirrel is distinguishable from the Fox Squirrel by its silver/white hair on the ventral side (belly) compared to the Fox Squirrel's reddish-orange ventral hair. Nuts are the main food source for both species, including acorns, pecans, and walnuts (Koprowski 1994a, 1994b). The Eastern Gray and Fox squirrels are a common component of urban and rural areas. Viruses and bacteria associated with these species include West Nile Virus (Kiupel et al. 2003) and Leptospira (Dirsmith et al. 2013) in fox squirrels and Salmonella from eastern gray squirrels (Jijón et al. 2007). Despite co-occurring with humans and agricultural animals at relatively high densities, little to no research investigates the role of tree squirrels in harboring foodborne pathogens or zoonotic diseases.
Invasive Species
House mouse
The ancestral range for the House Mouse (M. musculus) was most likely India; however, this species readily colonized worldwide as an invasive species due to their close proximity to humans (Boursot et al. 1993, Phifer-Rixey and Nachman 2015). Wild M. musculus are distinguishable from white-footed mice and deer mice by their shorter pelage, smaller eyes, and scaly tail (Schmidly and Bradley 2016). House mice are commonly found in urban and agricultural areas, although feral populations do exist in the wild (Phifer-Rixey and Nachman 2015). Wild M. musculus is heavily studied, and this species has been shown to be a vector for many human diseases and is frequently implicated as reservoir for foodborne pathogens (including multiple pathogenic Salmonella serovars, E. coli, Clostridium difficile, and so on) and other zoonotic pathogens (e.g., avian influenza) of special interest to agricultural production systems (Table 1) (Shimi et al. 1979, Henzler and Opitz 1992, Allen et al. 2011, Burt et al. 2012).
Norway rat
The Norway Rat (Rattus norvegicus), also known as the Brown Rat, is native to China and Mongolia but now has a worldwide distribution as its range expanded alongside humans, facilitated in large part to global trade (Frittelli 2008, Puckett et al. 2016). Like house mice, Norway rats are a common component of both rural and urban environments across the United States, especially on farms. Indeed, the authors of this review personally witnessed an active infestation of hundreds of R. norvegicus on a single farm in southern Minnesota in 2019. Norway rats have connections to several of zoonotic diseases and foodborne pathogens, including Hepatitis E virus (Kanai et al. 2012), C. burnetii (Reusken et al. 2011), Salmonella (Hilton et al. 2002), and C. difficile (Himsworth et al. 2014), among others (Table 1). The ability of these two invasive species, the house mouse and the Norway rat, to adapt to any environment and their quick reproductive cycles make them especially important to understand and document as reservoirs of foodborne pathogens.
Black rat
Also known as the Roof Rat or Ship Rat, the Black Rat (Rattus rattus) is native to Southeast Asia. As in the case of the Norway Rat, the Black Rat is commensal with humans and is now cosmopolitan due to human travel. This species is a major vector for numerous pathogens across the globe, including Y. pestis (plague), Bartonella spp. (Ellis et al. 1999), Trypanosoma cruzi (Panti-May et al. 2017), and several foodborne pathogens (Table 1). The current distribution of the Back Rat in the United States is primarily associated with coastal regions, and data regarding established R. rattus populations in the central United States are limited (Lack et al. 2013). It is likely that R. rattus first colonized North America alongside Columbus in 1492 and established populations in the continental United States by the 1500s (Armitage 1993). However, R. rattus populations across the United States have dwindled, possibly a direct result of being extirpated by the larger and more aggressive R. norvegicus (reviewed in Lack et al. 2013). Given the paucity of data regarding R. rattus within the Midwestern United States, and potential for misidentification with R. norvegicus, we recommend field-based studies aimed to better understand the extent to which R. rattus is distributed on Midwestern farms, especially near shipping ports and railway hubs (e.g., the Great Lakes region, Mississippi and Missouri Rivers, and so on).
Major Bacterial Foodborne Pathogens Linked to Rodents
STEC, nontyphoidal S. enterica, Campylobacter spp., Clostridium spp., and L. monocytogenes are classified as major zoonotic foodborne pathogens in the United States (Scallan et al. 2011). From a global perspective, these zoonoses have well-documented rodent reservoirs, yet there are clear knowledge gaps regarding the interplay between these zoonoses and rodents in the United States. Thus, we focus on the following for our review:
STEC and multidrug resistant generic E. coli
STEC, also known as verocytotoxigenic E. coli (VTEC), is an important foodborne zoonotic pathogen that causes illness ranging from mild diarrhea to hemorrhagic colitis and life-threatening hemolytic uremic syndrome (HUS) (Tarr 1995, Karmali 2004). Cattle and other ruminants are considered to be natural reservoirs for STEC; however, STEC strains have been isolated from other domestic species, as well as wild animals (e.g., goats, sheep, pigs, cats, dogs, deer, wild rabbits, birds, and rodents) (Espinosa et al. 2018). For example, in 2011 a large outbreak of E. coli O157:H7 was traced back to fresh strawberries contaminated with deer feces (Laidler et al. 2013).
With respect to rodent-specific analyses, several studies have identified STEC strains circulating in peridomestic species on both agricultural and urban landscapes, including the Norway Rat (R. norvegicus), Black Rat (R. rattus), and House Mouse (M. musculus) (Čížek et al. 1999, Blanco Crivelli et al. 2012, Himsworth et al. 2015, Williams et al. 2018). In particular, Nielsen et al. reported the isolation of a STEC strain from a Norway Rat (R. norvegicus) identical to a cattle isolate from the corresponding farm with respect to serotype, virulence profile, and pulsed-field gel electrophoresis type (Nielsen et al. 2004). Moreover, in a Czech cattle farm, E. coli O157 was isolated from 40% of Norway rats, emphasizing the importance of rodents as potential vectors of pathogenic E. coli in cattle productions (Čížek et al. 1999).
In pathogen survival studies, it was shown that E. coli O157 could survive for up to 9 months in the feces of experimentally infected rodents, whereas in ruminant feces the pathogen survived for up to 18 weeks (Fukushima et al. 1999, Guenther et al. 2013). This finding suggests that rodent mobility and longer survival periods of the pathogen in rodent feces may make rodents competent vectors of such pathogens particularly when they have access to livestock production farms. In addition, limited or no data exist with respect to STEC screening of populations of native rodent species occurring on farms across the Midwestern states (e.g., Peromyscus spp., Microtus spp., Sciurus spp.; see above); thus, additional research is required to investigate the role of these rodents in spreading or harboring strains of STEC.
Nonspecific E. coli strains with antibiotic resistance or multidrug resistance (MDR) are also of great health concern, as MDR infections are complicated to treat and, in worst case scenarios, are completely without antibiotic treatment options. Moreover, MDR bacteria can greatly contribute to the dissemination of antibiotic resistance genes through horizontal gene transfer to other bacteria.
MDR bacteria have been found at high prevalence in the intestinal bacteria of wild rodents living in close proximity to livestock. For example, one study reported 71% MDR E. coli positive samples (cecum) from 49 wild house mice (M. musculus) captured from swine farms (Allen et al. 2013). Furthermore, Allen et al. (2011) studied antimicrobial resistance in generic E. coli from wild small mammals (e.g., mouse, vole, shrew) living in swine farms, residential areas, landfills, and natural environments in Ontario, Canada. They observed the highest (48%) resistant E. coli population isolated from the small mammals living in swine farms compared to other areas (Allen et al. 2011). A study by Guenther et al. (2010) reported low (5.5%) prevalence of antibiotic resistance among 188 E. coli isolated from rodents captured from rural areas in Germany, which also suggests transmission of resistant bacteria from livestock or farm environment to wild rodents. With respect to species-specific behavioral traits, it was suggested by Himsworth et al. (2015) that Black Rats (R. rattus) are less likely to carry E. coli than Norway Rats (R. norvegicus) as the latter species is a ground dweller and thus experiences more fecal exposure than the upper level dweller. Moreover, Kozak et al. (2009) found that rodents residing in swine farms were five times more likely to carry tetracycline and multidrug resistant E. coli than rodents living in natural areas. Interestingly, 83% of the swine population in the specified farms also carried tetracycline resistant E. coli. However, the direction of transmission of these resistant pathogens remains unclear.
Nontyphoidal S. enterica
Salmonellosis is a common foodborne illness worldwide, caused by the bacteria Salmonella that has over 2500 serotypes making this pathogen very challenging to control. In the United States, nontyphoidal Salmonella bacteria cause 11% of all foodborne illness, 35% of all foodborne illness related hospitalization, and 28% of all foodborne illness related death (Scallan et al. 2011). In addition, 95% of all nontyphoidal Salmonella cases are foodborne, suggesting an origin from food animals or contaminated food products derived from food animals and fresh produce (Hoelzer et al. 2011).
Many studies have isolated Salmonella at higher rates from farm environments, indicating that the overall farm environment is an important Salmonella reservoir (Bondo et al. 2016). Both wildlife and companion animals represent potential sources of human salmonellosis, as Salmonella can cause disease in these animals and some animals can be asymptomatic carriers as well (Rodriguez et al. 2006). Rodents are effective amplifiers of Salmonella (see above, Fig. 1) with the capacity to output millions of Salmonella bacteria into a particular environment. Such rodent-associated Salmonella output is within the reported infective doses for poultry and livestock (e.g., pigs and cattle), which range from 106 to 108 CFUs (Barrow et al. 1987, Gast and Holt 1998, Silva et al. 2008, Hill et al. 2016).
Poultry will preferentially feed on rodent fecal pellets if present in feed or housing facilities, which is highly suggestive of commensal rodents being a critical risk factor in production operations (Davies and Wray 1995). Umali et al. (2012) showed an important connection between persistent Salmonella infection in layer chicken houses and rodents, where R. rattus intermittently shed Salmonella up to 24 weeks, thus reintroducing the pathogen to replacement flocks after cleaning and disinfection. Camba et al. (2020) report that continual fecal shedding by R. rattus inhabiting commercial layer farms in Japan likely played an important role in shifting the predominant Salmonella serotypes identified on layer farms. Over a sampling period from 2008 to 2017, the authors describe an initial identification of Salmonella Infantis in farm-collected R. rattus; however, over time the rat-associated serotypes changed to Salmonella Corvallis in 2013 followed by Salmonella Potsdam and Salmonella Mbandaka in 2017, which became the predominant on-farm serotypes (Camba et al. 2020). Thus, rodents associated with poultry farms are a serious public health concern (Antunes et al. 2016, Nidaullah et al. 2017, Camba et al. 2020).
Several other studies reported the isolation of highly similar strains of Salmonella from food animals (chickens, pigs) and rodents (R. rattus and an undefined species) circulating in the same farm environment (Lapuz et al. 2012, Andrés-Barranco et al. 2014). Moreover, Andrés-Barranco et al. (2014) suggested a greater role of rodents compared to wild birds in maintaining S. enterica in a farm environment, where the associated livestock population was a major source of Salmonella contamination.
There are a growing number of examples documenting the relationship between rodents and Salmonella outbreaks. Although not occurring in a wild or agricultural setting, a 2004 multistate outbreak of S. enterica serovar Typhimurium was traced to hamsters, rats, and mice sold in pet shops across the United States (Hoelzer et al. 2011). Studies investigating the contamination of chicken and pork sausage by Salmonella have implicated poor rodent-control measures within processing facilities (Trimoulinard et al. 2017). It is likely that rodent-related Salmonella outbreaks occur seasonally, coinciding with key demographic and/or climatic shifts that influence rodent behavior (e.g., reproductive cycles, seeking of indoor shelter with lower autumn nightly temperatures, seeking novel food sources during periods of drought, and so on). Given the strong connection between rodents and Salmonella, we recommend renewed and proactive monitoring of rodents associated with food animal farms and food production facilities.
Campylobacter spp.
Campylobacter are major pathogens that cause significant episodes of foodborne illness and hospitalization across the global human population (Scallan et al. 2011). Campylobacter jejuni and Campylobacter coli are the species most commonly associated with human campylobacteriosis; other less frequently reported species causing infection include Campylobacter fetus, Campylobacter lari, and Campylobacter upsaliensis (Meerburg and Kijlstra 2007). Campylobacteriosis typically causes symptoms of gastroenteritis, including vomiting, diarrhea, and fever (Franco 1988). The majority of Campylobacter infections are self-limiting; however, in the case of immunocompromised individuals, hospitalization and treatment are required. Thus, antibiotic resistant strains of Campylobacter pose a great threat to public health. Poultry, contaminated retail meat, and dairy products have largely been associated with campylobacteriosis cases (Scallan et al. 2011).
Poultry and other food animals are thought to be the primary reservoir of Campylobacter spp.; however, wildlife surrounding farm environments are a potential source (Wilson et al. 2008). Viswanathan et al. (2017) studied the prevalence of Campylobacter in livestock (i.e., beef cattle, dairy cattle, and swine) and wildlife, including rodents (i.e., meadow voles (M. pennsylvanicus), house mice (M. musculus), species of Peromyscus, and Norway rats (R. norvegicus)), and identified Campylobacter across their samples (Table 1). Importantly, the presence of rodents in poultry farms has been identified as a significant risk factor for Campylobacter colonization in turkeys and broilers (Agunos et al. 2014). Hiett et al. (2002) conducted molecular subtyping analyses of Campylobacter spp. from Arkansas and California poultry operations. They isolated identical strains of Campylobacter from broiler feces and from a mouse (undefined species) captured from the same farm. Similarly, other studies have reported greater prevalence of antimicrobial resistant strains of Campylobacter from wildlife living in or surrounding farms (Agunos et al. 2014).
Several studies have investigated the role of rodents as potential reservoirs of Campylobacter spp., showing an association between the presence of rodents on farms and an increased risk for flocks to become infected with Campylobacter (Berndtson et al. 1996, Adhikari et al. 2004, Meerburg and Kijlstra 2007). These studies collectively indicate that rodents are a risk factor for the transmission of Campylobacter on food animal operations, and additional studies are required to better understand these transmission dynamics.
Clostridium spp.
The Centers for Disease Control and Prevention (CDC) estimates that ∼1 million cases of foodborne illnesses each year are linked to Clostridium species (Grass et al. 2013). The most common species causing these foodborne illnesses include C. perfringens, C. difficile, and C. botulinum. Clostridia are anaerobic, spore-forming bacteria and produce enterotoxin that can cause mild diarrhea to fatal colitis and death of humans (Keessen et al. 2011, Silva et al. 2014). Over 90% of Clostridium outbreaks are associated with meat products (e.g., beef, pork, and poultry) (Grass et al. 2013). However, very limited information is known about animal or environmental reservoirs for Clostridium spp., information that could be crucial for implementing biological control measures at animal production farms and processing facilities (McClane 2007).
Companion and food animals have been considered as potential sources of Clostridium infection (Rodriguez et al. 2017); however, only a few studies have investigated the presence of Clostridium species in peridomestic rodents despite having clear connections (Hensgens et al. 2012, Himsworth et al. 2013). Himsworth et al. (2014) reported an overall prevalence of 13.1% of C. difficile across 724 rodents (R. rattus, R. norvegicus) collected in Vancouver, Canada. Another study in the Netherlands stated high prevalence (66%) of C. difficile in house mice (M. musculus) from a swine farm, indicating the potential role of these rodents in the maintenance and transmission of the pathogen (Burt et al. 2012). Many studies have established rodents as reservoirs for C. difficile (Burt et al. 2012, 2018, Himsworth et al. 2014). In addition, Krijger et al. (2019) reported six ribotypes of C. difficile associated with human infections that were isolated from rodents captured from swine and dairy farms in the Netherlands. A study by Andrés-Lasheraset al. (2017) in Spain observed that the odds of finding C. difficile from environmental rodent (Rattus sp., M. musculus) fecal pellets were 10 times higher than from swine fecal samples, suggesting that rodents can transmit the pathogen on farms. Another study reported the isolation of identical clonal types of C. difficile from rodents (R. rattus, M. musculus) and piglets from two swine farms in Brazil, suggesting that rodents were introducing the pathogen to the piglets (de Oliveira et al. 2018). Overall, these studies suggest that on-farm rodents are potential C. difficile reservoirs.
Listeria monocytogenes
Another important foodborne pathogen is L. monocytogenes, which causes significant hospitalization and is a leading cause of death by foodborne illness (Scallan et al. 2011). Listeriosis is less commonly reported, perhaps due to its long incubation period and/or because the bacteria are not detected by routine stool culture (Smith et al. 2018). However, listeriosis can cause severe conditions, including stillbirths, abortions, septicemia, and meningitis in high risk groups such as pregnant women, elderly, young children, and immune compromised individuals (Montero et al. 2015).
L. monocytogenes can be found naturally in the environment (e.g., soil, water) and, consequently, in fresh produce, livestock, and wild animals (Weis and Seeliger 1975, Lyautey et al. 2007). Fecal carriage of L. monocytogenes is common and widely documented in food animals (cattle, goat, sheep, swine) and wildlife (mammals, birds) (Weber et al. 1995, Yoshida et al. 2000, Kalorey et al. 2006). The specific reservoir status of farm-associated rodents for Listeria spp. is poorly understood; nevertheless, a few studies have reported varying levels of Listeria presence in rodents. For example, Ayyal et al. (2019) reported 5% prevalence of Listeria spp. among 120 black rats within an urban setting in Baghdad, Iraq. Lesley et al. (2016) conducted a study in Kubah National Park (Sarawak, Malaysia) to detect the prevalence and antibiotic resistance of L. monocytogenes from wild animals (bats, rats, and shrews) and water samples. They reported 33% prevalence of L. monocytogenes from the samples collected and found the isolates to be uniformly resistant to tetracycline and erythromycin.
Another species, Listeria ivanovii, also causes listeriosis, and Cao et al. (2019) described 3.7% prevalence of the bacteria in wild rodents collected from six regions of China, suggesting that wild rodents might be a long-term host of the pathogen. Inoue et al. (1992) captured 254 wild rats, including 41 R. rattus and 126 R. norvegicus in buildings from six different areas in Kanto, Japan, and they observed high prevalence of Listeria (up to 77.8%). These same authors also reported frequent isolation of L. monocytogenes from R. rattus inhabiting Tokyo restaurants (Inoue et al. 1991). Trimoulinard et al. (2017) investigated contamination of chicken and pork sausage by L. monocytogenes, and their findings suggested a positive association with fresh rodent droppings. Additional studies are required to describe the epidemiology of Listeria spp. from commensal rodents inhabiting farms and to assess the public health risk.
Conclusions
At present, many epidemiological studies of zoonotic foodborne pathogen outbreaks in humans focus almost entirely on the role and risk factors of food animals in transmission dynamics. There remains limited published research on the role that individual species of peridomestic rodents play in the transmission of these pathogens to humans, food animals, and within food production systems of the United States. Despite the connections between rodents and zoonotic foodborne pathogens highlighted above, there is a lack of research on this subject across the diverse agricultural landscape of the United States. In light of this paucity of data, we recommend novel research that focuses on the rodent-agriculture interface and that leverages modern metagenomic approaches. For example, high-throughput metagenomic analyses are ideally suited for investigating the rodent-agriculture interface, and such approaches could elucidate the origins of zoonotic foodborne pathogens (Koskela et al. 2017, Sekse et al. 2017, Andersen and Hoorfar 2018, Carleton et al. 2019). Moreover, mammalogists must be engaged in this research to appropriately identify those rodent species that are associated with food production pipelines in the United States. This is important for several reasons, especially when considering that species-specific behaviors and natural history traits (as described above) can influence pest control measures (e.g., avoidance of poison baits), facilitating large on-farm populations.
When considering native rodent species, the local habitat and ecology surrounding farms are important for understanding seasonal population densities, as local environmental conditions can directly influence behavior and on-farm population density. Knowing the resident rodent species composition associated with our food production facilities would allow for the development of sophisticated predictive modeling of rodent populations and the implementation of species-specific biosecurity measures.
There is an undeniable connection between peridomestic rodents and agriculture that has existed since the dawn of civilization. This connection is ongoing and should not be ignored when investigating the source and/or spread of zoonotic foodborne pathogens. Therefore, we recommend the formation of multidisciplinary One Health research teams consisting of farmers, veterinarians, epidemiologists, microbiologists, and mammalogists to focus on this area of research. It is likely that such efforts will identify novel reservoirs for emerging zoonotic foodborne pathogens and will ultimately help to secure food production systems across the United States.
Footnotes
Acknowledgment
The authors thank the University of Minnesota Agricultural Research, Education, Extension and Technology Transfer program for startup funds awarded to PAL.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This study was supported by discretionary funds awarded to PAL.