
Abstract
The blacklegged tick, Ixodes scapularis, can acquire and transmit tick-borne pathogens (TBPs) responsible for diseases such as human granulocytic anaplasmosis (Anaplasma phagocytophilum [ANPH]), babesiosis (Babesia microti [BABE]), Lyme borreliosis (Borrelia burgdorferi sensu lato [BBSL]), and the relatively novel relapsing fever-like illness, Borrelia miyamotoi (BMIY) disease in the northeastern United States. Coinfections with these pathogens are becoming increasingly more common in I. scapularis and their hosts, likely attributed to their shared enzootic cycles. Urban habitats are favorable to host species such as white-tailed deer (Odocoileus virginianus) and these ungulates are known to be important to I. scapularis for reproduction and dispersal in North America. To understand the relationship between TBPs, white-tailed deer, and I. scapularis, we sampled eight sites across central Maryland collecting I. scapularis using standard tick dragging/flagging methods and retrieved others from deer carcasses. Pathogenic TBP species in each tick were determined using qPCR. In total, 903 adult ticks (deer: n = 573; questing: n = 330) revealed landscape-level prevalence of ANPH (27.8%), BABE (1.3%), BBSL (14.6%), and BMIY (0.8%) as singular infections overall. However, secondary coinfections of ANPH and BBSL were highest (9.9%) in ticks feeding from deer while associations of BBSL and BABE (4.2%) were highest in questing ticks. Results from this study provide evidence suggesting that adult I. scapularis acquire pathogenic species through phenologically associated host use, and thus, subsequent infections found in adults may provide insights into coinfection relationships.
Introduction
Tick-borne diseases are increasingly problematic and constitute great concern to public health (Richards et al. 2017). The blacklegged tick, Ixodes scapularis, is one of the most medically important tick species that can harbor and transmit numerous zoonotic pathogens, including bacteria, viruses, and protozoans (Cabezas-Cruz et al. 2018, Eisen and Eisen 2018). Several of these tick-borne pathogens (TBPs) in the eastern United States are endemic, including the protozoan Babesia microti (BABE), responsible for human babesiosis; and bacterial microbes including Anaplasma phagocytophilum (ANPH), the causal agent of human granulocytic anaplasmosis caused by the Anaplasma-ha variant (Massung et al. 2005); the Borrelia burgdorferi (sensu lato) complex (BBSL), the etiologic agents of Lyme borreliosis (Eisen 2018); and Borrelia miyamotoi (BMIY) causing the relatively novel BMIY relapsing fever-like disease (Scoles et al. 2001, Krause et al. 2013, Wagemakers et al. 2015). Because these pathogens are enzootic and share common host and vector species, coinfections frequently occur and may lead to within-host interactions (Civitello et al. 2010).
Within-host and within-vector pathogen interactions are important when assessing the potential influence of TBP maintenance in the environment. Recent advancements in genomic resources (Vayssier-Taussat et al. 2015, de la Fuente et al. 2017) are proving increasingly useful in examining coinfection transmission potentials (Kenedy et al. 2012), infectivity (Ramamoorthi et al. 2005, Abraham et al. 2017), virulence (Kasuma et al. 2016), and host-pathogen fitness (Neelakanta et al. 2010, Hajdušek et al. 2013, Narasimhan et al. 2014). Molecular approaches have also elucidated evolutionary processes and mechanisms of vector and host responses to infection and how these might influence pathogen distributions (Humphrey et al. 2010, Bonnet et al. 2017). Thus, inferences into the ecology of pathogen-specific species interactions may aid directing disease-prevention strategies (Johnson et al. 2015).
In North America, the northern expansion of I. scapularis distributions has been attributed to changes in climate (Ogden et al. 2013). Also, changes in land-use practices and habitat fragmentation have altered host community structure favoring highly adaptable generalist species, further expanding distributions of ixodid tick species, particularly I. scapularis, and the pathogens they may carry (Grunwaldt et al. 1983, Sweeny et al. 1998, Swanson et al. 2006). Most notably, white-tailed deer (Odocoileus virginianus; WTD) have experienced significant population resurgences throughout the species' historic range (Kiffner et al. 2011, Hewitt 2015, Gallo et al. 2017, Kilpatrick et al. 2017). These ungulates serve as important hosts for I. scapularis reproduction and dispersal, furthering the range of ANPH, BABE, BBSL, and BMIY throughout the United States (Campbell and VerCauteren 2011, Levi et al. 2012). Changes in climate and land-use pattern, when considered concomitantly, require urgent assessment of the occurrence and co-occurrences of TBPs on the landscape through active surveillance measures (Cartter et al. 2018, Eisen and Eisen 2018).
Here we present data collected from ongoing surveillance of TBP in Maryland. We compare differences between within-tick infection status of feeding adult ticks removed from WTD and questing adult ticks collected by dragging/flagging methods. Although some pathogens may not be harmful to humans, such as the Anaplasma—variant 1 (Trost et al. 2018), coinfections within hosts can induce facultative relationships favoring TBP maintenance in the environment. These data may provide baseline evaluations of TBP prevalence in central Maryland and evince pathogen/vector/host trends between I. scapularis and WTD.
Materials and Methods
Tick sampling
Adult I. scapularis ticks were collected from deer at six field sites between November 2015 and November 2016 across central Maryland as part of the autumn deer management regimens in Howard County, Maryland, including: Alpha Ridge (ARP; 39.32° N, 76.91° W), Blandair (BLD; 39.21° N, 76.83° W), David Force (DF; 39.29° N, 76.88° W), Friendship Park (FR; 39.61° N, 77.76° W), Gorman (GOR; 39.15° N, 76.87° W), and Middle Patuxent Environmental Area (MPEA; 39.20° N, 76.90° W). Adult blacklegged ticks were collected from dead deer during managed hunts by the Deer Management Team, Howard County Department of Recreation and Parks. No IRB approval was required. Collection of questing adult I. scapularis through flagging or dragging occurred at four sites, including BLD, MPEA, Howard County Conservancy (HCC; 39.31° N, 76.87° W) in Howard County, and the wooded area on the east campus of the U.S. Department of Agriculture, Agriculture Research Service, Beltsville (ARS; 39.02° N, 76.92° W) in Prince George's County, MD. All adult ticks were removed with forceps from deer or flags and placed in individual vials filled with ≥70% ethanol for laboratory analysis.
Tick species identification, measurement, and pathogen detection
Tick species identification was based on: Cooley (1946); Keirans and Clifford (1978); Keirans and Litwak (1989); and Durden and Keirans (1996). Ticks were categorized by species and developmental stage (larva, nymph, or adult). Species identification of I. scapularis was confirmed by a species-specific TaqMan PCR assay (Xu et al. 2019). Body size measurements of adult I. scapularis removed from deer, including scutal length, scutal width, body length, and body width, were made through Leica Application Suite X software on a Leica S9 stereomicroscope.
Scutal index (SI) was calculated as the proportion of body length/scutal width to estimate feeding duration of female ticks removed from WTD (Yeh et al. 1995, Falco et al. 1996, Gray et al. 2005). Naive and infected feeding females were included in the SI analysis. All adult males were unfed and were not included in the duration study. We identified six categorical time frames (0, 24, 48, 72, 96, and ≥120 h), and partitioned SI calculations using the range of standard errors reported in previous works (Yeh et al. 1995, Falco et al. 1996, Gray et al. 2005). Each tick was tested for TBP species, including ANPH, BABE, BBSL, BMIY, Borrelia mayonii, Ehrlichia muris-like bacterium (EMLB), deer tick virus, Powassan virus, Colorado tick fever virus, and Heartland virus.
DNA was extracted from each tick using the Epicenter Master Complete DNA and RNA Purification Kits (Epicenter Technologies, Madison, WI) following the manufacturer's protocols. ANPH, BBSL, BMIY, BABE, and EMLB were detected by a multiplex TaqMan PCR assay targeting different genes (Xu et al. 2016, 2019). Borrelia detection was performed by first applying a genus-specific detection assay for a conserved target, followed by specific qPCR assays for BBSL and BMIY. A TaqMan assay targeting hard tick 16S mtDNA gene was used as an internal control (Xu et al. 2016, 2019).
Statistical analyses
All statistical evaluations were conducted in R (R Development Core Team 2019) or the data-analysis functions of Excel. Student's T-tests (α = 0.05) were performed to determine if significant differences existed in the frequency of TBP infection between sexes both in questing adults and those removed from WTD. Figures were generated using the ggplot2 package (Wickham et al. 2018). Site prevalence was calculated treating collection method (i.e., dragging or deer) as independent factors. Although both methods may have been utilized at certain sites, sampling efforts differed temporally. Thus, landscape-level pathogen prevalence was calculated by combining all ticks collected. This pooled calculation accounted for individual site-specific contributions, the unknown origins of ticks found on deer, and minimized artificial influences of actual tick and pathogen abundances when sample sizes were nominal. The frequency of occurrence (f) of TBP coinfections over the established feeding time frames was used to estimate changes in TBP species richness over time. In addition, we used multivariate principal component analysis (PCA) to evaluate trends in secondary (2°), tertiary (3°), and quaternary (4°) coinfection relationships in ticks shown to harbor at least two TBPs (Kellow 2006, Vayssier-Taussat et al. 2015) using the R package FactoMineR (Husson et al. 2018).
Results
Landscape prevalence
We collected I. scapularis at eight sites across central Maryland. In total, 903 adult I. scapularis were collected seasonally (Table 1) using dragging/flagging methods (n = 330) and from WTD (n = 573). Ticks were opportunistically collected from 78 WTD in October and November 2015–2016 at the stated field sites during managed culls. Roughly half (43.6%, 144/330) of unfed questing adults were infected with one-or-more TBP (Table 1). When we consider the influence of fasted males from WTD (n = 268), landscape prevalence of TBP infection increased to 53.7% (321/598). Moreover, the only 4° infection with all TBPs of interest to this study was attributed to a single fasted male (Table 1).
Pathogen Frequency of Occurrence and Prevalence Found in Male (♂) and Female (♀) Ixodes scapularis Removed from the White-Tailed Deer (Odocoileus virginianus) and Collected Using Dragging/Flagging Methods at Sites in Central Maryland, USA
Primary, secondary, tertiary, and quaternary infections are indicated by 1° 2°, 3°, and 4°, respectively.
ANPH, Anaplasma phagocytophilum; BABE, Babesia microti; BBSL, Borrelia burgdorferi (sensu lato); BMIY, Borrelia miyamotoi.
With high numbers of coinfections found in our samples, comparisons of infection status between males and females required inferences derived from within-tick pathogen occurrence. We found no differences in TBP frequencies between questing males and females (t 328 = 0.83; p = 0.20). Females feeding on WTD differed significantly from fasted males removed from deer (t 464 = −6.67; p < 0.001), notably Anaplasma infection rate. Females from WTD were found to have the highest prevalence (52.5%; 160/305) of 1° ANPH (Table 1). Although most ticks were infected with a single TBP, ANPH was often coupled with BBSL, particularly within males sampled from WTD (14.6%, 39/268). No ticks were found to be infected with B. mayonii, EMLB, deer tick virus, Powassan virus, Colorado tick fever virus, or Heartland virus.
Landscape-level prevalence of each TBP varied greatly in terms of (ƒ) with ANPH and BBSL representing most infections (Table 2). ANPH was found as 1° infection in 27.7% (251/903) of ticks followed by BBSL singly infected presence in 14.6% (132/903). Not surprisingly, when examined as coinfections, ANPH-BBSL represented the highest 2° infection at 7.0% (60/903) and ANPH-BABE-BBSL 3° infections present in 1.2% (11/903) of ticks. It is also noteworthy that although coinfections with ANPH, BABE, and/or BMIY were present, the (ƒ) coinfections were not found to be related to tick feeding status (ANOVA: F5,298 = 0.82; p = 0.54) (Fig. 1). In addition, we did not observe 2° coinfections of either BABE or BMIY in association with ANPH in questing ticks (Table 1).
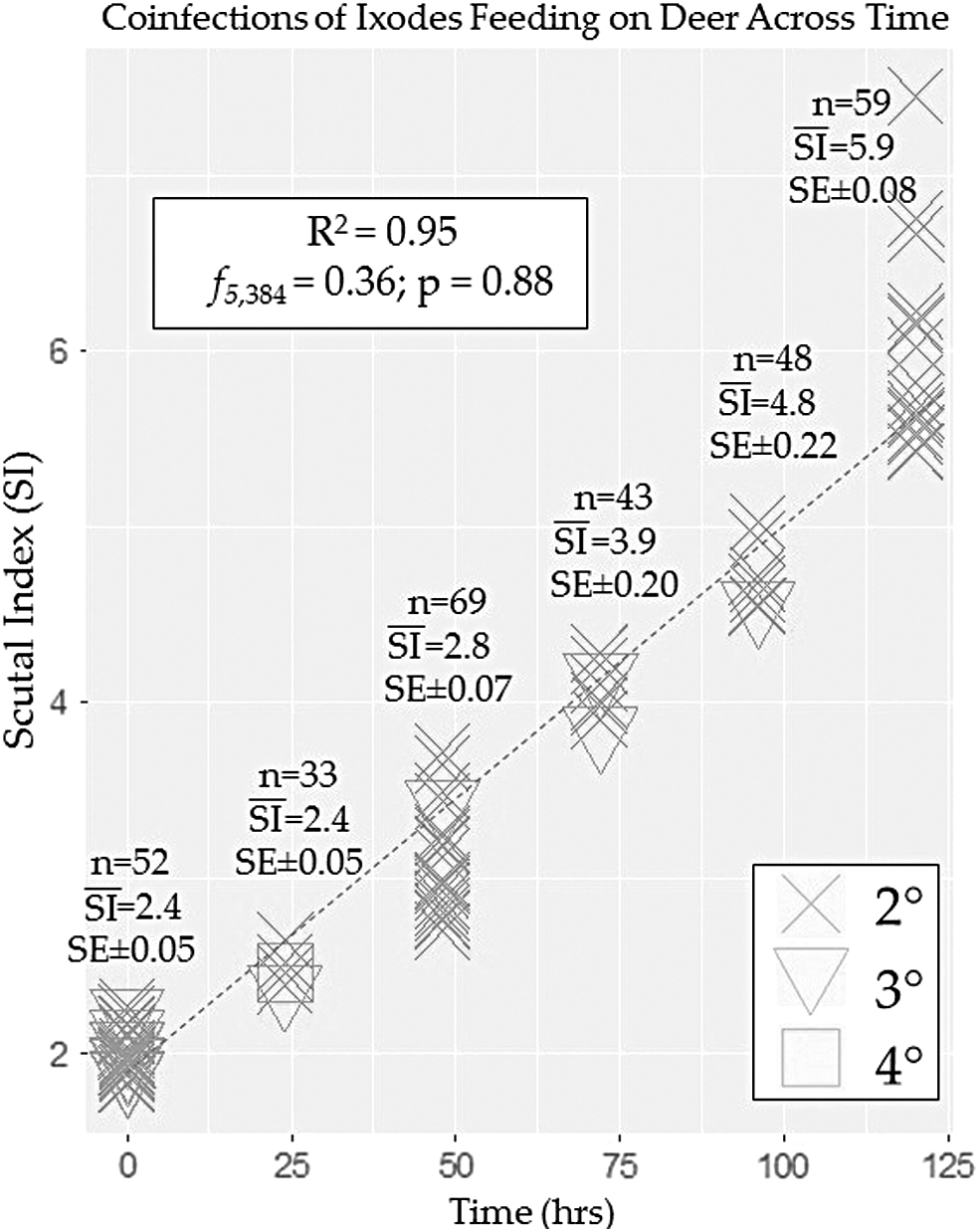
Coinfection of pathogens in adult females of Ixodes scapularis removed from white-tailed deer (Odocoileus virginianus) in relation to feeding duration estimated according to body-length/scutal-width (SI). Only individuals with secondary (2°), tertiary (3°), or quaternary (4°) coinfections are shown.
Pathogen and Coinfection Prevalence at the Landscape Level
Landscape prevalence includes male and female Ixodes scapularis (n = 903) sampled from deer and collected from the environment. The total number of infections is shown with prevalence (%) within parentheses; 1° indicates single (primary) infections; 2°, 3°, and 4° indicate the number of secondary, tertiary, and quaternary coinfections, respectively.
Principal component analysis
As described above, multivariate PCA is helpful in terms of elucidating relationships in often complex and nested interactions. Because of the biological relationships and influences among and between cocirculating TBPs, we used PCA to explore the relationships between coinfecting pathogens in questing and feeding adult I. scapularis. We found that coinfected questing ticks in our study area tended to have BBSL-related 2° infections shared with either BABE or BMIY, although to a lesser degree (Fig. 2). However, after adult females have fed on WTD, BBSL-associated 2° infections appear to have a higher affinity for ANPH than BABE or BMIY (Fig. 2). In our data set, although low in overall sample size, we are able to account for approximately half of the variation normally associated with coinfection relationships in questing ticks and coinfections observed in feeding ticks (51.6% and 46.18%, respectively) when compared with random chance alone.

Principal component analysis showing coinfection relationships between Anaplasma phagocytophilum, Babesia microti, Borrelia burgdorferi (sensu lato), and Borrelia miyamotoi in questing
Discussion
In the current study, we compare the presence of TBPs responsible for human disease in questing and WTD-associated adult I. scapularis in central Maryland. We also highlight important relationships among TBPs known to be carried and successfully transmitted by this vector tick species. In addition, we were able to present evidence of multiple coinfections circulating in Maryland and provide inferences into pathogen acquisition of ANPH by female I. scapularis feeding on WTD. Although specific variants of ANPH cause disease in humans, nonpathogenic ANPH is known to facilitate the success of other TBPs (Trost et al. 2018) through microecological interactions (Richards et al. 2017).
It has been shown that ANPH-BBSL coinfections are advantageous to pathogen-vector fitness (Massung et al. 2005, Rikihisa 2011). When comparing this 2° infection between feeding and questing ticks, our data suggest that ANPH is largely acquired by feeding on WTD although we found no association with feeding duration. The acquisition of ANPH is beneficial to I. scapularis survival in colder temperatures and over winter (Neelakanta et al. 2010). Thus, maintenance of TBPs in the environment is likely tied to the facultative relationships shared between TBPs and their vector hosts (Moutailler et al. 2016, Sanchez-Vicente et al. 2019). We recommend further exploration into the premise of the lethality of WTD blood on BBSL (Telford et al. 1988), as coinfections with other TBPs circulating in the environment may facilitate BBSL maintenance in WTD. While the role of BMIY in TBP maintenance remains unclear (Krause et al. 2013), our data provide support for Han et al. (2016) suggesting that WTD may play a role in the maintenance of BMIY through host use as seen in other artiodactyl species (Ticha et al. 2016).
Landscape-level prevalence of TBPs derived from questing and feeding adult I. scapularis ticks can be difficult to accurately estimate (CDC 2019). When considering prevalence estimates, it is important to consider that adult male ticks are not typically found feeding on WTD. Therefore, prevalence can resemble infection statuses of fed nymphal-stage ticks when estimated from WTD collections. In addition, the retention of TBPs between life stages can vary depending upon the interactions between coinfections within the ticks (Swanson et al. 2006). Conceptually, the natural movement patterns of WTD may not necessarily reflect pathogen prevalence of a given study area where questing adult ticks are found (Eisen and Eisen 2018). Also, in open systems, the infection status of unfed female ticks is unknown, thus making it difficult to ascertain the source of recently acquired pathogens or determine cascading consequences of hierarchical TBP acquisition leading or contributing to successful coinfections (Vayssier-Taussat et al. 2015).
Active surveillance of TBP circulation should not only focus on the tick vectors but also consider the pathogens residing in WTD blood and other tissues. In addition, future research should investigate the role of mesocarnivores, such as coyotes (Canis latrans), raccoons (Procyon lotor), red fox (Vulpes vulpes), given the lack of knowledge regarding the role of TBP maintenance provided by these mammals in the northeastern United States.
Conclusions
Our surveillance of TBP in the vector tick species I. scapularis uses comparisons of TBP infections in feeding and questing adult ticks to elucidate the potential role WTD play in pathogen acquisition on the Maryland landscape. In addition, infections of ANPH were detected in I. scapularis associated with WTD. We also found associations of BBSL-BABE and BBSL-BMIY coinfections in questing ticks, while ANPH-BBSL coinfections were strongest in deer-fed ticks.
Footnotes
Acknowledgments
The authors thank Phil Norman, Brenda Belensky, and the rest of the deer project team at Howard County Department of Recreation and Parks, Maryland, for support in collecting ticks from deer during managed hunts. They also thank James McCrary, USDA-ARS, Invasive Insect Biocontrol and Behavior Laboratory, Beltsville, Maryland, for collecting questing ticks for this work.
This article reports the results of research only. Mention of a proprietary product does not constitute an endorsement or a recommendation by the USDA for its use. The USDA is an equal opportunity provider and employer.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This study was supported by the USDA-ARS in-house Project (#8042-32000-008-00) and by funds from the Areawide Tick IPM Project supported by the USDA-ARS Office of National Programs.