
Abstract
Blue mussels (Mytilus edulis) are among the most consumed fishery products globally. Foodborne outbreaks of mussel-associated infections by viral, bacterial, and parasitic pathogens have been reported in the last years. In this study, we investigated the occurrence, genetic diversity, and zoonotic potential of the diarrhea-causing enteric protozoan Giardia duodenalis in blue mussels from Caleta Córdova in Chubut Province, southeast Patagonia, Argentina. A total of 344 free-living blue mussels were collected and distributed in 53 aliquots of pooled mussel tissue (each containing 5‒7 specimens) during the period 2015‒2018. Conventional optical microscopy was used as screening method for the detection of G. duodenalis cysts in pooled, homogenized tissues. Samples with a positive result were assessed by a multilocus sequence genotyping scheme based on the amplification of partial fragments of the glutamate dehydrogenase and β-giardin genes of the parasite. G. duodenalis cysts were found in 30.2% (16/53) of the aliquots of pooled mussel tissue tested. PCR and sequencing analyses revealed the presence of G. duodenalis subassemblage BIV in selected aliquots. To the best of our knowledge, this is the first description of zoonotic subassemblage BIV from blue mussels in Argentina.
Introduction
Foodborne parasitic diseases (FBPD) have become increasingly recognized as important foodborne pathogens during recent decades, affecting not only people's health but also causing serious economic consequences (Trevisan et al. 2019). Potential causes of the emergence and reemergence of FBPD include climate change and the rising human population (Robertson 2018). The latter has led to the implementation of new food production systems, increased global trade of foodstuff, new dietary habits and trends, and increased consumption of raw or undercooked animal products, such as fish, meat, and shellfish (Broglia and Kapel 2011). Indeed, outbreaks of gastrointestinal illness associated with the consumption of crustaceans, shellfish, molluscs, and their product represented 3.8% of the foodborne outbreaks reported in the European Union in 2017, affecting 243 people (EFSA and ECDC 2018).
Blue mussels (Mytilus edulis) are marine edible bivalve molluscs that filter large volumes of water (100–120 L/24 h). Particles in suspension, including nutrients and also bacteria, viruses, parasites, and others water pollutants, can thereby become concentrated in their organs and tissues. These pathogens can remain viable in contaminated shellfish that can act as potential sources of human infections, as people often eat them undercooked, steamed, or even raw (Tedde et al. 2019, Manore et al. 2020). Contamination of blue mussels by parasites, including the diarrhea-causing protozoan species Giardia duodenalis and Cryptosporidium spp., is not infrequent (Adell et al. 2014); in fact, mussels are considered good candidates to be used as sentinel organisms in coastal pollution monitoring (Beyer et al. 2017).
The enteric protozoan parasite G. duodenalis is regarded as the fifth leading cause of foodborne illnesses (485.000 estimated infections) in the WHO European Region in 2010 (World Health Organization 2017). This pathogen, transmitted through the fecal-oral route, is also a major contributor to the burden of diarrheal illness globally. It is estimated that 280 million human cases of symptomatic giardiasis occur every year (Einarsson et al. 2016). G. duodenalis is regarded as a complex species comprising eight (A–H) assemblages with marked differences in genetic variability and host specificity and range (Cacciò and Sprong 2010). Assemblages A and B are the dominant assemblages in humans but are also commonly detected in a broad range of other mammal species and are, therefore, considered zoonotic (Ryan et al. 2019). Host-specific canine (C, D), bovine (E), and feline (F) assemblages have been sporadically reported in humans, whereas assemblages G and H have been only identified in rodents and marine pinnipeds, respectively (Ryan and Cacciò 2013). Human isolates within assemblages A and B are further categorized within five (AI, AII, AIII, BIII, and BIV) subassemblages based on multilocus genotyping (MLG) and single-nucleotide polymorphism data of the small subunit ribosomal RNA, glutamate dehydrogenase, β-giardin, and triose phosphate isomerase genes of the parasite (Lebbad et al. 2010).
Giardia duodenalis has been reported in shellfish from Asian, European, and North American countries (Robertson 2007, Gómez-Couso and Ares-Mazás 2012, Ligda et al. 2019). To date, two foodborne outbreaks of giardiasis associated to the consumption of raw oysters have been documented in the United States (Ryan et al. 2019). However, molecular studies aiming at investigating the molecular diversity of the parasite are scarce. A Spanish survey revealed the presence of assemblage A in saltwater clams (Macoma balthica, Macoma mitchelli) and assemblage B in flat oysters (Ostrea edulis), all of them used for human consumption (Gómez-Couso et al. 2005). The role of shellfish in the epidemiology of G. duodenalis in Argentina is currently unknown, as no prevalence and/or molecular studies have been conducted to date in seafood. In this regard, the presence of trematode parasites belonging to the family Gymnophallidae, some of them potentially zoonotic, has been previously reported in molluscs of commercial interest on the coasts of Argentina and Uruguay (Vázquez et al. 2018). Investigations assessing the microbial quality of sea water have also confirmed the presence of fecal bacteria in coastal areas of the country, including the same region (San Jorge Gulf) where this survey was conducted (Pucci et al. 2012, Estevao Belchior et al. 2016, Verga et al. 2020).
In Argentina, the prevalence of human giardiasis is between 6% and 44%, depending on the clinical status, sociodemographic conditions, and geographical area of the populations investigated (Soriano et al. 2005, Gamboa et al. 2014, Garraza et al. 2014). In the Patagonian coastal region, the reported prevalences varied from 6% to 36% (Sánchez Thevenet et al. 2004, Torrecillas et al. 2014). This is an area with intense fishery and shellfish picking activities. It is estimated that 38% of its population consume locally collected mussels (unpublished data). It should also be noted that there are no sewage treatment plants in the area, so the coastal strip is exposed to the impact of sewage-derived pollutants from the surrounding urban centers, which effluents discharge directly into the sea with no prior decontamination treatment.
In this molecular epidemiological survey, we present novel data on the occurrence and genetic diversity of G. duodenalis in blue mussels collected in the coastal region of Chubut Province (Patagonia, Argentina). These data were used to make a preliminary assessment of the role of blue mussels as a suitable source of human giardiasis.
Materials and Methods
Ethics
The study was approved by COBIMED ref numbre: 16/2015 (Committee for Bioethics and Research Ethics-Faculty of Medical Sciences, Universidad Nacional de la Plata).
Study area
This study was carried out in the coastal area of Caleta Córdova, located 45° 45′ S, 67° 21′ W of the San Jorge gulf, in the province of Chubut, Patagonia, Argentina (Fig. 1).
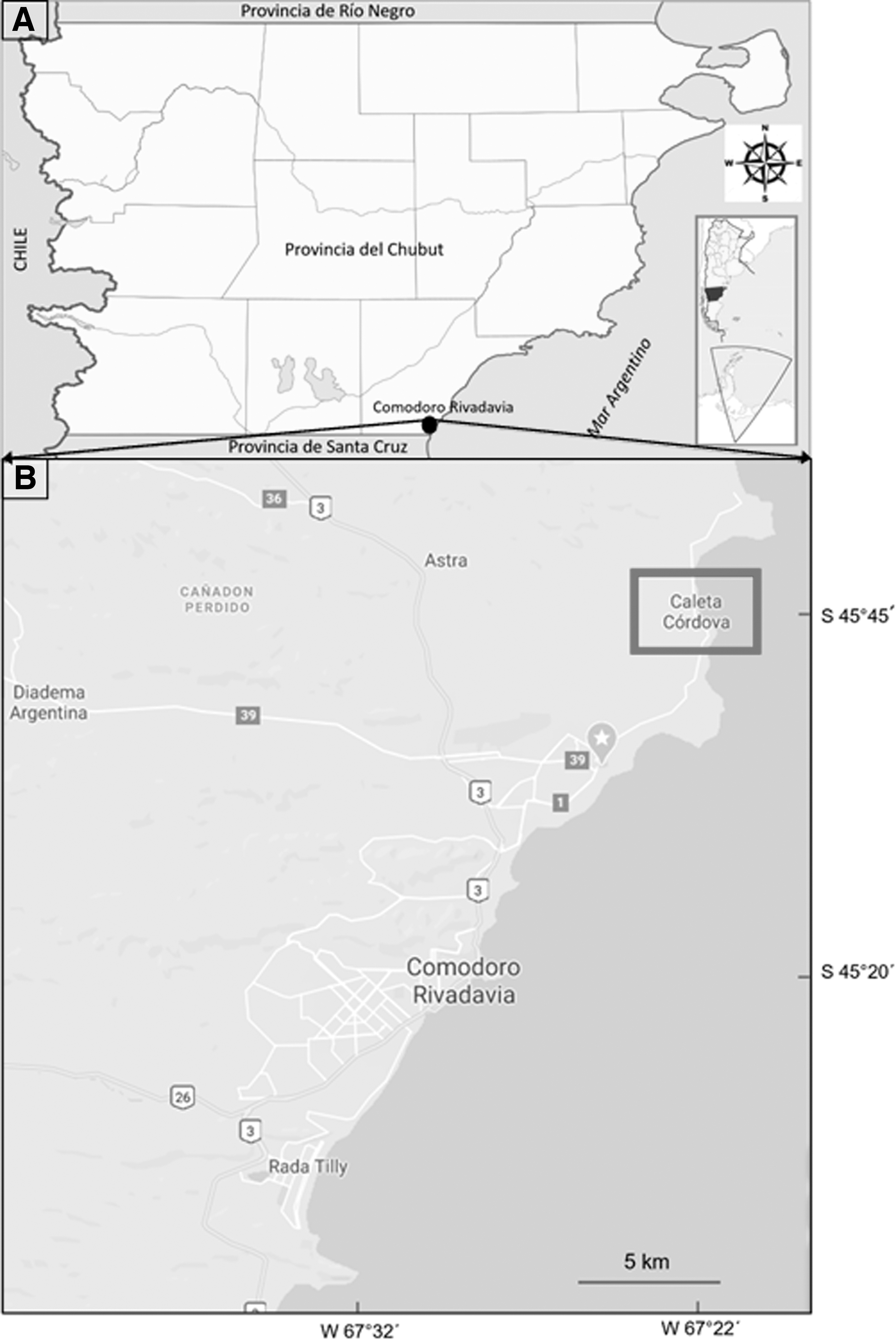
Geographic localization of the study area.
Mussel sample collection and processing
A total of 344 samples of free-living blue mussels (M. edulis) with valve sizes ranging from 3.5 × 5.5 cm (±0.5 × 0.7 cm) were collected by convenience sampling during the spring (September to December) or summer (January to March) months at 4 consecutive periods from 2015 to 2018 (Table 1). The average temperature of sea water was 13.5°C. Mussels were placed in containers with an airtight lid and immediately transported to the laboratory, where they were stored at 4°C and processed within 12 h of collection.
Microscopy-Based Samples of Mitylus edulis Positive to Giardia duodenalis Cysts from Caleta Córdova (Chubut Province, Patagonia Argentina)
Conventional microscopy with Lugol staining.
Pooled mussel tissue aliquots, each one containing five to seven individual mussels, were processed and analyzed. To do so, the specimens were aseptically opened at the hinge with a sterilized scalpel and processed as described by (Gómez-Couso et al. 2004). After homogenization and concentration, each pooled tissue sample was separated in three aliquots: (i) aliquot A was resuspended in 1.5 mL of 10% formal saline and used for conventional microscopy examination, (ii) aliquot B was kept in 70°C alcohol for downstream molecular analyses, and (iii) aliquot C was preserved at −20°C as backup.
Optical microscopy
Conventional microscopy examination was used as screening test. Aliquot A was centrifuged at 1250 rpm for 5 min and the supernatant discarded. Fresh smears of the whole resulting sediment were prepared with physiological solution and extemporaneous Lugol staining and microscopically examined at 10 × and 40 × magnification by three independent microscopists for the presence of parasite structures.
DNA extraction and purification
Total DNA was extracted and purified from selected homogenates with a positive result to G. duodenalis by conventional microscopy. An aliquot (∼200 μL) of each sample (aliquot B) was processed using the QIAamp® DNA Stool Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer's instructions. Purified DNA samples (200 μL) were stored at −20°C for further molecular analysis.
Molecular characterization of G. duodenalis
Selected G. duodenalis-positive pooled samples by conventional microscopy were investigated by sequence-based MLG of the genes encoding for the glutamate dehydrogenase (gdh) and β-giardin (bg) proteins of the parasite. A seminested PCR was used to amplify a partial fragment of the gdh gene (Read et al. 2004). PCR mixtures (25 μL) included 5 μL of template DNA and 0.5 μM of the primer pairs GDHeF/GDHiR in the primary reaction and GDHiF/GDHiR in the secondary reaction. A nested PCR was used to amplify a partial fragment of the bg gene (Lalle et al. 2005). PCR mixtures (25 μL) consisted of 3 μL of template DNA and 0.4 μM of the primer sets G7_F/G759_R in the primary reaction and G99_F/G609_R in the secondary reaction. The main features of the PCR methods used are shown in Table S1. Seminested and nested PCR reactions were run on a 2720 thermocycler (Applied Biosystems). Reaction mixtures included 2.5 units of MyTaq DNA Polymerase (Bioline GmbH, Luckenwalde, Germany) and 5 × MyTaq Reaction Buffer containing 5 mM dNTPs and 15 mM MgCl2. Water (no-template) and genomic DNA (positive) controls were included in each PCR run. gdh-PCR and bg-PCR amplicons were visualized on 2% D5 agarose gels (Conda, Madrid, Spain) and stained with Pronasafe nucleic acid staining solution (Conda). Positive-PCR products at the gdh and bg loci were directly sequenced in both directions by capillary electrophoresis using the BigDye® Terminator chemistry (Applied Biosystems).
Sequence analyses
Raw sequencing data were viewed using the Chromas Lite version 2.1 sequence analysis program (
Results
G. duodenalis cysts were found by conventional microscopy in 30.2% (16/53) of the pooled mussel tissue aliquots collected (Table 1). No obvious differences in prevalence rates based on the sampling season were observed. Genomic DNA was extracted and purified from three representative pooled samples and used for genotyping purposes. Only one of them was successfully amplified both at the gdh and bg markers. Subsequent sequence analyses revealed the presence of G. duodenalis assemblage B, subassemblage BIV.
Discussion
In this study, zoonotic G. duodenalis subassemblage BIV was detected in blue mussels from Caleta Córdova, Argentina. The finding of G. duodenalis subassemblage BIV in marine mussels has relevant public health implications. First, considering that BIV is the most prevalent genetic variant of the parasite circulating in humans (Feng and Xiao 2011), it seems reasonable to suspect that the BIV genotype identified in mussels has a human origin. Contamination of marine aquatic environments with G. duodenalis cysts may occur when surface waters and aquifers are contaminated with untreated urban sewage discharge in coastal areas (Géba et al. 2020), as it is the case of the surveyed region in the present study. Second, local consumption of BIV-infected mussels may be a source of foodborne giardiasis for human and animal populations (Fayer and Xiao 2008).
In Argentina, G. duodenalis has been previously identified in environmental samples, including fecal animal matter, soil, and surface, and drinking waters in different areas of the country (Abramovich et al. 2001, Soriano et al. 2001, Sánchez Thevenet et al. 2003). In the Chubut province, G. duodenalis cysts have been found in soil samples and canine feces collected in public spaces for recreational use (Sánchez Thevenet et al. 2004). Regarding genetic diversity, assemblage B (unknown subassemblage) is responsible for 82–93% of the reported human cases, with subassemblage AII being identified in the remaining cases (Minvielle et al. 2008, Molina et al. 2011).
In Argentina, the Argentine Food Code (CAA) and the National Secretariat for Agrifood Health and Quality (SENASA) do not regulate the detection and maximum allowed concentration of parasites in edible molluscs and other shellfish. However, the growing evidence around the world highlights the need of effective control and prevention interventions to minimize the risk of foodborne disease. Only in the United States, at least 38 foodborne outbreaks of giardiasis have been documented, 2 of them involving the consumption of contaminated mussels (Adam et al. 2016, Ryan et al. 2019). Measures to be implemented should include standardized detection and quantification methods, proficient food safety authorities, improved regulation and enforcement of food safety legislation, and development of better surveillance and outbreak investigation systems (Ryan et al. 2019). Protozoan parasites, including G. duodenalis and Cryptosporidium sp., pose additional public health challenges, as standard disinfection and removal procedures used in water and food industries are ineffective in inactivating/removing their cysts and oocysts (Willis et al. 2013). Furthermore, more research should be conducted to ascertain how cooking and/or preservation (e.g., freezing) temperatures may affect the viability and infectivity of (oo)cysts in food matrices. Establishment of good practices in the food production industry, food services, and consumers are essential for controlling and preventing FBPD (Ryan et al. 2019, Tedde et al. 2019).
This study has some methodological limitations that should be taken into consideration when interpreting the obtained results and the conclusions reached. First, conventional optical microscopy was used as a screening method. Its limited diagnostic sensitivity suggests that the true occurrence of G. duodenalis in free-living mussels may be higher than that reported here. Second, molecular genotyping analyses were conducted in a limited number of G. duodenalis-positive samples, suggesting that genetic variants other than BIV may be also present in mussels. Third, suboptimal preservation of DNA samples may have negatively influenced the performance of the conducted PCR methods.
Conclusions
This is the first description of zoonotic subassemblage BIV from free-living blue mussels in Argentina. Mussels contaminated with viable cysts are a potential source of foodborne human giardiasis. Our results demonstrate the need for public health intervention; education and awareness raising may be important components of FBPD control.
Footnotes
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This study was funded by grants from ReNIS (Registro Nacional de Investigaciones en Salud, Ministerio de Salud, Argentina) ID002752.
Supplementary Material
Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.