
Abstract
Ross River virus (RRV) is a mosquito-borne zoonotic arbovirus associated with high public health and economic burdens across Australia, but particularly in South East Queensland (SEQ). Despite this high burden, humans are considered incidental hosts. Transmission of RRV is maintained among mosquitoes and many nonhuman vertebrate reservoir hosts, although the relative contributions of each of these hosts are unclear. To clarify the importance of a range of vertebrates in RRV transmission in SEQ, a total of 595 serum samples from 31 species were examined for RRV exposure using a gold-standard plaque reduction neutralization test. Data were analyzed statistically using generalized linear models and a coefficient inference tree, and spatially. RRV exposure was highly variable between and within species groups. Critically, species group (“placental mammal,” “marsupial,” and “bird”), which has previously been used as a proxy for reservoir hosts, was a poor correlate for exposure. Instead, we found that generalized “diet” and greater “body mass” were most strongly correlated with seropositivity. We also identified significant differences in seropositivity between the two major possum species (ringtail possums and brushtail possums), which are ecologically and taxonomically different. Finally, we identified distinct hotspots and coldspots of seropositivity in nonhuman vertebrates, which correlated with human notification data. This is the largest diversity of species tested for RRV in a single study to date. The analysis methods within this study provide a framework for analyzing serological data in combination with species traits for other zoonotic disease, but more specifically for RRV highlight areas to target further public health research and surveillance effort.
Introduction
Ross River virus (RRV) accounts for the greatest number of human arboviral infections in Australia (Australian Government 2018). It is an alphavirus of the Togaviridae family, and is associated with clinical symptoms, including polyarthritis, rash, fever, and myalgia (Harley et al. 2001). At present, there is no vaccine or treatment available for RRV and management of individual human cases focuses on symptomatic care (Harley et al. 2001). As RRV is a zoonotic arbovirus that is maintained in complex transmission dynamics between multiple vectors and nonhuman vertebrate hosts, current vector-centric efforts to reduce RRV transmission would be enhanced by also considering the roles of environmental drivers and reservoir communities in transmission (Garchitorena et al. 2017).
The vectors of RRV are diverse: the virus has been isolated from more than 40 species of mosquitoes to date (Harley et al. 2001, Russell 2002). These species are predominantly of the Aedes and Culex genera and persist in a range of climates and habitats, including freshwater and saltwater environments. The different vertebrate feeding patterns exhibited by these mosquitoes may also alter their importance as vectors (Stephenson et al. 2019). A recent review of the nonhuman reservoirs of RRV found that a broad range of vertebrate species may play a role in the maintenance and transmission of the virus, including marsupials, birds, horses, and flying foxes (Stephenson et al. 2018). Along with reports of RRV transmission in humans in the Pacific Island Countries, in the absence of marsupial hosts (Aubry et al. 2015, Aubry et al. 2017, Lau et al. 2017), these studies challenge the long-held dogma that marsupials are more important reservoirs than placental mammals, which in turn are more important than birds.
For other zoonotic arboviruses, including West Nile virus (WNV), Sindbis virus, and Murray Valley encephalitis virus, serosurveys have been informative in understanding transmission among nonhuman vertebrate species (Marshall et al. 1982, Lundström et al. 2001, Kilpatrick et al. 2006, Carver et al. 2009). Although serosurveys do not indicate host competence, they do provide evidence of past exposure by measuring antibodies in blood, which are easier to detect and typically longer lasting than the infectious agent itself (Gilbert et al. 2013). RRV serosurveys in nonhuman vertebrates have previously targeted marsupial populations due to these animals being considered the primary vertebrate reservoir. To date, other species potentially important for RRV disease ecology, such as birds and flying foxes, have been greatly underrepresented in the literature (Stephenson et al. 2018).
Serosurveys are most informative when combined with additional data, such as spatial, temporal, or species trait information. This can be challenging in free-living populations as it requires repeated sampling from the same individuals or populations (Gilbert et al. 2013). For RRV, while there are 30 published serosurveys in nonhuman species, most of these only tested a single taxonomic group in a single location, or used different serological methods, making it difficult to interpret results (Stephenson et al. 2018). A recent meta-analysis took a novel approach to interpreting RRV serology by assessing life-history traits for any mammalian species that had been reported in the literature to have previous exposure to RRV (Walsh 2019). The findings of that study were that diet range, population density, and length of gestation positively correlated with exposure to RRV.
This study aims to identify infection status of nonhuman vertebrate hosts to RRV in South-East Queensland (SEQ) and analyze ecological and spatial determinates of seroprevalence. The majority of human notifications for RRV originate in this region (Australian Government 2018). This, in combination with its subtropical climate (promoting a diversity of habitats and species), makes it an ideal location to study the disease ecology of a multihost vector-borne disease.
Materials and Methods
Sample collection and storage
The cross-sectional study included wildlife and domestic horse populations in SEQ from February 2017 to June 2018 and was performed in accordance with Griffith University Animal Ethics Committee standards (approval number: ENV/04/17/AEC). Clinics were largely recruited through the Australian Veterinary Association and Wildlife Health Australia. Serum samples were collected from horses of consenting owners and wildlife admitted for veterinary care. To avoid a selection bias, this study excluded horses with clinical symptoms of RRV (pyrexia, ataxia, stiffness, lameness, swollen joints, reluctance to move, lethargy, inappetence, mild colic and “poor performance”) up to 1 year before admission. Each sample was accompanied with the following information: species, sex, age (either number of years for horses or age class (juvenile, subadult, adult, or mature adult) for wildlife), location animal was found/housed, reason for veterinary attention, and any additional relevant note (such as body condition score).
For horses, samples were collected using a 19-gauge needle from the jugular vein into a plain tube and allowed to coagulate at ambient temperature. For wildlife, samples were collected in serum separator tubes using a 22–27-gauge needle from the jugular vein, caudal vein, or medial metatarsal vein, depending on the species and the individual animal. A minimum of 400 μL of whole blood was collected from each individual, unless this exceeded 10% of their body weight, in which case, no more than 10% of their body weight was collected. Whole blood samples were labeled, centrifuged at 5514 × g for 6 min, and stored at −20°C until analysis.
Sample analysis
Vero cells (ATTC # CCL-81) were seeded at 4 × 105 cells/mL with 1 mL added in each well for 12-well plates and cultured at 37°C for 24 h. Serum samples were inactivated for 30 min in a 56°C water bath and diluted 1:10 in serum-free Opti-MEM media (Gibco). RRV (T48) was diluted to 350 plaque-forming unit (PFU) in 500 μL and mixed with an equal volume of diluted serum and incubated for 1 h at 37°C. A volume of 400 μL virus/serum mixture (giving a final virus concentration as 140 PFU per well) was then inoculated onto the vero cells after removing the culture media and incubated for an additional 2 h at 37°C with gentle shaking every 10–15 min. The mixture was removed and 1 mL of a 1% agarose overlay containing 1% penicillin/streptomycin (Gibco) and 2% fetal bovine serum was added to each well. Plates were incubated for 48–60 h at 37°C, after which the agarose overlay was removed and cells were stained with 0.1% crystal violet solution (in 20% EtOH). Plates were washed with tap water, dried, and the plaques were counted. Samples were considered to have neutralizing antibodies if the test serum sample had at least 50% less plaques than the virus-only control wells.
To ensure the antibodies we detected were specific for RRV, we assessed a common co-circulating alphavirus, Barmah Forest virus (BFV). BFV-positive sera were not able to neutralize RRV in our PRNT assay. Although BFV and RRV both co-circulate in SEQ, they are in phylogenetically distinct complexes belonging to separate serocomplexes. Furthermore, others have and not found cross-reactivity between RRV and BFV (Dalgarno et al. 1984). Sindbis virus has historically been reported to circulate in mosquito populations in Queensland (Sammels et al. 1999), but is not considered a common arbovirus and was not tested for cross-reactivity in this study.
Species trait data
A dataset of ecological and life-history traits potentially relevant for RRV exposure was compiled for each of the vertebrates (Supplementary Table S1). Traits included the following: longevity; mean number of offspring per season; mean body mass, diet rank based on increasing protein and energy content (1 = grass/leaves, 2 = seeds, forbs, grass, roots, and fungi, 3 = nectar, gum, and insects [insects <50% of the diet], and 4 = insects of vertebrates [>50% of the diet]); habitat number (number of categories of vegetation structure in which the species occurs); estimated age class of the individual; season the sample was collected in; and associated taxonomic information (order, family, genus, and species group). For mammals, these data were obtained from the PANTHERIA dataset (Jones et al. 2009), and for birds, it was obtained from Garnett et al. (2015).
Statistical analysis
Generalized linear models (GLM), with a binomial error distribution and stepwise variable selection based on Akaike Information Criterion, were used to evaluate the relative importance of each of our explanatory variable on seroprevalence. We fitted a full model, including all variables (all variables listed in Species Trait Data section), and then a reduced model, including just those variables selected in the stepwise procedure using the function “step” in R. These variables were then used to construct a conditional inference tree using the function “ctree” in the R package “party” (Hothorn et al. 2006).
To analyze spatial trends for RRV serology in nonhuman vertebrates, a two-step approach was used. First, point maps demonstrating positive and negative results were constructed for the most commonly assessed species and taxa, namely, brushtail possums, ringtail possums, koalas (Phascolarctos cinereus), horses (Equus caballus), flying foxes (Pteropus alecto and Pteropus poliocephalus), and all bird species. Second, to identify spatial clusters of positive or negative vertebrates, a hotspot analysis was undertaken in ArcGIS, which uses the Getis-Ord Gi* statistic (Getis and Ord 2010). This analysis works by assessing each point in the context of surrounding points. For example, if a single sample is positive, but the samples surrounding it are negative, it would not be statistically significant as a hotspot of seropositivity. Instead, hotspots of seropositivity must be positive and also in close proximity to other positive samples.
Results
Collected samples
Serum samples from a total of 595 individuals from 31 species were tested for the presence of RRV antibodies. Of these, 327 individuals or 54.96% (95% confidence interval [CI]: 40.17–69.1) from 23 species were seropositive (Supplementary Table S2). Antibodies to RRV were detected across all species groups (marsupials, placental mammals, and birds) (Fig. 1), with the highest average seroprevalence reported in placental mammals (70.83%; 95% CI: 61.84–78.78) and lowest in birds (28.87%; 95% CI: 20.11–38.95), although high heterogeneity was observed between-species and within-species groups. Results for individual species are reported in the Supporting Information (Supplementary Table S2).
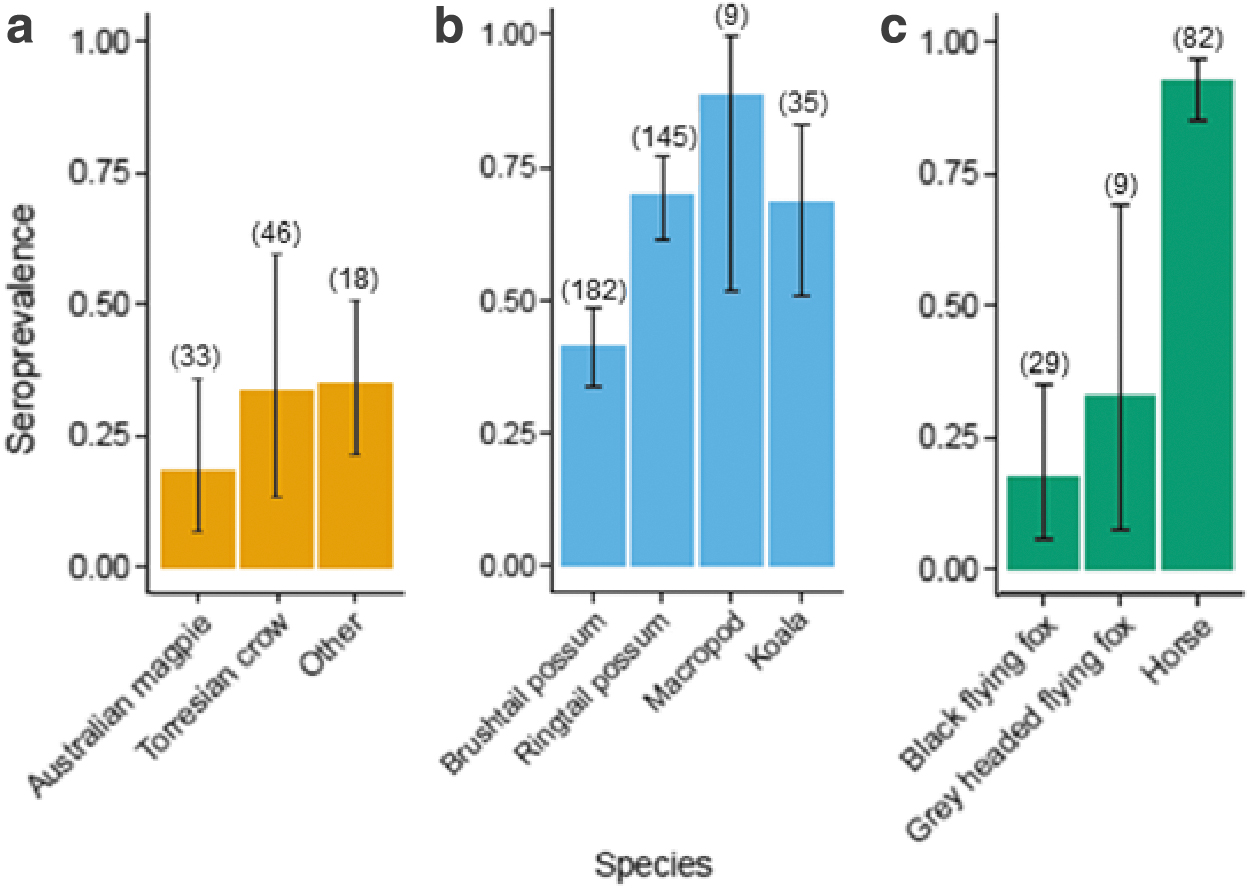
The prevalence of RRV antibodies, expressed as a proportion on the y-axis, for species (x-axis) with a minimum of nine samples, grouped by
Species traits
From the full GLM model, only log body mass and diet significantly correlated with seroprevalence (Table 1). The reduced GLM model retained log body mass, maximum longevity, number of offspring per season, social group size, and diet, and thus were selected for the conditional inference tree analysis.
Standard Error and p Values for Each Variable Included in the Generalized Linear Model Analysis
The conditional inference tree (Fig. 2) indicated that diet, log body mass, number of offspring, and longevity drive key splits in the dataset between seropositive and seronegative individuals, and that the crucial (first) dichotomous split was based on diet. Species with the lowest protein intake (diet ≤1: namely grazers, including horses and macropod species) had higher seropositivity than all other species (Fig. 1). The second node split species based on their log mass, with heavier animals (in this case horses) having higher seropositivity. Log mass also separated pigeons (0% seropositive) from ringtail possums (69.7% seropositive) in node 10. Among animals with higher protein diets, carnivores (diet >3) had lower seropositivity than the combined omnivore group (diet ≤3), which then split according to number of offspring per season, effectively separating ringtail possums from the other possum species, as well as separating birds from flying foxes. Finally, the longevity split in node 7 largely separated flying foxes from brushtail possums and short-eared possums.
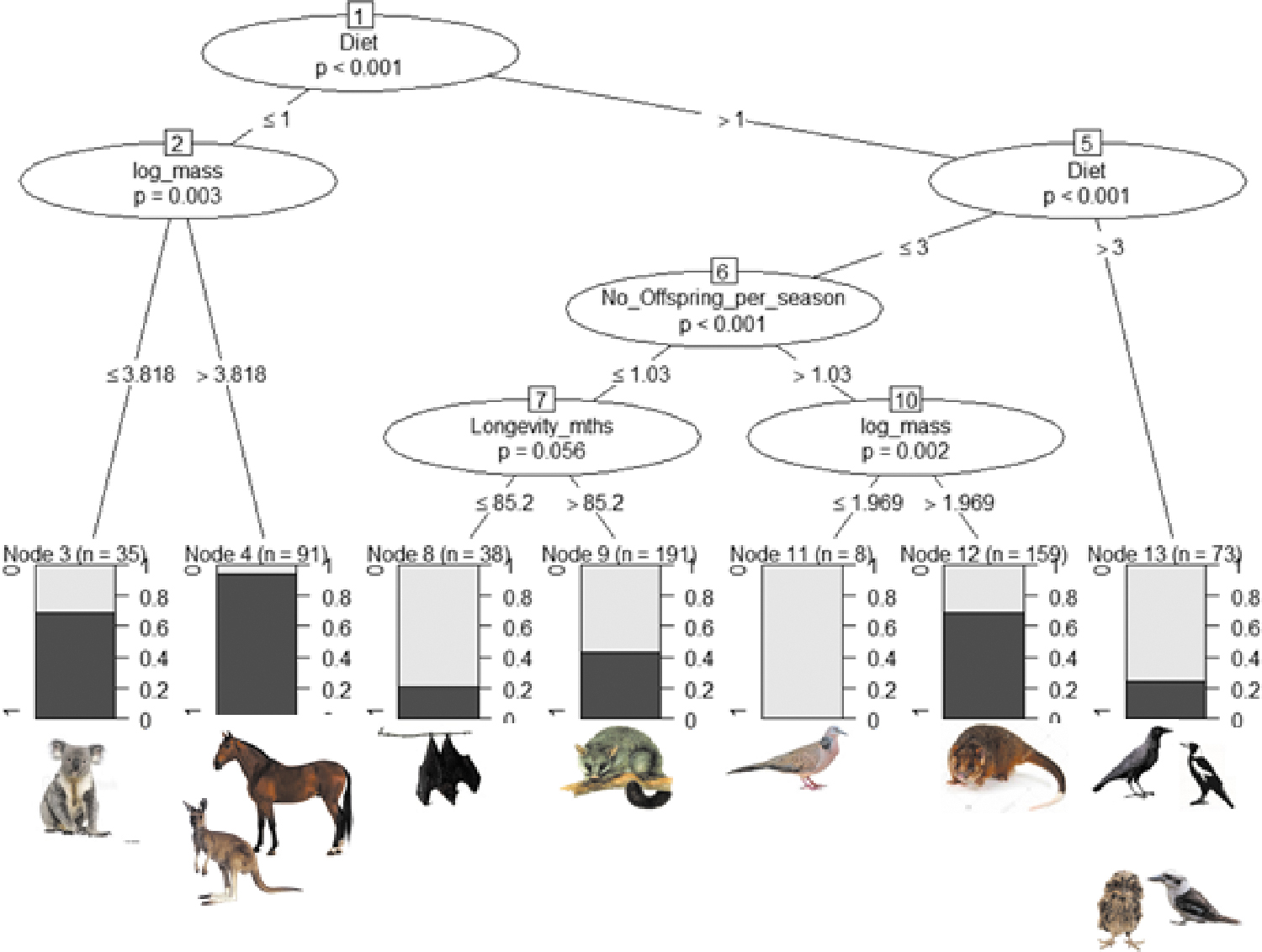
Conditional inference tree based on the six traits most strongly associated with seropositivity (as determined by generalized linear models). The numbered ellipses identify the nodes and variables that best split the data for seropositivity at each point in the tree, with Bonferroni adjusted p values shown together with the cutoff values for the split along the branches. At the bottom of the tree, shading in the boxes representing the terminal nodes shows the proportion of seropositive individuals (darkest shading), and n = the number of individuals in each of the final groups. Although social group size and latitude were included in the analysis, they did not generate a significant split between seropositive or seronegative individuals. Color images are available online.
There was a highly significant difference in seropositivity between ringtail possums Pseudocheirus peregrinus and brushtail possums Trichosurus vulpecula (x 2 = 26.276, p < 0.001), with the odds of a ringtail possum being seropositive 3.2 times higher (CI 2.02–5.34) than for a brushtail possum being seropositive.
Spatial analysis
Point maps showing seropositive and negative results were derived for ringtail possums, brushtail possums, koalas, horses, flying foxes, and birds, separately (Fig. 3a). No strong spatial pattern was observed for the three marsupial species (ringtail possums, brushtail possums, and koalas). Conversely, horses, flying foxes, and birds had a greater aggregation of negative samples in northern SEQ and a greater aggregation of positive samples in southern SEQ (Fig. 3a). This trend was more apparent in the hotspot analysis, which identified a significant coldspot (99% confidence) of RRV seropositivity in the northern SEQ locations, and a significant hotspot (99% confidence) formed around southern Brisbane (Fig. 3b).
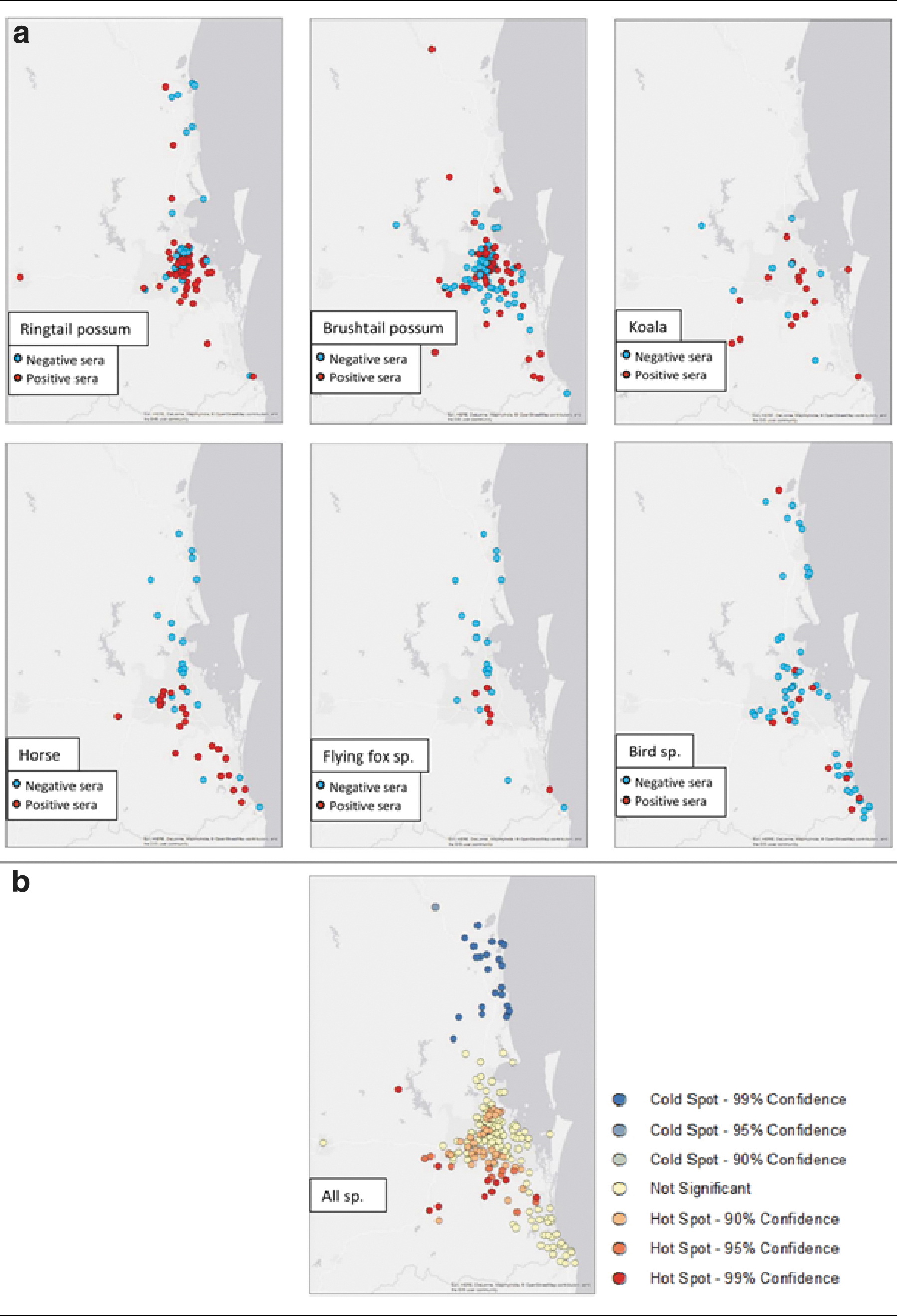
Maps of South East Queensland representing locations and results of samples tested for RRV. Specifically, the
Discussion
RRV is Australia's most epidemiologically important arbovirus and effectively managing transmission requires an understanding of the natural exposure rates of RRV in nonhuman vertebrates. This study tested a greater diversity of vertebrate species for RRV seroconversion than previous studies, using a plaque reduction neutralization assay (PRNT50), which is recognized as the gold standard for arboviruses (Hobson-Peters 2012). We found that placental mammals as a group had higher seropositivity than marsupials or birds. However, these aggregated results may be misleading as there is high heterogeneity of RRV seropositivity within each taxonomic group, indicating that these groupings may not be useful predictors of RRV seropositivity.
Rather than taxonomic relatedness, ecological and life-history traits could be driving some heterogeneity in seroprevalence results. In particular, ringtail possums had a much higher seroprevalence against RRV than brushtail possums, despite both being marsupials. This is interesting because brushtail possums have long been implicated as an important urban reservoir of RRV (Boyd et al. 2001, Hill et al. 2009), and by comparison, ringtail possums have been greatly understudied for RRV transmission (Gard et al. 1973, McManus and Marshall 1986). Brushtail possums and ringtail possums are both abundant in Brisbane and other Australian urban areas. They differ somewhat ecologically: brushtail possums primarily inhabit roofs in urban areas, whereas ringtail possums build nests in trees (Inions et al. 1989, Statham and Statham 1997). These differences could bring ringtail possums into contact with mosquitoes more frequently than brushtail possums, but more studies, including mosquito bloodmeal analyses, would be needed to test this hypothesis.
Vector-host interactions are critical for the transmission of arboviruses and are influenced by a number of factors. Mosquito host-seeking behavior is predominantly guided by detection of heat and carbon dioxide (CO2), but can also be influenced by various odorants and chemicals that are released by hosts, host abundance, and host defensive behavior (Takken and Verhulst 2013, Stephenson et al. 2019). In the conditional inference tree analysis, “diet” was the first characteristic influencing an individual's likelihood of being seropositive or seronegative. One possible explanation for this is that diet is closely linked with CO2 production (Gillies and Wilkes 1972). In general, large-bodied herbivores produce larger quantities of CO2, in part, due to the rumination and fermentation process required to digest their meal, than carnivorous or omnivorous species (Passey et al. 2005). In this study, species with herbivore diets (category 1) were split from other species and had a higher than average seroprevalence. Within this group, the larger bodied vertebrates (horse) had the highest seroprevalence, which could be due to a larger body surface area and heat for mosquitoes to detect and feed on (Kleiber 1947, Franz et al. 2010). By comparison, species with the greatest protein intake (in this case, insectivorous and carnivorous birds) had a lower than average seroprevalence, which may be a result of smaller body sizes or exhibiting host behaviors, which may include feeding on mosquitoes (Torr et al. 2006). However, further research is needed to determine if there are direct links between diet and RRV infection in nonhuman species.
Although horses are assumed to be the only species other than humans to have clinical signs to RRV (Azuolas 1998), this has been based largely on prospective case studies, as opposed to experimental infection results (Kay et al. 1987, Barton and Bielefeldt-Ohmann 2017). Regardless, the high seroprevalence in horses in our study echo that of other published seroprevalence studies (Azuolas 1997, Gummow et al. 2018) and is of interest for several reasons. First, the individuals in this study had no previous sign of RRV infection within the past 12 months, suggesting either that infections were asymptomatic (or sufficiently so to be unobserved by their owners and veterinarian) or that infections occurred more than 12 months before sample collection, indicating that RRV antibodies may be long lasting in horses. Long-term immunity for RRV in horses requires further investigation, given that early experimental studies demonstrate the hemagglutination-inhibition response lasting ∼100 days (Kay et al. 1987), whereas anecdotal reports from horse racing in Australia have found detectable IgG more than 12 years postoriginal infection. * Second, while naive horses have a demonstrated ability to transmit RRV to mosquitoes when experimentally infected (Kay et al. 1986), the high herd immunity reported in this study and in other studies may suggest that they could reduce ongoing transmission through a dilution effect if infected mosquitoes bite previously exposed, and immune, horses. Having a large body size, and limited host defensive strategies, horses are an ideal target for mosquitoes, which is reflected in their frequent detection in blood feeding studies (Stephenson et al. 2019). The duration of RRV antibodies in horses and the association between antibody levels and immunity against reinfection are therefore critical areas for further research to assess the role of horses in maintaining RRV in periurban environments.
Differences in immune responses and abilities of producing neutralizing antibodies among vertebrate species might influence serological findings across the species studied in this analysis. For example, no experimentally infected little corellas seroconverted, despite 50% developing detectable viremias and being capable of infecting susceptible mosquitoes (Kay et al. 1986). Similarly, experimentally infected gray-headed flying foxes showed highly variable immune responses, and were capable of transmitting RRV to susceptible mosquitoes despite undetectable viremias (Ryan et al. 1997). As such, the immune response and assay sensitivity need to be assessed on a species by species basis to fully evaluate whether seroprevalences observed in this study are a true reflection of RRV exposure under natural conditions. To date, no long-term studies for the persistence of RRV antibodies in nonhuman vertebrates have been done. Such studies would be critical in quantifying the variability and detectability of RRV in nonhuman vertebrates.
Human notification patterns of RRV share both similarities and inconsistencies with the results of the hotspot analysis. Consistent with our results, a south-eastern hotspot in Brisbane has previously been identified as a hotspot of human infections over the 1991–1996 time period (Muhar et al. 2000), and again in 2001 (Hu et al. 2007). This could suggest the area experiences frequent spill-over from reservoir species and could be of interest for management interventions. In contrast, more recent data on human notifications in SEQ (from 2018) identify the Sunshine Coast (a coldspot in our analysis) as consistently reporting the highest notification rates (number of notifications/human population) compared to metropolitan Brisbane and Gold Coast (Australian Government 2018). Human notification data differ from the seroprevalence data presented in this study, in that notifications occur closer to the timing of an infection, and only represent those infections that were associated with clinical signs of RRV in humans (as opposed to asymptomatic infections). This, in combination with the complicated disease ecology of RRV (involving multiple vectors and vertebrate hosts), may suggest that transmission patterns between humans and nonhuman vertebrates may vary and/or be highly localized (Claflin and Webb 2015, Flies et al. 2018). However, while the hotspots and coldspots were identified using an objective statistical algorithm, it is important to note that because the sampling method used in this study was opportunistic and not a random survey, there is the potential for sampling bias.
Ecological applications
Our findings differ to a recent evaluation of ecological characteristics in seropositive species (Walsh 2019), which noted that diet breadth, population density, and gestation period were factors associated with seropositivity. However, our study differs in that we use a continuous measure of seropositivity (rather than a binary “positive” or “negative” response variable), include nonmammal species, and use empirical data from a single location and time period (rather than a literature review at an aggregated national and historical level) to reach a different conclusion. We strongly believe that it is important for both studies to be considered in any future work.
The findings from this research have practical applications for targeted management of RRV. For example, if there are high levels of exposure identified in a given species that is known to amplify RRV, mosquito control measures can be put in place to reduce mosquito-host contact rates. The output of the results in a decision tree makes it easy to visualize which populations are best to target. A limitation with this method is that the results are only applicable to the species and life history traits that are included in the analysis. While our study has assessed the greatest diversity of vertebrate species tested for RRV, it does not reflect the full biodiversity of species in SEQ and there may be other important life history traits that we have not yet considered. However, the framework will be a valuable addition to future studies incorporating more species and to assess the outcomes of management efforts when used over a temporal scale.
Future directions
Serosurveys provide an informative tool to understand patterns of transmission; however, for vector-borne diseases, they are limited in identifying reservoir hosts. An individual who has a detectable immune response to RRV has previously been infected, but this does not correlate to an acute infection capable of amplification. Few experimental infection studies have been undertaken to determine which vertebrates can infect susceptible mosquitoes with RRV, and without this information, it is unclear whether or not the seropositive species identified in this study are amplifying or diluting RRV in this complex transmission system. Experimental infection studies that utilize multiple vector species to infect and measure infectivity of vertebrates with RRV are needed to fill this gap.
Equally important is long-term serological sampling of a community. For WNV, the use of localized studies that incorporated (1) vector availability, (2) host abundance, (3) host exposure rates, and (4) human infections was successful in identifying migratory bird species as a key reservoir (Kilpatrick et al. 2006). For RRV, there are no published studies that take repeated samples from a community. Repeated sampling from a free-living community in combination with climatic and vector variables would provide novel insights into the local transmission dynamics of RRV.
Conclusion
Seroprevalence data provide critical information about the transmission and distribution of pathogens in nonhuman species. In this study, we interpret the RRV seroprevalence data across the greatest diversity of species in a single study, using a novel modeling methodology. We confirm that nonhuman vertebrates are frequently infected with RRV across SEQ. The highest seroprevalence was reported in horses, followed by koalas and ringtail possums. Species traits, in particular diet and body mass, may play a role in vector-host contact rates, ultimately influencing exposure and ongoing transmission. Ringtail possums seroprevalence was significantly higher than brushtail possums, which is particularly interesting as only the latter have been investigated as potential reservoirs for RRV. Further research is needed, encompassing experimental infection studies and long-term serosurvey studies to identify the transmission dynamics between vectors and hosts. The results from this study provide a framework to examine serological results and ultimately inform management of pathogens more broadly.
Footnotes
Authors Contributions
E.B.S. collected all samples; E.B.S., P.A.R., and L.J.H. undertook all laboratory analyses; E.B.S., A.J.P., and H.M. conducted modeling analyses. All authors contributed to the conceptualization of the study, interpretation of results, and writing and editing of the article.
Ethical Approval
This work was performed in accordance with Griffith University Animal Ethics Committee standards (approval number: ENV/04/17/AEC).
Data Accessibility
Data are currently accessible through the CloudStor Repository using the link below; once published, data will be uploaded to figshare:
Acknowledgments
We would like to acknowledge the following veterinary clinics for contributing to this research: RSPCA Wildlife Hospital, Wacol; Currumbin Wildlife Sanctuary, Currumbin; Australia Zoo Wildlife Hospital, Beerwah; Brisbane Bird Vets, Chermside; WestVets, Anstead; and Gold Coast Equine Clinic, Bundall. Cassie Jansen is a supervisor of Eloise Stephenson and contributed to the project design. We would also like to acknowledge Narayan Gyawali from QIMR for providing the serology protocol. We would like to broadly acknowledge the L.J.H.'s and H.M.'s laboratory members for their guidance and revisions of this article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
L.J.H. is the recipient of the Australian National Health and Medical Research Council Career Development Award (ID: 105760). A.J.P. was supported by a Queensland Government Accelerate Postdoctoral Research Fellowship. E.B.S. is the recipient of an Australian Government Research Training Program. Sample collection was supported by a Student Research Award from the Ecological Society of Australia, and a Future Fellow award from Graduate Women Gold Coast.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.