
Abstract
Parasitic diarrheal disease is a major cause of morbidity and mortality in the developing world. Calves are highly susceptible to Cryptosporidium spp. infection that resulted in diarrhea, growth retardation, and weight loss, and was one of the most common enteropathogens. It is especially difficult for molecular detection of calves with inapparent or subclinical infections of cryptosporidiosis. In view of this, this study established a real-time quantitative PCR (RT-qPCR) detection method to clarify its epidemic characteristics, based on Cryptosporidium 18S rRNA gene with the 150 bp product length to investigate the infection of Cryptosporidium spp. in northeastern China The standard curve equation is Ct = −2.91 × lg (Cryptosporidium spp. copies) +10.18, with better sensitivity, stability, and reproducibility. A total of 148 out of 425 fecal samples (34.82%) were detected Cryptosporidium positive with RT-qPCR, including (36.11%) in Heilongjiang province (29.60%), (29.6%) in Jilin province, and (37.50%) in Liaoning province. The infection prevalence of Cryptosporidium parvum, Cryptosporidium ryanae, Cryptosporidium andersoni, and Cryptosporidium bovis from calves in order from high to low was 14.35% (95% confidence interval [CI], 11.2–18.1), 6.12 (95% CI, 4.0–8.8), 2.35 (95% CI, 1.1–4.3), and 0.47 (95% CI, 0.1–1.7), respectively, suggesting C. parvum was the predominant species in calves in northeastern China. Using 60-kDa glycoprotein gp60 gene, all of the 61 C. parvum-positive specimens were further precisely confirmed to IIdA19G1 subtype. This suggested that IIdA19G1 subtype of C. parvum could threaten to cause diarrhea calves from notheastern China (p < 0.01). The prevalence of 34.82% (148/425) using RT-qPCR had a significant difference compared with the prevalence of nested-PCR (23.29%) and microscopic examination (3.76%). The findings improved the epidemiological knowledge of calves infected with cryptosporidiosis in China, highlighting the importance of ongoing Cryptosporidium surveillance.
Introduction
Cryptosporidiosis infected by Cryptosporidium spp. is a global and opportunistic protozoan disease of which the main clinical features include persistent diarrhea. Presently it can infect humans and many animals in >40 countries around the world (Ahamed et al. 2015). Cryptosporidium can cause severe persistent diarrhea in young ruminant mammals, and result in the catarrhal respiratory symptoms in poultry, and gastritis in fish and amphibians (Ahmed et al. 2015). Moreover, cryptosporidiosis is a key factor that causes diarrhea in children and immunodeficiency in patients (Mirhashemi et al. 2016).
Especially, long-lasting and intense infection leads to severe watery diarrhea, gastroenteritis, and dehydration in immunocompromised animals, and is even life threatening for the patients with low immune function, such as infants and AIDS patients (Ondrackova et al. 2009). In recent years, the large outbreak of cryptosporidiosis has been reported in the world, and its harmfulness has gradually attracted great attention (Caccio and Chalmers 2016). Until 2006, cryptosporidiosis has been classified as a neglected disease initiative by the World Health Organization (Chavez and White 2018). Now, effective and therapeutic counter measures for cryptosporidiosis remain absent (Ryan et al. 2014). This manifests the requirement of epidemiological data in illuminating the transmission dynamics and in planning and implementing prevention strategies (Xiao 2010).
Since the first case of Cryptosporidium infection in cattle in the United States was known, cryptosporidiosis has developed into one of the most important causes of diarrhea in cattle (Wang et al. 2017). Serious diarrhea, weight loss, growth stagnation, and even death are infected by Cryptosporidium spp. in newborn calves, endangering the healthy development of cattle industry, and bringing a huge threat to public health and safety. Two kinds of important sources of human infection include the ingestion of contaminated food or polluted water by tainted feces and the fecal-oral pathway of Cryptosporidium spp. (Xiao 2010). Human clinical presentations have been significantly associated with virulence variations of Cryptosporidium species and subtypes on DNA polymorphisms of the small subunit (SSU) rRNA gene (Cama et al. 2008). At present, there are 37 Cryptosporidium species and >70 genotypes. And the genotyping, subtyping, and population genetic studies increasingly exhibit the genetic diversity (Ryan et al. 2014, Ouakli et al. 2018). Four Cryptosporidium species were common in cattle, referring to Cryptosporidium parvum, Cryptosporidium bovis, Cryptosporidium Andersoni,and Cryptosporidium ryanae, whereas C. andersoni and C. bovis have occasionally been reported in humans (Ryan et al. 2014). In addition, according to literature reports, Cryptosporidium hominis, Cryptosporidium meleagridis, Cryptosporidium ubiquitum, Cryptosporidium suis, Cryptosporidium scrofarum, and Cryptosporidium felis have been found in cattle (Ouakli et al. 2018).
Detection of Cryptosporidium spp. is important for epidemiological surveys, disease surveillance, monitoring, and identifying the causation of diarrhea in clinical calves. Detection of Cryptosporidium spp. DNA or antigens and oocysts visualization are the cornerstone of diagnosis of cryptosporidiosis (Striepen 2013). Therefore, microscopic, molecular, and immunological methods have been developed to detect Cryptosporidium spp. It is difficult to adopt the requirement of clinical specimen because of the presence of other oocyst-like structures, yeast in fecal debris, and poor specificity of microscopic method (Striepen 2013). Thus, molecular techniques have replaced staining for the diagnosis of cryptosporidiosis. Target genes to commonly detect Cryptosporidium genotyping currently include SSU ribosomal RNA (SSU rRNA), Cryptosporidium oocyst wall protein (COWP) gene, heat shock protein 70 (HSP70) gene, etc. (Ahmed et al. 2015).
With the development of molecular biological detection technology, PCR technology has become the basis of developing a new generation of diagnostic methods. At present, many PCR diagnostic methods for Cryptosporidium detection are used to check clinical samples and environmental samples. Nested PCR is widely used to detect infection of Cryptosporidium in humans and animals (Avendano et al. 2018). However, this method failed to PCR products with dark electrophoretic bands or asymptomatic animals with shedding of few oocysts per gram of feces. Real-time PCR (RT-qPCR) detection method has also been widely used in pathogen detection in recent years (Stark et al. 2011). Compared with the common PCR method, this method has higher sensitivity and specificity, and can achieve relative quantization of Cryptosporidium oocysts in feces (Stark et al. 2011).
Sensitive and specific RT-qPCR method is needed to obtain exact epidemiological data in calves. The aims of this study were to (1) establish a new RT-qPCR detection method for investigating the epidemiology of Cryptosporidium spp.; (2) grasp the positive rate and distribution of Cryptosporidium in calves in northeastern China; (3) compare the difference of prevalence using three methods, such as RT-qPCR, nested PCR, and microscopic examination; and (4) determine the species/genotypes of Cryptosporidium. In short, this study is to grasp the infection and epidemic characteristics of Cryptosporidium spp. in calves in some areas of northeastern China through establishing a new RT-qPCR method, which could help us to understand the potential harm and transmission dynamics of Cryptosporidium spp. between animals and humans, and to provide basic data for prevention and control for calves in northeastern China.
Materials and Method
Specimen collection and preparation
The fresh fecal samples from 425 diarrhea or nondiarrhea calves were directly obtained from three different provinces, northeastern China from December 2016 to December 2018, referring to 108 samples from Heilongjiang Province (longitude 121°11′–135°05′, latitude 43°26′–53°33′), 125 samples from Jilin Province (longitude 121°38′–131°19′, latitude 40°52′–46°18′), and 192 samples from Liaoning Province (longitude118°53′–125°46′, latitude 38°43′–43°26′) (Fig. 1). The four seasons from spring to winter were defined as January–March (n = 96), April–June (n = 101), July–September (n = 107), and October–December (n = 121), respectively. Each calf was sampled once in a cowshed. The fecal samples were collected using sterile spoons, then taken into sterile gloves and transported back to the laboratory to be stored in 4°C.
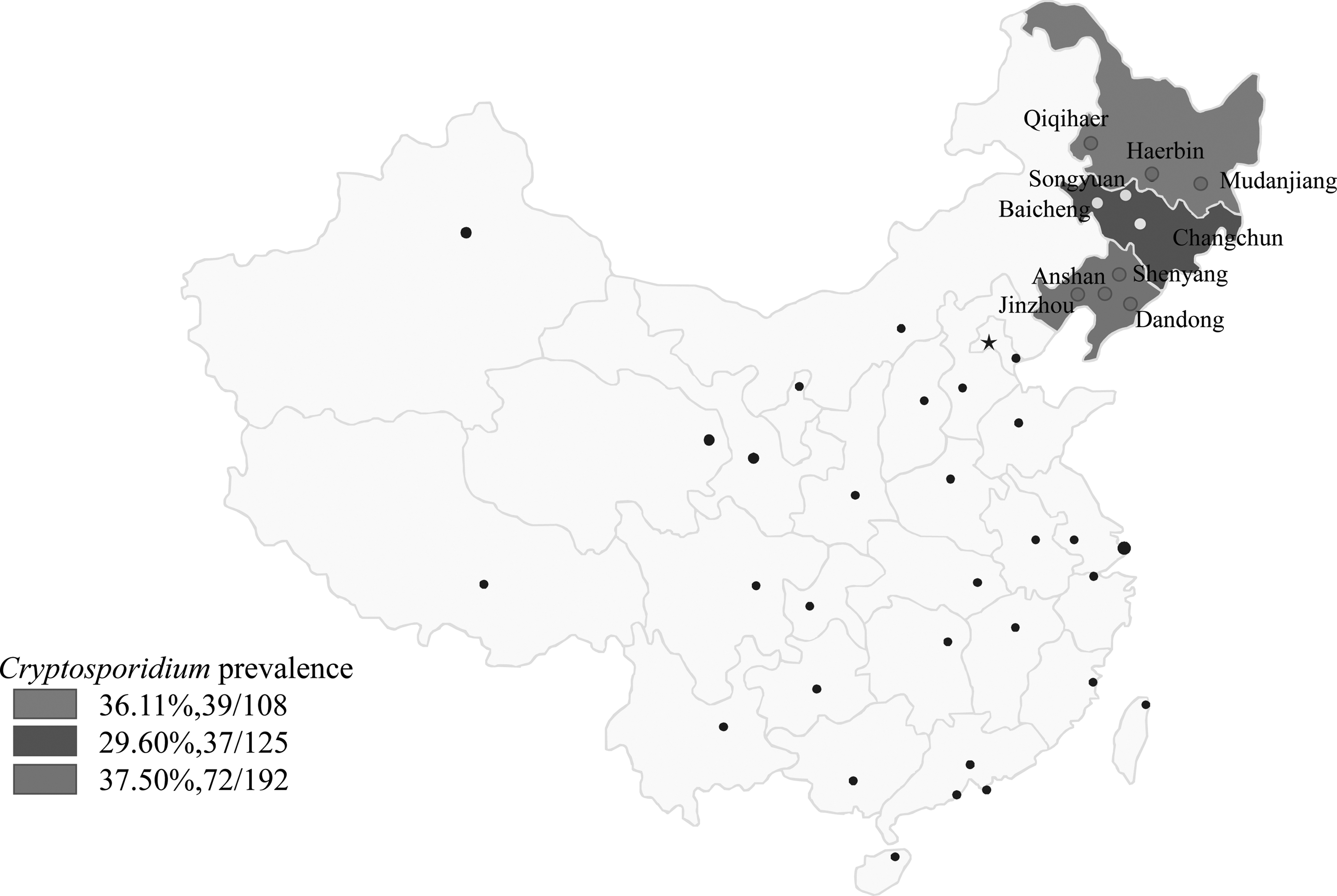
Location of positive samples obtained from northeastern China with RT-qPCR.
Microscopic examination of feces for Crytoporidium oocyts
After using the discontinuous sucrose gradient flotation technique, 10 μL of sediment was made a smear (Karanis et al. 2007). The smears were air dried, heat fixed, and stained according to the Kinyoun method that was an acid-fast procedure to the species of Cryptosporidium spp. (Kehl et al. 1995). After drying and fixing with the cover glasses, the smears were observed using the oil immersion objective under a Nikon Eclipse E 200 light microscope (Nikon, Japan). Positive samples showed round or oval isolated structures (4–6 μm) with pink to deep purple.
Primer design and optimization of RT-qPCR
Primers of Cryptosporidium species were designed using specific software (Beacon Designer 8.0), according to 18S rRNA gene from Cryptosporidium spp., available in Genbank. The characteristics and specificity of primers were tested and evaluated using Primer Premier 5.0, prime rBLAST, NCBI, Bethesda, and MD. The optimized primers amplifying a 150 bp fragment of 18S rRNA gene of Cryptosporidium spp., (Table 1) were commercially synthesized by Integrated DNA Technologies (Coralville, IA). The positive control DNA as a template was used to generate concentration curves and optimize the primer. The positive DNA sample of C. parvum was obtained from a symptomatic calf and verified by RT-qPCR targeting the SSU of the nuclear ribosomal RNA (SSUr RNA) gene. The annealing temperatures for individual RT-qPCR assays were first optimized, with three replicates in each degree using the ABI 7500 Fast Real-Time PCR System (Applied Biosystems, Foster City, CA). According to the results of the RT-qPCR assay, the best annealing temperature was selected. RT-qPCRs consisted of 9.5 μL PCR-grade water, 12.5 μL TB Green II PCR Master Mix, 1 μL each of the forward and reverse primers at 20 μM, and 1 μL Cryptosporidium spp. (100 ng/μL) DNA sample.
Primer Sequences and Amplification Protocol of RT-qPCR
Plasmid construction
The purified PCR product of 830 bp with SSUr RNA gene of C. parvum was cloned into pGEM-T vector (Promega) and then propagated in Escherichia coli DH5a competent cells. The recombinant plasmid was purified using EasyPure Plasmid MiniPrep Kit (TransGen Biotech, Beijing, China) and sequenced. The number of copies of the plasmid (copies/μL) was calculated through the concentration of the extracted plasmid using a NanoDrop 2000 system (Thermo Fisher Scientific). Serial 10-fold dilutions of the standard plasmid DNA template from 101 to 106 were made and frozen at −20°C.
Analytical sensitivity, specificity, and reproducibility of RT-qPCR
Serial decimal dilutions of the amplicon that has been cloned into plasmid were used to evaluate the analytical sensitivity. The sensitivity of the RT-qPCR was measured by the plasmid with number of copies versus Cq values. And 10-fold serial dilutions of plasmid at concentrations ranging from 1 × 10 to 1 × 106 per μL were subjected to RT-qPCR analysis, and a standard curve was generated for each individual assay with six separate dilution series performed for each point of the curve in triplicate,. The linear regression, along with the determination of correlation coefficient (R 2) formed after determination of each point of the curve, can be used to evaluate whether RT-qPCR assay has been optimized. Each reaction's efficiency was determined through the slope of standard curve using the following formula: efficiency = 10 (−1/slope) −1. Specificity of the assay was assessed by DNA sample of E. coli, Eimeria tenalla, and Toxoplasma gondii. Nuclease-free water was used as a negative control. Tenfold dilutions of the standard plasmid DNA from 102 to 104 copies were used to evaluate the reproducibility with coefficient of variation (CV) of cycle threshold values, as previously described (Xu et al. 2011). After being optimized, the final selected cycling profiles were as follows: initial denaturation at 95°C for 30 s, 40 cycles at 95°C for 15 s, annealing temperatures 60°C for 30 s. To ensure the specificity of SYBR Green PCR amplification, a melting curve analysis program was incorporated as follows: 95°C for 15 s, 60°C for 30 s, and 95°C for 15 s. A curve of dissociation was generated to verify the specificity of the amplifications.
Nested PCR
Genomic DNA of all fecal samples was extracted from feces using the Stool Genomic DNA kit (ComWin, Beijing, China), according to the manufacture's recommendations (Liang et al. 2019). Aliquots (2 μL) of individual genomic DNA samples were subjected to nested PCR-based amplification of the SSUr RNA gene (830 bp) of Cryptosporidium spp., and sequencing by employing (individually) two distinct loci of nuclear ribosomal DNA in separate PCR assay systems consisting of 1 × PCR buffer, 10 μM of each primer (the primary amplification primers SSU rRNA F1 (5′-3′ CCCATTTCCTTCGAAACAGGA) and SSU rRNA R1 (5′-3′ TTCTAGAGCTAATACATGCG) as outer primer, and SSU rRNA F2 (5′-3′ AAGGAGTAAGGAACAACCTCCA) and SSU rRNA R2 (5′-3′GGAAGGGTTGTATTATTAGATAAAG) as inner primer (Zhao et al. 2013).
And further genotypic/subgenotypic classification of C. parvum was achieved by 60-kDa glycoprotein gp60 gene (Ahmed et al. 2015). The amplification product was analyzed directly by 1.5% agarose gel electrophoresis with ethidium bromide solution (1 μg/mL). Positive control of sequenced DNA sample and negative control of sterile water were included in each PCR: PCR (25 μL) was composed of 1 × Ex Taq buffer (Mg2+free), 2 mM MgCl2, 200 μM each deoxy-ribonucleoside triphosphate, 0.4 μM of each primer, 0.625 U of Ex Taq DNA polymerase (TAKARA, Japan), and 2 μL of DNA template. The cycling conditions were 5 min at 95°C, 35 cycles of 45 s at 94°C, 45 s at suitable temperature (Table 1), and 1 min at 72°C, followed by final extension at 72°C for 5 min.
The sensitivity of the nested PCR was analyzed by the plasmid concentrations with 10-fold serial dilutions of plasmid at concentrations ranging from 1 × 10 to 1 × 106/μL using 1.0% of agarose gel electrophoresis.
Sequencing and phylogenetic analysis
After amplification of the targets, the PCR products of all positive samples were sequenced by the Sanger method in an ABI 3730 sequencer (Comate Bio, Changchun, China) using bidirectional sequencing. The obtained sequences were analyzed by BlastN and ClustalW (Thompson et al. 1994), and were compared with the sequences of different genospecies. The four main sequences of infecting calves were added in phylogenetic tree through the software MEGA 5.0, such as C. parvum, C. bovis, C. andersoni, and C. ryanae. The neighbor-joining method was employed to construct a phylogenetic tree. The reliability of branches in the tree was evaluated by bootstrapping analysis with 1000 replicates, and the bootstrap value >75% was considered significant.
Statistical analysis
The prevalence of Cryptosporidium spp. infection in calves was calculated using SPSS version 17.0 (SPSS, Inc., Chicago, IL) with 95% confidence interval (CI) with one-sample analysis. The variation in Cryptosporidium prevalence (y) of calves of different geographical location (x1), age (x2), and season group (x3) was analyzed by χ2 test using SAS version 9.1 (SAS Institute, Inc.). The best model was judged by Fisher's scoring algorithm. All tests were two sided, and values of p < 0.05 were considered statistically significant. And their 95% CI was estimated to explore the strength of the association between Cryptosporidium positivity and the conditions investigated.
Results
Concentration of standard plasmid DNA from C. parvum
The concentration of standard plasmid DNA from Cryptosporidium spp. Was 173.91 ng/μL, and the number of copies of this plasmid DNA was 1.59 × 1011 copies/μL.
Establishment of the RT-qPCR standard curve
Plasmid with known number of copies was diluted serially in concentrations from 1.59 × 101copies/μL to 1.59 × 106 copies/μL for RT-qPCR. The linear correlation between the Ct value and logarithm of plasmid with number of copies is 0.997 (R 2 = 0.997) and efficiency was 1.19, respectively. Based on the standard curve equations: Ct = −2.91 × l g (the number of Cryptosporidium spp. copies) +10.18 (Fig. 2).
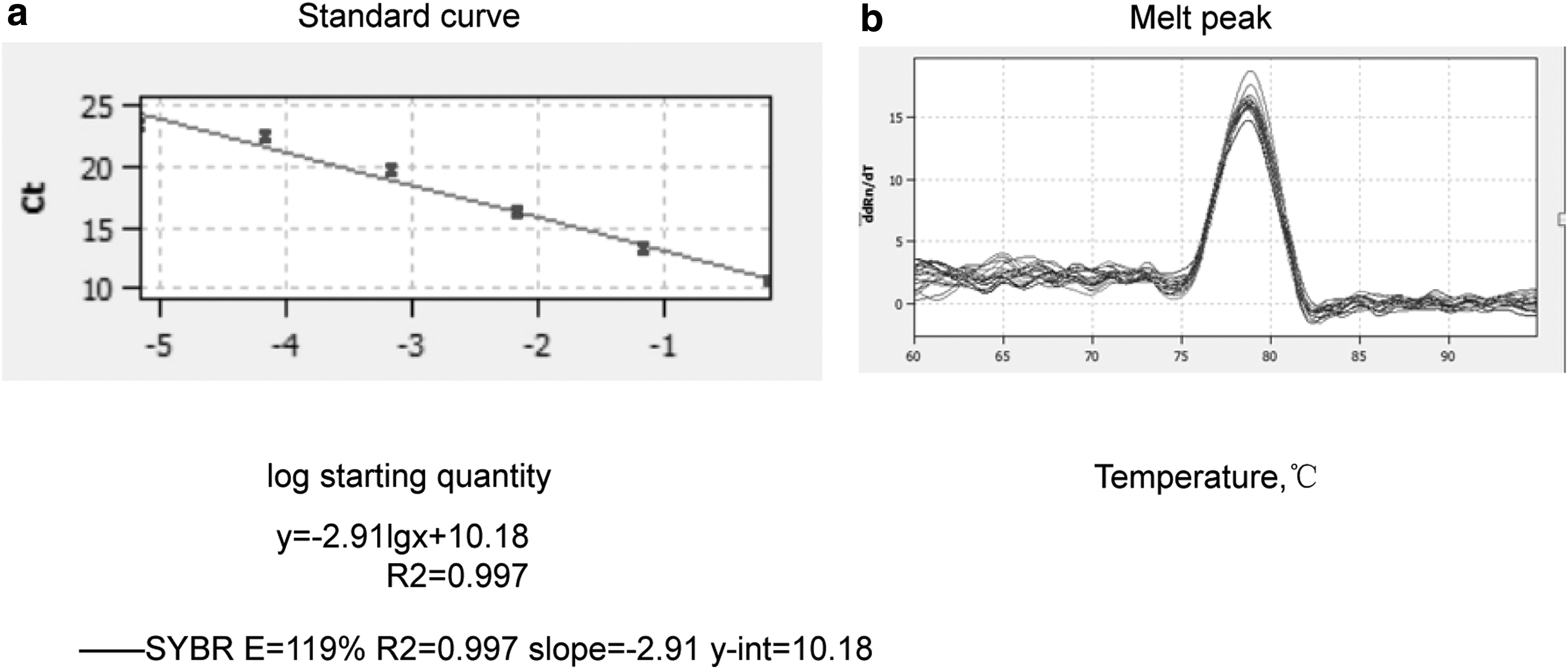
Standard curve and melt peak results of SYBR Green II real-time PCR.
Analytical sensitivity, specificity, and reproducibility of RT-qPCR
According to the result of the agarose gel electrophoresis, the lowest detection limit of the assay is 1.59 × 101 copies/μL (Fig. 3a); however, nested PCR only detected 1.59 × 102 copies/μL. Both intra- and interassay reproducibility were calculated through 10-fold serial dilution of standard plasmid DNA, ranging from 1.59 × 101 copies/μL to 1.59 × 106 copies/μL (Fig. 3b). CV was ranging between 0.36% and 1.86%, suggesting the RT-qPCR had high reproducibility. CV values of intra- and interassay were ranging from 1.33% to 1.86% and from 0.36% to 0.94%, respectively. And neither DNA templates from E. coli, E. tenalla, and T. gondii nor negative control generated the detectable fluorescence signals (Fig. 3c), suggesting that the established RT-qPCR assay was specific to Cryptosporidium spp.

Detection of Cryptosporidium spp. in the calves from northeastern China using RT-qPCR
A total of 148 out of 425 fecal samples (34.82%, 95% CI 30.3–39.6%) were detected Cryptosporidium positive with RT-qPCR, such as 36.11% (95% CI 27.17–45.9) in Heilongjiang province, 29.60% (95% CI 21.8–38.4) in Jilin province, and 37.50% (95% CI 30.6–44.8) in Liaoning province (Table 2). The results of some positive samples were shown with 2.5% agarose gel electrophoresis (Fig. 4). The prevalence of Cryptosporidium spp. had statistical difference between diarrhea (48.29%, 95% CI 13.1–24.6) and nondiarrhea calves (18.32%, 95% CI 41.7–54.9). In different seasons, the prevalence ranged from 29.17% (95% CI 20.3–39.3) to 41.12% (95% CI 31.7–51.0) (Table 3). Fisher's scoring algorithm showed significant symptom-, age-, and gender-associated difference in positive Cryptosporidium calves (p < 0.01, Table 3).

Results of real-time amplification of Cryptosporidium spp. in calves. M, DNA MarkerDL1000; 1–12, sample number.
Detection of Cryptosporidium DNA by RT-qPCR from Heilongjiang, Jilin, and Liaoning Provinces, Northeastern China
Prevalence of Cryptosporidium infection was calculated using SPSS version 17.0 with 95% CI with one-sample analysis.
CI, confidence intervals; nd, no data of this area.
Prevalence of Cryptosporidium Infection of Calves in Heilongjiang Province, Jilin Province, and Liaoning Province
Prevalence of Cryptosporidium infection was calculated using SPSS version 17.0 with 95% CI with one-sample analysis.
Distribution of Cryptosporidium species/subtypes and phylogenetic analyses
A total of 99 samples (23.29%) out of 148 real-time positive samples were only detected and sequenced by nested PCR with SSUr RNA gene (830 bp) of Cryptosporidium. After BLAST on the NCBI website, sequences in the GenBank database, most similar to the query sequences, were retrieved and used for phylogenetic analysis. Our result revealed that four Cryptosporidium spp. such as C. parvum, C. ryanae, C. andersoni, and C. bovis were obtained from 10 cities in Heilongjiang, Jilin, and Liaoning provinces (GenBank accession numbers: MT374186–MT374189).
The infection prevalence of C. parvum, C. ryanae, C. andersoni, and C. bovis from calves in order from high to low was 14.35% (95% CI, 11.2–18.1), 6.12 (95% CI, 4.0–8.8), 2.35 (95% CI, 1.1–4.3), and 0.47 (95% CI, 0.1–1.7), respectively. This result suggested that C. parvum was the predominant species in calves in northeastern China. Compared separately with three provinces, C. parvum is the predominant species in calves in Heilongjiang (18.52%, 95% CI 11.7–27.1) (p < 0.01) and Liaoning province (20.83%, 95% CI 15.3–27.3) (p < 0.01). Nonetheless, C. bovis (8.00%, 95% CI 3.9–14.2) and C. andersoni (8.00%, 95% CI 3.9–14.2) were predominant species in Jilin province (p < 0.01) (Table 4). Moreover, the distribution of Cryptosporidium spp. showed the obvious regional differences. For example, C. andersoni was only detected in Jilin province, and C. ryanae was only found in Liaoning province. Altogether, all the 61 C. parvum-positive specimens were further precisely confirmed as IIdA19G1 subtypes using 60-kDa glycoprotein gp60 gene (Alves et al. 2003).
Different Prevalence of Cryptosporidium Infection of Calves with Microscopy, Nested PCR, and RT-qPCR
Prevalence of Cryptosporidium infection was calculated using SPSS version 17.0 with 95% CI with one-sample analysis.
Phylogentic analysis using neighbor-joining method suggested that all Cryptosporidium-positive sequences were clustered into four clades such as C. parvum, C. ryanae, C. andersoni, and C. bovis, which had ∼100% homology with C. parvum in calves from France (Genebank: MK0147750), C. ryanae (Genebank: MF671875) in dairy cattle from China, C. andersoni in cattle from China (Genebank: MH754166), and C. bovis in calves from oocyst (Genebank: MK982466), respectively (Fig. 5).

Phylogenetic relationships (neighbor joining tree-building method) among Cryptosporidium as deduced from structure alignment of the 18S rRNA gene. Samples isolated in this study were compared with isolates selected from GenBank. Giardia intestinalis 18S rRNA gene was used as out group. Numbers at the nodes indicate the bootstrap confidence values obtained after 1000 replicates.
Comparison of the prevalence of microscopic examination, RT-qPCR, and nested PCR
The prevalence of 34.82% (148/425) using RT-qPCR had a significant difference compared with the prevalence of nested-PCR (23.29%) and microscopic examination (3.76%) (Table 5). A significant difference (p < 0.01) was shown with one-way ANOVA.
Detection of Cryptosporidium spp. Prevalence in Calves by Nested PCR in Jilin, Heilongjiang, and Liaoning Provinces, Northeastern China
Prevalence of Cryptosporidium infection was calculated using SPSS version 17.0 with 95% CI with one-sample analysis.
Discussion
Cryptosporidiosis was caused by Cryptosporidium spp., such as C. hominis and C. parvum. And it is one of the most common causative agents of diarrhea in a variety of animal species and humans, especially in children and immunocompromised individuals. In preweaned calves, it was responsible for significant morbidity of diarrhea (Xiao 2010). Thus, the reliable, fast, convenient, and inexpensive tests for precisely identifying Cryptosporidium infection are indispensable to monitor, control, and prevent cryptosporidiosis.
RT-qPCR has improved greatly the detection ability of a pathogen (Ilharreborde et al. 2009). Our study developed a kind of RT-qPCR assay, based on SYBR Green targeting Cryptosporidium spp. 18S rRNA gene for quantitative and diagnostic infection of Cryptosporidium spp. in calves. And its melting curve and Tm value can intuitively analyze whether the product of the reaction is the intended target (Wang et al. 2017).
The advantage of 18S rRNA gene should ideally be able to detect all Cryptosporidium spp, because it covers the major region of interspecies/genotype variability in the gene, allowing identification of nearly all Cryptosporidium spp. and genotypes by sequence analysis of the PCR product while being relatively short to allow efficient amplification and real-time detection (Ahmed et al. 2015). Our RT-qPCR described here targets the 18S rRNA gene to detect all members of the Cryptosporidium genus, which detects Cryptosporidium in clinical samples with better sensitivity and specificity than those of nested PCR. And it decreased the handling times with costs comparable with those of conventional PCR-based methods. Moreover, this assay can detect 1.59 × 101 copies/μL with the lower CV of intra- and interassay between 0.36% and 1.86%. Thus, this result suggested that our detection method had high sensitivity, stability, and reproducibility. Besides, it played the important role in further quantitative study of Cryptosporidium spp. infection.
The results of RT-qPCR showed that the positive rate of Cryptosporidium (34.82%) in calves from northeastern China was within the scope of the prevalence of Cryptosporidium (3.45–47.68%) in cattle from China (Wang et al. 2017). It was lower than that in Xinjiang (38.4%) (Qi et al. 2015) and Heilongjiang (47.68%) province (Zhang et al. 2013) in previous studies, whereas it was higher than Guangdong (24.0%) (Feng et al. 2019), Henan (21.5%) (Wang et al. 2011), Sichuan (14.4%) (Zhong et al. 2018), and Jiangsu provinces (22.7%). Similarly, Cryptosporidium infection in dairy calves elsewhere was observed around the world. The prevalence of Cryptosporidium in our research was considerably lower than that in some countries with high Crytosporidium co-occurrence in cattle, such as in the United Kingdom (80.0%) (Thomson et al. 2017), France (70.4%) (Mammeri et al. 2019), and the United States (96.6%) (Nguyen et al. 2007). A relatively low occurrence of Crytopsporidium was found in Argentina (25.5%) (Lombardelli et al. 2019) and Korea (4.4%) (Lee et al. 2019).
The mentioned research showed that different prevalence of Cryptosporidium infection was related to geographical area, feeding management, weather factor, and detection method in different areas (Tao et al. 2018). The prevalence of Cryptosporidium spp. (54.03%, 95% CI 46.0–61.9) in preweaned calves was significantly higher than those in postweaned calves (23.10%, 95% CI 5.4–8.9), showing a negative correlation between Cryptosporidium infection and age. This result was consistent with the study that the infection rate was decreased with the increase of animal age (Mi et al. 2014). In general, the species of Cryptosporidium infection in cattle are related to age, for example, C. parvum was mainly found in preweaned calves, whereas C. bovis and C. ryanae were detected in postweaned calves (Mi et al. 2013). In this study, the infection rate of Cryptosporidium spp. was significantly different between diarrhea and nondiarrhea calves (p < 0.001), showing that Cryptosporidium spp. may be an important cause of diarrhea in calves in northeastern China. Therefore, the risk of diarrhea in infected calves was significantly higher than that in noninfected calves. This requires us to pay more attention to the management of preweaned calves, including timely isolation from cows, good colostrum feeding, environmental health, etc.
Phylogentic analysis showed that four Cryptosporidium species were found. And the highest infection rate of C. parvum was 14.35% (95% CI, 11.2–18.1), followed by C. ryanae 6.12% (95% CI, 4.0–8.8), C. andersoni 2.35% (95% CI, 1.1–4.3), and C. bovis 0.47% (95% CI, 0.1–1.7) in order, respectively, suggesting C. parvum was the predominant species in calves in northeastern China. This result hinted that C. parvum is the main factor of diarrheal outbreaks in calves from northeastern China. This result was consistent with those of other places around the world. For example, the dominant species of preweaned calves in Algeria with the prevalence of C. parvum (72.4%) (Ouakli et al. 2018) and diarrheic calves in central Argentina with the prevalence of C. parvum (25.5%) (Lombardelli et al. 2019), which was not in accord with the dominant species of Cryptosporidium in two previous reports about Heilongjiang province that C. andersoni and C. bovis were the most common Cryptosporidium species in cattle of all ages (Liu et al. 2009, Zhang et al. 2015 ).
Sample size, sample location, and season factor may contribute to the dominant species of Cryptosporidium (Zhang et al. 2015). First, it may be associated with different cattle farms and sample size in Heilongjiang. Second, seasonal differences also affect the changes of dominant species. Indeed, Wang et al. (2017) have proved that a seasonal shift may be responsible for the dominance of Cryptosporidium species over others in the preweaned calves. Finally, the dominant species in our research was consistent with Tao et al.'s (2018) study in dairy cattle from Helilongjiang province in 2018.
Meanwhile, C. parvum may cross species barriers to infect humans, although most Cryptosporidium species and genotypes are highly specific to hosts (Xiao 2010). In China, C. parvum infection in humans has been found in Henan (2/1366, 0.1%) (Xiao 2010). In European countries, C. parvum are common in humans, such as in children from Turkey (4/707, 0.98%) (Tamer et al. 2007). In Middle East, C. parvum is the dominant species in humans (Pirestani et al. 2008). In this study, C. parvum dominated in calves from northeastern China. It showed that precaution for Cryptosporidium spp. infection had important public health significance, which can guard against cryptosporidiosis transmission through fecal-oral route in humans. All C. parvum-positive specimens were successfully classified as IIdA19G1 subtype, based on gp60 gene. Up to now, five IId subtypes of C. parvum (IIdA14G1, IIdA15G1, IIdA17G1, IIdA19G1, and IIdA20G1) have been reported (Zhang et al. 2013). Therefore, three subtypes (IIdA15G1, IIdA19G1, and IIdA20G1) in the previous survey were identified in C. parvum from northeastern China (Tao et al. 2018, Zhong et al. 2018).
In our study, IIdA19G1 subtype has a wide range of host and geographical distributions. This subtype was described in equine animals, ruminants, rodents (Alves et al. 2006), and humans in Portugal, Sweden (Insulander et al. 2013), and Hebei, Henan, Heilongjiang, and Shanghai of China (Wang et al. 2011, Cai et al. 2017, Hu et al. 2017). The virulence variations of Cryptosporidium species and subtypes have remarkable correlation with human clinical manifestation of cryptosporidiosis. For example, IIdA15G1 and IIdA19G1 subtypes resulted in the death of newborn calves or neonatal calves due to severe watery diarrhea in a large dairy farm in Jiangsu and Ningxia provinces (Cui et al. 2014, Hu et al. 2017). Moreover, in Australia, the first report of Cryptosporidium species (IIdA19G1 subtype. 4/50, 8%) in wild buffalo was reported (Zahedi et al. 2016). A total of 88 positive diarrhea calves (IIdA19G1 subtype, 59.5%) in our study suggested that it may cause huge economic losses to the breeding industry. Meanwhile, human beings may transmit cryptosporidiosis through the fecal–oral route in this area. So it is very important to establish more sensitive molecular diagnoses to control cryptosporidiosis. And more attention should be directed toward preventing the dissemination.
In this study, microscopy detection showed the lowest sensitivity compared with RT-qPCR assay and nested-PCR. In previous study, poor sensitivity of microscopy on Cryptosporidium has been proved (Elsafi et al. 2013). Sampling time of the oocyst shedding periods and low number of eggs in samples could result in poor prevalence under microscopy (Elsafi et al. 2013). Therefore, morphology observation cannot be considered a good test method. In addition, observing Cryptosporidium spp. is difficulty on modified Kinyon's acid-fast stained smear, because of variable staining of the oocysts and small size (4 to 6 μm) (Newman et al. 1993). Moreover, microscopic tests consume huge labor cost, and also need to train microscopists with staining and examining smears (Mank et al. 1997). Prevalence difference between RT-qPCR assay (34.82%) and nested PCR (23.29%) demonstrated higher sensitivity of RT-qPCR assay than the nested PCR (p < 0.01). Not merely because RT-qPCR may detect the target molecule with low parasitemia in sample, nested PCR easily denotes false negative when parasite content is low (Bustin 2002).
In conclusion, this research mainly established a new RT-qPCR assay with high sensitivity, stability, and reproducibility, based on SYBR Green for Cryptosporidium spp. detection. RT-qPCR was found to be the most sensitive technique for the detection of Cryptosporidium app. in this study through comparing the RT-qPCR assay, nested PCR, and microscopic examination. Meanwhile, the epidemic characteristics of Cryptosporidium spp. in calves from northeastern China were obtained for the first time. The significant difference in association in symptom, age, and gender from positive Cryptosporidium calves was proved. It also revealed that the outbreak was possibly caused by C. parvum subtype IIdA19G1 in diarrhea calves from northeastern China. The findings of this study develop the epidemiological knowledge for prevention and control for calves in northeastern China.
Footnotes
Authors' Contributions
Y.C.W. was involved in conceptualization, methodology, data curations, parasite description, writing, and editing of the article; B.H.Z. carried out conceptualizations, methodology, data curation, and parasite description; J.P.L. worked on data curation, resources, investigation, and formal analysis; N.Z. and S.L.Y. were involved in conceptualization, validation, investigation, resources, data curation, and visualization; S.T.L. carried out methodology, software, formal analysis, and writing of the article; Y.Y.Z. performed molecular analysis, phylogenetic analysis, and lineage data; J.L. carried out molecular work, microscopic examination, and parasite images; N.M. was involved in molecular work, data curation, resources, investigation, and visualization.
Ethics Statement
The collection of fecal specimens from calves was agreed with farm manager, and the research protocol was reviewed and approved by the ethics committee of the Jilin Agricultural University and in accordance with the Animal Ethics Procedures and Guidelines of the People's Republic of China.
Acknowledgments
We warmly thank all the participants of interest.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was financially supported by the National Key R&D Program of China (Grant No. 2018YFD0501605).