
Abstract
The plague bacterium, Yersinia pestis, is a generalist pathogen of flea (Siphonaptera) vectors and mammalian hosts. In colonies of prairie dogs (PDs, Cynomys spp.), Y. pestis causes occasional epizootics, killing ≥90% of PDs within weeks to several months. We evaluated the effectiveness of deltamethrin, a pyrethroid insecticide, as a tool for preventing plague epizootics among three PD species. Specifically, we studied PD population growth on paired plots treated with deltamethrin for flea control or left untreated as baselines. We also evaluated PD population growth relative to flea abundance and PD density. All epizootics occurred on nontreated plots. Epizootics occurred on plots with very low PD densities as well as high densities. Mean population change, assessed by comparing visual counts of PDs in years before and during epizootics, was +88% for treated plots and −97% for nontreated plots. For comparison, an experimental oral vaccine against plague had an average change in population index or estimate during epizootics of −69% on vaccine plots compared with −83% for associated nontreated (placebo) plots. In our study and on plots not treated with deltamethrin, PD population growth was negatively correlated with flea abundance in the year before the epizootic, lending support to the hypothesis that flea abundance plays a critical role in plague transmission under natural conditions. Generally speaking, deltamethrin is a highly effective tool for plague management on PD colonies. That said, continued study is needed to refine deltamethrin treatments and to develop a more integrated strategy for plague management.
Introduction
Plague is a lethal disease that has impacted human societies for millennia and became more globally distributed during the current plague pandemic when it was introduced to multiple islands and continents, including North America (Keim and Wagner 2009). The bacterium responsible for plague, Yersinia pestis, is a generalist pathogen capable of infecting a wide range of mammalian species (Gage and Kosoy 2005). That attribute increases plague's ability to cause ecological disruptions (Biggins and Kosoy 2001) and the threat it poses to human health (Kwit et al. 2015). The varied mammalian hosts are accompanied by a large array of flea species (Siphonaptera), the primary plague vectors (Eisen et al. 2009). Vector control has proven effective in reducing plague transmission for both conservation and human health applications (Stewart and Mackie 1938, Biggins et al. 2010, Matchett et al. 2010, Eisen et al. 2020).
Conservation applications of plague management include protecting federally listed North American species such as Utah prairie dogs (PDs) (Cynomys parvidens) and black-footed ferrets (Mustela nigripes) (Biggins et al. 2010, Matchett et al. 2010). Pyrethroids have been popular tools for vector control in these and other contexts (e.g., human health; Miarinjara and Boyer 2016). Although permethrin and deltamethrin seem to have halted progression of epizootic plague in PDs, the absence of nontreated sites (“controls”) for comparison weakens that interpretation in some studies (Seery et al. 2003, Hoogland et al. 2004). Detecting the presence of plague on a PD colony does not necessarily predict imminent population collapse due to an epizootic (Hanson et al. 2006, Biggins et al. 2010, Rocke et al. 2017); so, lack of collapse following such plague detection and subsequent intervention is not incontrovertible evidence of treatment efficacy. Treatments to prevent epizootics can best be interpreted as effective if treated sites persist while paired nontreated sites nearby are decimated by plague.
One controlled study (Biggins et al. 2010) was designed to evaluate the possible existence and persistence of enzootic plague in populations of several species of PDs. In that study, the term enzootic was the binomial alternative to epizootic using a cutoff of 90% mortality within 1 year, although all three attributes of an articulate definition (mortality rate, area of coverage, and time interval) are continuums that are commonly categorized by imposing rather arbitrary cut points (Biggins and Eads 2019). The study by Biggins et al. (2010) employed a paired-plot design on three species of PDs in Montana and Utah. All burrow openings on one member of each pair were treated with deltamethrin for flea control and its counterpart was left untreated. Apparent annual survival was the measure of comparison. Because that study focused on enzootic plague, multiple sites that underwent epizootics on deltamethrin-treated or nontreated plots were excluded from analyses. We here examine those excluded data to evaluate the effectiveness of deltamethrin in preventing the occurrence of epizootics but used visual counts as the measure of comparison instead of survival.
We were concerned primarily with the influence of plague on PDs in our experimental plots. Plague causes death, but changes in populations can also be due to emigration and immigration; movements of PDs between plots of an experimental pair could contribute to differences in population counts between treated and nontreated plots. There are two seemingly contradictory hypotheses to explain why PDs might move. First, there may be movement from areas of depauperate PD densities to those of high densities, for example, if PDs prefer higher densities that might enhance survivorship and reproduction (Jones et al. 2012, Hoogland 2013). Second, PDs could be attracted from areas of high to low population densities if forage is more abundant on the latter. We examined interplot movements of PDs to evaluate the potential for disproportionate influences of emigration on treatment-specific changes in populations.
The excluded data of Biggins et al. (2010) also provided an opportunity to assess the predictive value of prior conditions for the occurrence of plague epizootics. Two pre-existing conditions that seem worthy of evaluation are vector densities and host densities (May and Anderson 1979). Lorange et al. (2005) provided a compelling argument that both the evolution of Y. pestis lethality to hosts and a need for relatively high flea abundance to maintain and enhance transmission are explained by low transmission efficiency by fleas. Although increases in survival of free-ranging hosts that result from flea control (Biggins et al. 2010, Matchett et al. 2010, Tripp et al. 2017) provide field support for their hypotheses, such extreme experiments do more to underscore the importance of fleas to plague transmission than to quantify the relationship between flea abundance and probability of epizootic development. We asked the question, during periods of epizootic plague, are PD population growth rates correlated with flea abundance or host density in the prior year?
Materials and Methods
We conducted this study on black-tailed PDs (BTPD, Cynomys ludovicianus) in north-central Montana, white-tailed PDs (WTPD, Cynomys leucurus) in eastern Utah, and Utah PDs (UPD) in southwestern Utah. We evaluated five site-year combinations that met our definition of an epizootic on at least one plot of a pair and were thus excluded from the former study (Biggins et al. 2010). Epizootics developed in 2004 in WTPDs and in 2005 in UPDs and BTPDs. We also included a sixth site-year of Montana BTPD data that were excluded from the former study because sampling began in 2004 and ended in 2005 with an epizootic on the nontreated plot. Recreational shooting of PDs was minimal to nonexistent, and poisoning of PDs was nonexistent on all pairs considered herein (Biggins et al. 2010).
We classified epizootics as ≥90% population declines during the sampling time frame of 1 year (Biggins et al. 2010). Proving that plague was the cause of such declines, however, is problematic. Plague is seldom (if ever) diagnosed in all animals that die. Various forms of evidence can suggest plague as the cause. Detections of plague preceding or during a die-off are helpful; in our case, four of the six pairs had plague detections via PD serology, fleas, or carcasses on or near the colony (table 1 and figure 2 of Biggins et al. 2010). Characteristics of the die-off itself are suggestive; precipitous collapses of PD populations are not presently known to be caused by any other disease (Barnes 1993, Cherry et al. 2019).
Visual Counts of Three Species of Prairie Dogs on Paired Dusted and Non-Dusted Plots in a Year When Plague Epizootics (Population Declines of ≥90% on at Least One Paired Plot) Were Prevalent and in the Prior Year
PD, prairie dog; BTPD, black-tailed prairie dog; WTPD, white-tailed prairie dog; UPD, Utah prairie dog.
We established experimental plots as pairs that exhibited similar ecological attributes (see Biggins et al. 2010 for details). Pairs were either on adjacent portions of the same colony or on two proximate colonies with similar characteristics. Each year, we dusted each burrow opening on the same, randomly selected member of each pair of plots with a targeted rate of 4 g DeltaDust® (0.05% deltamethrin; Bayer Environmental Science, NC), a powdered formulation of deltamethrin that is waterproof and provides effective flea control (Biggins et al. 2010, Eads and Biggins 2019).
We used visual counts as an index to PD populations on plots (Fagerstone and Biggins 1986, Menkens et al. 1990, Biggins et al. 2006). Visual counts of PDs, used as the response variables to evaluate population changes, are biased indices, but they can have sufficient precision to document changes and make comparisons (Skalski et al. 2010). Our principal assumptions were equal detectability of PDs on dusted and non-dusted plots of a pair and equal detectability between years (within plots). Visual count conditions (and ecological attributes in general) were similar within pairs within and between years.
We used these counts, converted to density (counts per ha), to evaluate the effectiveness of deltamethrin to prevent plague epizootics and assess relationships with prior year host density and flea abundance at the plot level. For each plot, we counted PDs using binoculars and/or spotting scopes from a single location outside the plot that gave the best view of the entire plot and repeated these counts on three (usually consecutive) days. We began counts just after sunrise and continued to conduct repeated systematic scans of the plot until the counts declined to about half the peak number (usually by late morning as PDs went below ground for their typical mid-day break; Hoogland 1995). We conducted visual counts on the same days for dusted and non-dusted plots of each pair, and counts were made from the same locations each year. The estimate we used to calculate density was the highest count obtained during any single scan of the plot for the 3 days (Fagerstone and Biggins 1986).
To compare changes in visual counts between dusted and non-dusted plots, we subtracted the count density estimate (count divided by area of plot; Fagerstone and Biggins 1986) of the prior year (t1) from the density estimate during the epizootic year (t2) for each plot and assessed the series of paired treatment-control differences using a nonparametric Wilcoxon signed-rank test (SYSTAT 12.0). Substantial declines were assured on at least one plot of each pair by definition (in this case, epizootics must involve ≥90% declines in population). Thus, the primary interest was the comparison of the plots with epizootics and their companion plots.
For assessing prior year effect of PD density and flea abundance, we wished to broaden the data set to include pairs of plots that did not meet the strict criteria of epizootic involvement but may have been substantially affected by increased circulation of plague during the years when epizootics became common. Thus, we included all PD density and flea abundance values from non-dusted plots (n = 14) sampled during the epizootic year (2004 for WTPD and 2005 for other species) and the corresponding year prior. We used visual counts of PDs for each plot in the prior year divided by area of the plot. For statistical analysis, we transformed the change in visual count density into a simple binomial; PD density on a plot either increased or decreased over the 1-year interval.
To assess an effect of flea abundance during the year before the epizootic, we combed the captured PDs and counted the number of fleas on each PD, as described in the study by Biggins et al. (2010). Capture and handling procedures are detailed in the study by Biggins et al. (2010). We wished to exclude the effect of deltamethrin dusting from flea evaluation, so analyses included data from non-dusted plots only. Our assessments of flea and PD densities were done at the scale of plots (colonies or portions thereof). We summarized flea data as a mean flea count per PD for each plot. We used the Mann–Whitney tests in two separate analyses to examine relationships between flea and PD density and population change.
Spatial data from trapping and marking were used to examine movements between plots. We used PD capture locations from 2000 to 2005 for the three species of PDs (Biggins et al. 2010), tallying all moves between plots of a pair by individually marked PDs. We examined all recaptures of PDs and categorized the total numbers of intertreatment movements between paired plots as either toward or away from the dusted plot. We used a chi-squared test to compare the proportions of movements in the two directional categories.
Our capture and handling of PDs followed guidelines of the American Society of Mammalogists (Sikes et al. 2011). Trapping and PD handling were authorized under a U.S. Fish and Wildlife Service endangered species permit (for UPD) and was permitted under state wildlife collection permits from Utah Division of Wildlife Resources and Montana Department of Fish, Wildlife, and Parks.
Results
All epizootics occurred on the non-dusted plots of pairs. Despite the small sample size of site pairs and attendant low power, the effect of dust treatment was significant (Z = 2.021, p = 0.028). The mean change in population density was an 88% increase on dusted plots and a 97% decline on non-dusted plots with no overlap in ordinal rankings of rate of change for pairs (Table 1).
For the nontreated plots, we detected a significant negative relationship between PD population change over the annual interval and the mean number of fleas per host at the beginning of the interval (X 2 1 = 4.840, p = 0.028; Fig. 1), but we failed to detect a relationship between population change over the annual interval and density of PDs at the beginning of the interval (X 2 1 = 1.284, p = 0.257; Fig. 2). Contrary to popular perceptions of host density dependence, epizootics (as defined) occurred on colonies with very low PD count densities (1.1 PD per ha) as well as higher densities (5.5 PD per ha).
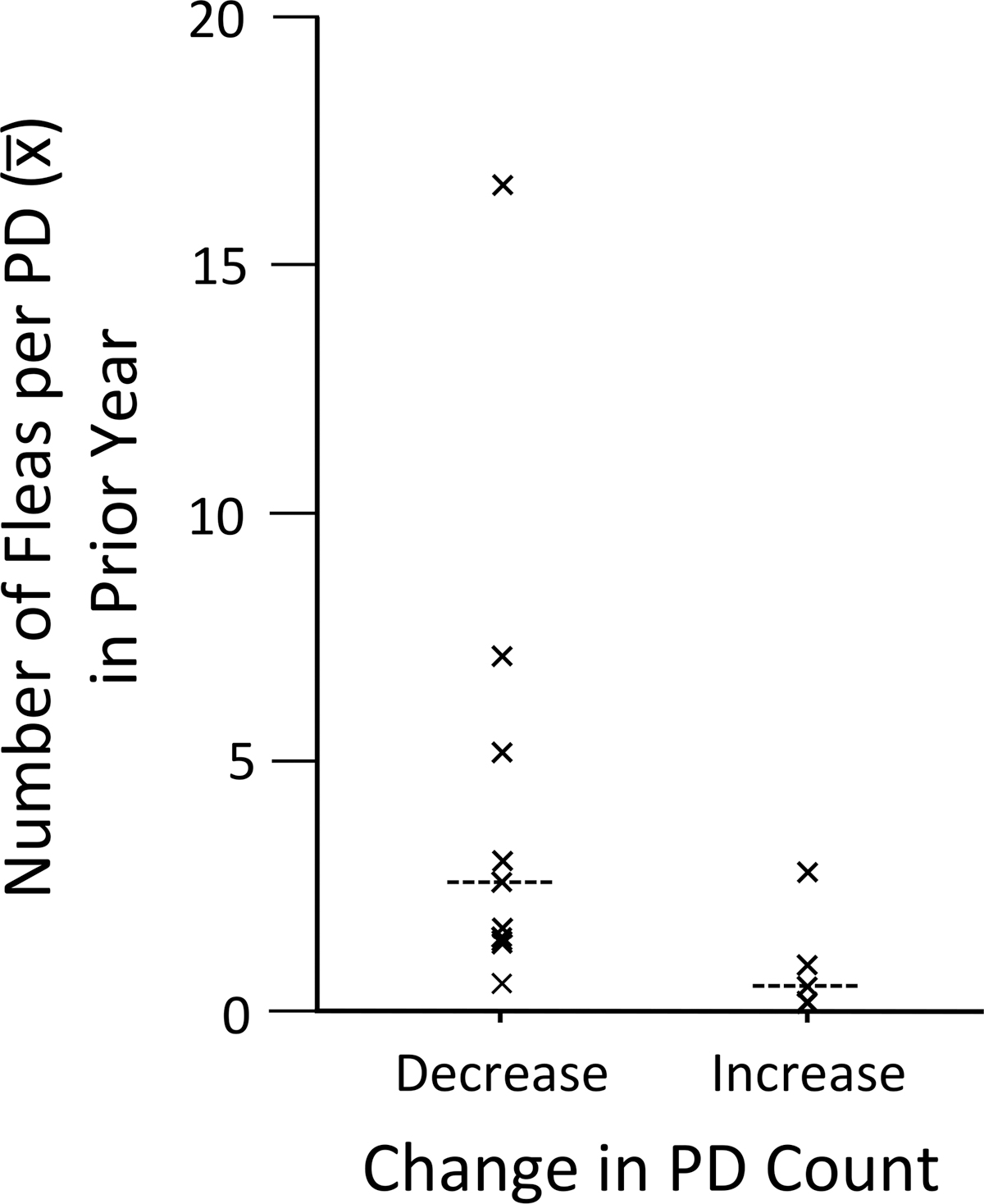
Flea abundance on prairie dogs in non-dusted plots the year before a plague epizootic year, grouped by plots that underwent decreases (n = 9) and increases (n = 5) in prairie dog visual counts during the following year (medians indicated by dashed horizontal line).
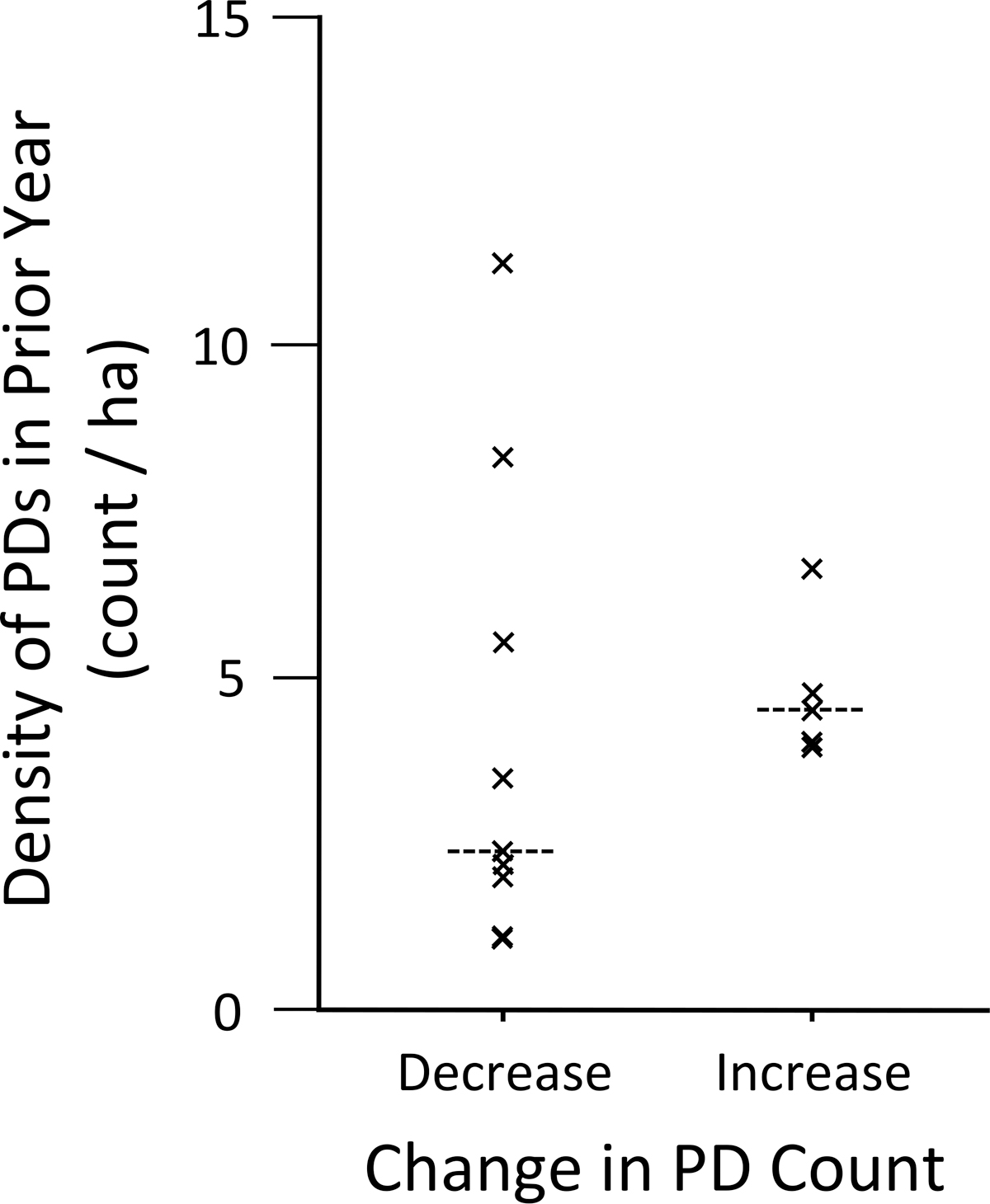
Visual count densities of prairie dogs in non-dusted plots the year before a plague epizootic year, grouped by plots that underwent decreases (n = 9) and increases (n = 5) in prairie dog visual counts during the following year (medians indicated by dashed horizontal line).
Centroids of all PD captures (average locations for X and Y axes) on dusted and non-dusted pairs of plots were separated by 256–10,590 m. Movements of PDs between paired plots were infrequent (1.54% of all 1550 recapture events). Departures from dusted plots (2.24% of 624 recaptures) tended to be more common (X 2 = 3.312, p = 0.069) than movement into dusted plots (1.08% of 926 recaptures), consistent with the idea that dispersal would be greater from plots with higher densities of PDs rather than decimated colonies. Considering just the plot pairs with epizootics, there was just a single detected move by a marked PD (a UPD that moved from the non-dusted to the dusted plot at the Tom Best colony).
Discussion
A central purpose to the present study was evaluating PD population changes on the plots that had plague vector control compared with their counterparts that had no treatment. The average reduction in visual counts was 97% for non-dusted plots of our experimental pairs (Table 1), a value that is no surprise because we had defined these members as experiencing epizootics (Biggins et al. 2010). The utility of the comparison comes from the average 88% increase in counts on the associated members of each pair that were treated with deltamethrin. The one dusted colony that underwent a 68% population decline as its counterpart collapsed remains a noteworthy exception. This occurred on the study area (Coyote Basin) where flea control was least effective, and annual survival rates of PDs were lowest under conditions of presumed enzootic plague (Biggins et al. 2010).
Variation in application rate of deltamethrin influences efficacy of flea control. Although we aimed to apply about 4 g of DeltaDust per burrow opening, there was considerable variation during this study (1.19–4.50 g/burrow; Eads and Biggins 2019). Other studies also have described occasional events when deltamethrin did not prevent substantial mortality of PDs from plague (Griebel 2011, Tripp et al. 2017, Hoogland et al. 2018). Flea populations at these example sites might have increased before the populations of PDs were resampled and/or deltamethrin was reapplied; in one case, 14 months had elapsed after the last application of deltamethrin (Griebel 2011). Perhaps, the current rates of application or the timing of application are not optimal to keep flea numbers low for the typical 12-month interval between applications (Tripp et al. 2016, Eads and Biggins 2019). Also, resistance to deltamethrin can evolve in fleas after repeated annual applications of the insecticide (Eads et al. 2018).
Did PD movements between paired experimental plots affect these results? The two hypotheses relating to density dependence and PD movements are not necessarily mutually exclusive; at the extremes of density, one tendency may be supplanted with the other. If protection from plague results in increased PD survival and population growth, patterns of movement toward or away from dusted plots could reduce or increase estimates of treatment effect. For analyses of individual PDs, apparent survival estimates can be influenced by such movements, and for metrics that do not involve marked PDs, population metrics (like our visual counts) can also be affected. In addition, for PDs that leave a dusted plot, residual dust in their fur likely does not dissipate immediately, perhaps providing them with residual protection against fleas and plague after arrival on non-dusted plots. In our study, neither influence is expected to be substantial judging from the low rates of PD exchange, even when plots were immediately adjacent to each other on split colonies (Table 1). Our study provided evidence that PDs show a high degree of site fidelity and rarely move between treatment units. Indeed, PD families are considered as groups that defend a common territory (Hoogland 1995), and such territoriality may anchor most PDs in place (assuming colony structure remains intact; Hoogland, 2013, Biggins and Eads 2019).
Our data partially support the contention of Lorange et al. (2005, p. 1911) that “… plague epizootics depend on a coincidence of high vector and host densities and that flea control is a sufficient preventative measure …” Responses in this study and others (Biggins et al. 2010, Matchett et al. 2010) to drastic reductions in flea abundance as accomplished by vector control underscore the importance of flea abundance to plague transmission. However, observations at the extremes of the continuum incompletely address the proposed (Lorange et al. 2005) effects of flea abundance on probability of transmission and maintenance of enzootic plague in the moderate portion of the range in density. Our study suggests that flea abundance in the prior year on plots without vector control was negatively associated with PD population growth rates in the subsequent year when epizootic plague became common. That result is consistent with other data showing that survival of individual UPDs was negatively correlated with flea parasitism in the previous year (Eads et al. 2020), data that were again generated at sites where plague had been detected during the study (Rocke et al. 2017). In an effort to separate cause from consequence, we used flea data collected the year before there was evidence of a plague epizootic. Increases in flea detections during an epizootic may be a consequence of the epizootic because rapidly declining host numbers likely cause fleas to aggregate on survivors (Tripp et al. 2009). During an epizootic, host deaths may reinforce a positive feedback loop that eliminates the distinction between cause and effect and dramatically increases transmission rates for Y. pestis (Biggins and Eads 2019).
In contrast to the results of our study, Brinkerhoff et al. (2010) reported flea abundance had little value in predicting epizootics in PDs. The authors mentioned that landscape context may be more important than other factors. That study was in a suburban area of Boulder County, Colorado, which may have had low colony connectivity due to intervening suburban and urban development. If a proximate source of Y. pestis is a critical factor in determining when and where epizootic outbreaks will occur (Collinge et al. 2005), persistent enzootic plague in more natural landscapes may allow other factors to play roles that might be overwhelmed by simple presence or absence of Y. pestis in suburban areas.
A link between host densities and epizootic outbreaks was not supported in this study. Similarly, Brinkerhoff et al. (2010) failed to detect an influence of PD density on epizootic development. Other factors that might trigger epizootics (e.g., factors increasing host mortality and flea questing, such as shooting and poisoning of PDs) may initiate positive feedback cycles in flea:host ratios and attendant infection rates that reduce the influence of initial host density on epizootic eruption (Biggins and Eads 2019).
Epizootic plague was detected in 2004–2005 over a broad geographic area of the western United States (Wimsatt and Biggins 2009). Besides the Montana and Utah outbreaks described herein, there was an apparent expansion of epizootic plague into South Dakota. A widespread outbreak was first noticed on the Pine Ridge Reservation in 2005, which by 2008 had expanded into an important black-footed ferret reintroduction area in Badlands National Park and the adjacent Buffalo Gap National Grasslands of Conata Basin (Griebel 2011). The latter scenario evolved into a treatment-control experiment because available resources limited plague management with deltamethrin to less than half of the PD colonies serving as black-footed ferret habitat. By 2011, populations of PDs on most nontreated colonies had collapsed, whereas the dusted colonies persisted and sustained ferrets (Griebel 2011).
Other controlled studies that evaluated plague management included additional trials of deltamethrin (Tripp et al. 2017) and a sylvatic plague vaccine (SPV; Rocke et al. 2017, Tripp et al. 2017, Boulerice 2017, David Eads and Dean Biggins, USGS, unpublished data, 2013–2016). Table 2 summarizes cases involving epizootic plague during those studies, using a relaxed loss criterion for defining an epizootic of ≥80% instead of >90%. Site pairs were included if either plot of a pair suffered a population reduction of ≥80% during the interval considered. Plague control seemed less than absolute with either vector control or SPV. There were cases involving substantial losses of PDs on dusted plots in both our study and that of Tripp et al. (2017). However, the average change in population index or estimate during epizootics was −67% on SPV plots, compared with −86% for associated nontreated (placebo) plots. By comparison, our dusting study data pooled with those of Tripp et al. (2017) show an average change of +56% for dusted plots and −97% for paired associates that were not treated. Pooling seemed warranted; reducing the criterion to ≥80% loss added no additional sites to our study because there were no cases with loss rates in the interval 80–90%.
Prairie Dog Population Attributes in Years Before Plague Epizootics and Years When Epizootics Were Detected (Defined Here As Population Declines of ≥80% on at Least One Paired Plot) on Plots Treated with Deltamethrin Dust, Sylvatic Plague Vaccine Baits, Placebo Baits, or Left Untreated (No Dust)
Abundance = mark–recapture abundance estimate; Density = mark–recapture prairie dog abundance divided by hectare.
MNA, minimum number alive; CPUE, catch per unit effort; SPV, sylvatic plague vaccine; GPD, Gunnison's prairie dog.
The apparent protection afforded by vector control during plague epizootics supports observations of curtailed epizootics after treatments of pulicides during uncontrolled studies (Seery et al. 2003, Hoogland et al. 2004). Deltamethrin treatments also improved survival of adult PDs by >40% under suspected enzootic circulation of plague (Biggins et al. 2010). At the Montana study area, the moderate effect of vector control on PD survival (Biggins et al. 2010) seemed to be amplified at higher trophic levels; survival rates of black-footed ferrets (prey specialists on PDs) increased by >200% when they were protected from enzootic plague by either vector control or F1-V vaccination (Matchett et al. 2010).
In conclusion, results herein support the hypothesis (Lorange et al. 2005) that increased flea abundance favors increased plague transmission. Deltamethrin is highly effective and is currently the most useful tool for plague management on PD colonies. Limitations of any tool, including deltamethrin, require careful consideration. Research is needed to investigate factors that influence the efficacy of deltamethrin, and additional tools are needed for integrated plague management (Tripp et al. 2017, Eads et al. 2018, 2019, Eads and Biggins 2019).
Footnotes
Acknowledgments
We appreciate assistance from L. Carter at the Centers for Disease Control and Prevention (CDC) for testing of flea pools, blood samples, and carcasses. We remain especially grateful to other colleagues at CDC (K. Gage, J. Montenieri, M. Kosoy, R. Eisen, Y. Bai) for their friendship and tutelage on plague over many years. We are deeply indebted to our many crew leaders and technicians who assisted them (n = 70). R. Reading and B. Miller of the Denver Zoological Foundation arranged logistical support for parts of the study. Additional in-kind support was provided by the Bryce Canyon National Park, Dixie National Forest and Bureau of Land Management offices in Utah (Vernal, Cedar City, Richfield, and Torrey), Colorado (Meeker), and Montana (Malta). We thank J. Hoogland, J. Hughes, and two anonymous reviewers for constructive comments on the article. Any use of trade, product, or firm names is for descriptive purposes only and does not imply endorsement by the U.S. Government. Data are available from Biggins and Eads (![]() ).
).
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
Funding was provided by several federal agencies, including the Fish and Wildlife Service, the Geological Survey, and the Bureau of Land Management, supplemented by additional funds from the Utah Division of Wildlife Resources and the Utah Department of Natural Resources Endangered Species Mitigation Fund.