
Abstract
In this study, we isolated a group of viruses that can cause a cytopathic effect in BHK-21 cells from sandfly specimens collected in Yangquan County, Shanxi Province, central China. The representative virus strain is SXYQ1860 and, with increased viral injection, the suckling mice became ill and died, with viral titers continually increasing. The results of molecular genetic evolution analysis of the three gene segments of the virus, L, M, and S, indicate that the newly isolated viruses from Yangquan County are the same as the Wuxiang virus (WUXV) previously isolated from sandflies collected in Wuxiang county, China. This is the first time that the WUXV was also isolated outside Wuxiang County. Therefore, strengthening the surveillance of neglected sandflies and the viruses they transmit to help prevent and control arboviruses and the associated diseases is essential for public health.
Introduction
Sandflies belong to the family Phlebotominae within the Psychodidae and Diptera and are distributed worldwide. Sandflies have a blood-sucking habit and are fond of human blood in addition to a variety of animal sources, including dog, chicken, and sheep (Killick-Kendrick 1999). Sandflies can spread various viruses through blood sucking (Depaquit et al. 2010). Due to the broad geographic distribution of sandflies and the variety of pathogens that they can transmit, sandflies are important public health harmful insects. They can transmit diseases that cause fever, bleeding, and even encephalitis, thereby threatening public health and imposing a burden of disease (Depaquit et al. 2010, Ayhan et al. 2017).
Sandfly-borne phleboviruses are distributed worldwide and its infection in humans can cause fever, and even severe clinical symptoms such as meningitis and encephalitis, making sandfly-borne phleboviruses a significant public health issue (Tesh et al. 1976, Charrel et al. 2005, Izri et al. 2008). According to the report of the International Committee on Taxonomy of Viruses (ICTV), the genus Phlebovirus contains numerous viruses, including Rift Valley fever virus and sandfly fever Naples virus (Kuhn et al. 2020). In recent years, new viral species with genomic characteristics of phleboviruses have been isolated from sandfly specimens collected around the world (Elliott and Brennan 2014).
Our research group previously reported that a virus was isolated from sandfly specimens collected in Wuxiang County, China. Molecular genetic analysis of the virus showed that it was a new member of the genus Phlebovirus, and it was named Wuxiang virus (WUXV). This is the first time that a phlebovirus has been isolated from sandfly specimens collected from the wild in China (Wang et al. 2020). In the present study, we report that WUXV was isolated again from wild sandfly specimens collected in Yangquan, Shanxi Province, China (Fig. 1). This is the first time that WUXV has been isolated from outside Wuxiang County, suggesting that WUXV has a relatively broad geographic distribution.
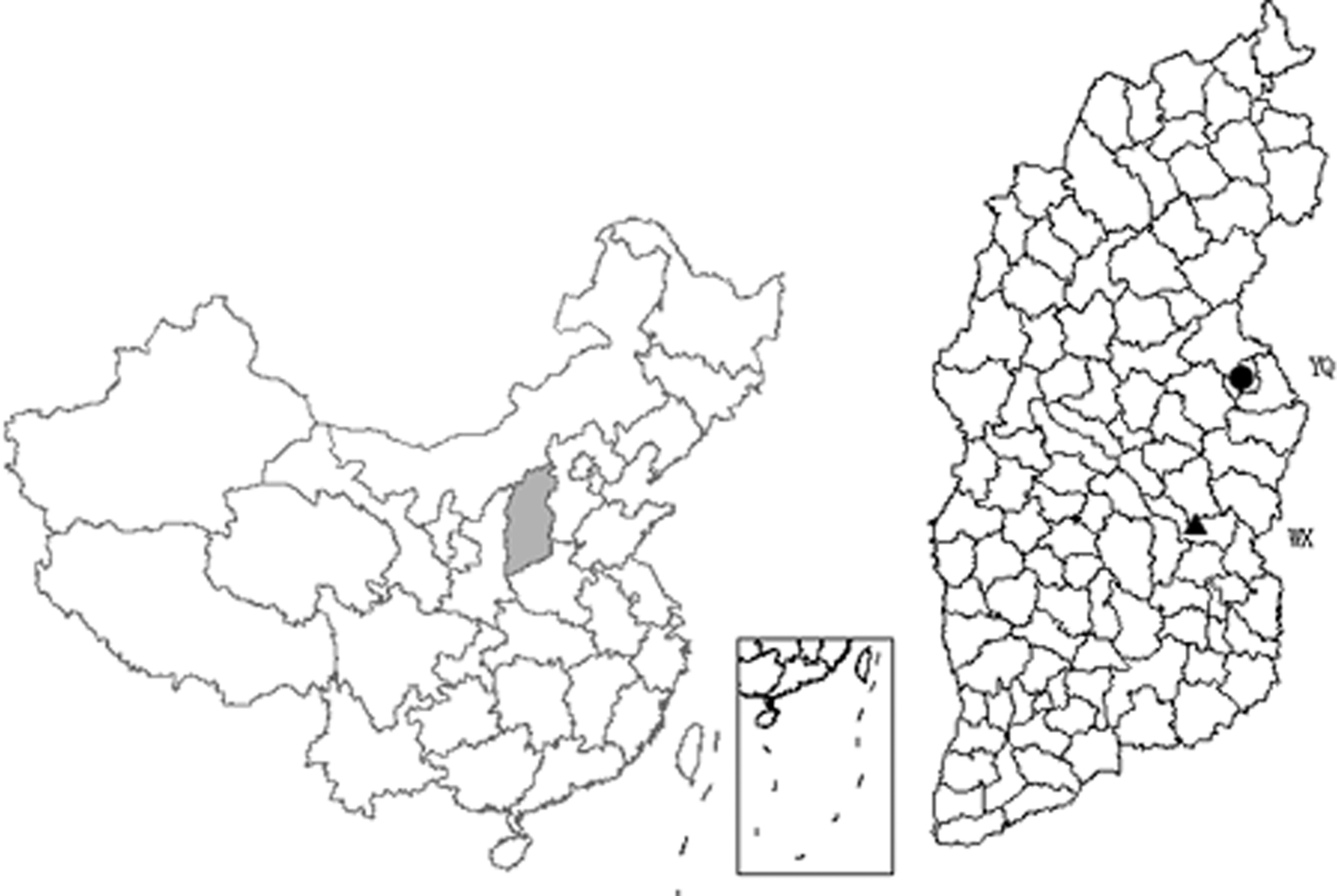
Collection of blood-sucking insect specimens in Yangquan County, Shanxi Province, 2018. The shaded area in the figure is Shanxi Province in central China. Yangquan County (YQ), where specimens were collected for this study, is represented by a circle, and the triangle represents Wuxiang County (WX), where WUXV was first isolated in China. The distance between the two sites is about 200 km. WUXV, Wuxiang virus.
Materials and Methods
Collection of blood-sucking insect specimens
The specimen collection tools used for this study are Wen Taitai, a collection tool of sucking insects (Battery 12 V 15 Ah, bulb 0.1 A, fan 0.14 A, MM200BL;Guangzhou Changsheng Chemical Technology Service Co., Ltd.,) and a carbon dioxide mosquito trap lamp (MT-1; Beijing Detelong Technology Development Co., Ltd.). The collection tool was hung in a livestock pen from 18:00 pm to 7:00 am the next morning to collect specimens. All specimens were morphologically identified, packaged, and numbered under ice bath conditions. Specimens were stored in liquid nitrogen until laboratory testing (Huemer et al. 2017, Song et al. 2017, Wang et al. 2020). To identify the sandfly species of the positive pools, PCR was used to amplify the cytochrome c oxidase I (COI) gene (Wang et al. 2020).
Cell culture and virus isolation
BHK-21 cells (Golden hamster kidney cells) and C6/36 cells (Aedes albopictus egg cells) were stored in the laboratory. BHK-21 cell culture: 90% Eagle's medium (laboratory preparation), 7% fetal bovine serum (FBS; Invitrogen), 1% penicillin and streptomycin (100 U/mL), 1% glutamine (30 g/L), and 1% NaHCO3; and C6/36 cell culture: 89% RMPI 1640 (Invitrogen), 10% FBS (Invitrogen), and 1% penicillin and streptomycin (100 U/mL). BHK-21 and C6/36 cells were cultured in an incubator containing 5% CO2 at 37°C and 28°C, respectively (Song et al. 2017, Wang et al. 2020).
The sandflies were pooled in groups of 50–100, and mosquitoes were pooled in groups of 50. After washing, abrasive fluid was added and specimens were ground repeatedly in an ice bath with 1.5 mL of grinding fluid. After grinding and centrifugation (4°C, 12, 000 rpm, 30 min), 100 μL of the supernatant was removed to inoculate BHK-21 cells or C6/36 cells in 24-well plates (Corning, Incorporated), then the cells were continuously cultured to observe the cytopathic effect (CPE) (Huemer et al. 2017, Song et al. 2017, Wang et al. 2020). SXWX1813-2 isolate had been isolated previously in the Wuxiang county in 2018 (Wang et al. 2020).
Viral plaque assay and viral titer determination
Viral plaque assay
BHK-21 cells were introduced into a six-well culture plate (Corning, Incorporated) and grown into a monolayer with coverage of 80%. A series of 10-fold viral dilutions (10−1–10−6) was successively added to the six-well culture plate (0.1 mL/well). After incubation for 1 h at 37°C under 5% CO2, 1% agarose-MEM (mineral essential medium) containing 2% FBS was added to cover the cells (3 mL/well). After culturing the cells for 3 days, we added a second layer of 1% agarose-MEM containing 7% neutral red and 2% FBS to cover the cells (3 mL/well). The appearance time and number of plaques were observed and the diameter of plaques was measured. The data are statistically processed by SPSS software, and independent sample t test is used (Song et al. 2017, Wang et al. 2020).
Viral titer
A series of 10-fold dilutions of the virus were added to the 6-well culture plate. After incubation, 1.3% methylcellulose-MEM semisolid medium containing 2% FBS was added to each well (4 mL/well). After culturing for several days, plaques were obvious, and the cells were stained with crystal violet solution. Then, the plaque-forming units (pfu) and viral titers were calculated (Song et al. 2017, Wang et al. 2020).
Electron microscope observation
Brain tissues of suckling mice that had died of the virus after inoculation were collected, and the samples were fixed with 2% formaldehyde and 2.5% glutaraldehyde solution. The samples were sliced on an ultramicrotome (80 nm), dried at room temperature, stained with uranium acetate and lead citrate, and then observed using a transmission electron microscope (TF20; FEI Company) (Feng et al. 2017).
Animal experiment
Suckling mice (Kunming mice) were purchased from Beijing Vital River Laboratory Animal Technology Co., Ltd. (quality code SCXK Beijing 2016–0006). All mice were maintained under sterile conditions in the animal facility of the Chinese Center for Disease Control and Prevention. All animal experiments were conducted in strict accordance with the animal experiment regulations established by the Ethics Committee of the Chinese Center for Disease Control and Prevention.
Each suckling mouse that was aged 3 days was inoculated with 20 μL of viral infection supernatant and allowed to continue feeding. Seven to ten suckling mice were inoculated in each group, and the mice were sacrificed and dissected 14 days after viral inoculation. Then brain tissue of each passage of suckling mice was ground separately.
The supernatant was obtained after centrifugation at 12,000 rpm, 30 min, without filtration. The supernatant was collected and inoculated into the next group of suckling mice. Viral isolates were passed continuously four times in suckling mice. The virus inoculated in the brain of each passage was the grinding supernatant of the brain tissue of suckling mice have already become ill and died after the last inoculation. The morbidity and mortality of suckling mice were observed and recorded every day, and the plaque assay was used for the determination of virus titer (Cao et al. 2016).
Viral RNA extraction and complementary DNA synthesis preparation
We used the Viral RNA Mini Kit (QIAamp; Qiagen, Valencia, CA), following the instructions to extract total RNA from each sample, and used the Ready-To-Go kit (GE Healthcare, Little Chalfont, Buckinghamshire, United Kingdom) to prepare a complementary DNA (cDNA) synthesis of the extracted viral RNA for subsequent viral gene amplification (Song et al. 2017, Wang et al. 2020).
Viral gene amplification and nucleotide sequence determination
The PCR system was 25 μL in volume, including cDNA template, GoTaq® Green Master Mix 2 × (Promega, Madison, WI), and 10 pmol/μL each of the upstream and downstream primers. WUXV L, M, and S gene amplification primers were used for viral gene amplification from viral isolates (Wang et al. 2020). The success of gene amplification was detected using 1% agarose gel electrophoresis before nucleotide sequence determination. The sequencing method is Sanger dideoxy sequencing.
Nucleotide sequence analysis
BLAST (NCBI) alignment of nucleotide sequences. SeqMan software (DNAStar, Madison, WI) was used for nucleotide sequence splicing and quality analysis. BioEdit (Version 7.0, Thomas) software was used for nucleotide multiple sequence alignment (Bootstrap NJ tree). MEGA6.0 software was used for phylogenetic analysis based on the neighbor-joining method with a bootstrap value of 1,000. MegAlign was used for homology analysis of nucleotide and amino acid sequences (Song et al. 2017, Feng et al. 2019, Wang et al. 2020).
Results
Blood-sucking insect collection
In June 2018, blood-sucking insects were collected from chicken pens, sheep pens, and abandoned caves in villages in Yangquan County (112°5′–114°4′ east longitude, 37°40′–38°31′ north latitude), Shanxi Province. A total of 4070 blood-sucking insect specimens were collected, including 3996 sandflies. In addition, 74 mosquito specimens were collected (including Culex [52], Armigeres [13], Aedes [8], and Anopheles [1]). In addition, the sandflies species for each of the 10 phlebovirus isolates positive pools were identified as Phlebotomus chinensis.
Virus isolation
The blood-sucking insect specimens were divided into 54 pools (51 pools of sandfly specimens and three pools of mosquito specimens). After grinding and centrifugation, the supernatant was inoculated with BHK-21 cells and C6/36 cells in parallel. Three days after inoculation of BHK-21 cells with SXYQ1860 (representative strain), CPE was apparent, showing that the cells contracted and detached (Fig. 2A). CPE could be stably serially passaged in BHK-21 cells. After 51 pools of sandfly grinding supernatant were inoculated onto BHK-21 cells, a total of 10 isolates similar to the SXYQ1860 virus strain, which could cause CPE in BHK-21 cells, were obtained. The grinding supernatant of three pools of mosquitoes did not cause CPE in BHK-21 cells. All 51 pools of sandfly specimens and three pools of mosquito specimens were also inoculated into C6/36 cells after grinding, and no CPE was observed after three consecutive passages.
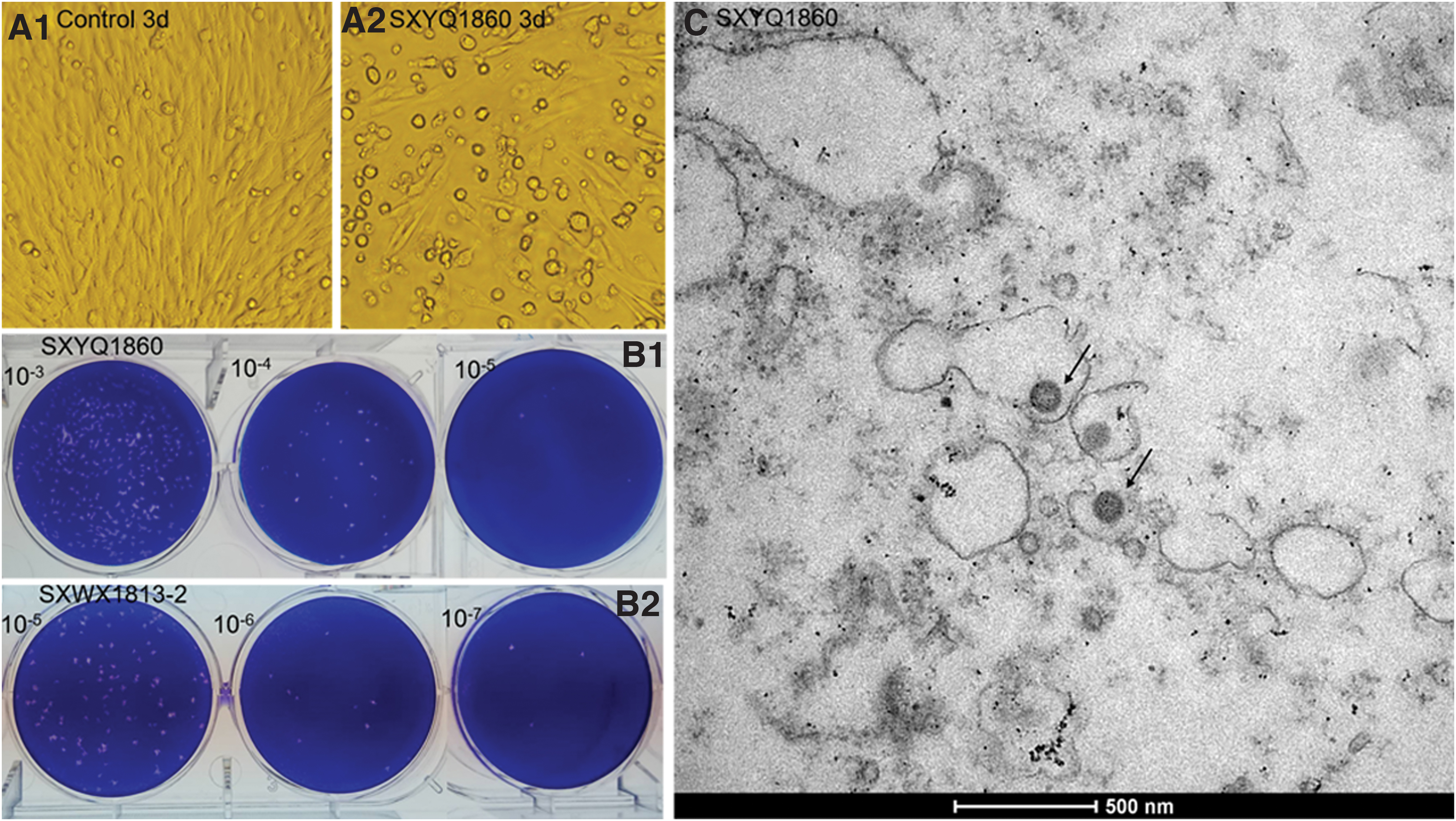
SXYQ1860 virus-caused CPE of BHK-21 cells, viral titer and electron micrograph.
In the supernatants of 10 strains of virus isolates that caused CPE in BHK-21 cells, WUXV S gene amplification primers showed a positive result (Table 1) and WUXV was identified through nucleotide sequence determination and analysis. The corresponding 10 pools of C6/36 supernatants (3 consecutive passages) did not show positive amplification of WUXV genes. The three pools of mosquito specimens did not show positive WUXV gene amplification in either BHK-21 or C6/36 cell supernatants.
Isolation and Identification of Wuxiang Virus in Yangquan County, Shanxi Province
S gene of WUXV amplification primers were used for viral gene amplification from viral isolates and 10 isolates of S gene amplification results are all positive. SXYQ1849-1 virus isolate came from sheep pens, SXYQ1875-2 virus isolate came from an abandoned cave, and the remaining 8 virus isolates came from chicken pens.
CPE, cytopathic effect; WUXV, Wuxiang virus.
SXYQ1860 virus can form plaques in BHK-21 cells (Fig. 2B1), and the average diameter of those plaques is about 2.6 ± 0.4 mm (n = 10) (Table 2). The SXYQ1860 virus was subjected to two rounds of plaque purification in BHK-21 cells to obtain a single viral plaque. The purified plaques were inoculated into BHK-21 cells to amplify the virus, then aliquoted and frozen in an ultra-low temperature refrigerator. The virus-induced CPE, electron microscopy, animal experiments, and viral genome sequencing conducted in this study used the SXYQ1860 virus after plaque purification. Spherical viral particles were visible on ultrathin sections of infected suckling mouse brain tissue, which had a diameter of 80–100 nm (Fig. 2C).
Phenotypic Characteristics of SXWX1813-2 Virus and SXYQ1860 Virus
“—” Indicates that the test result was negative.
pfu, Plaque-forming units.
Animal challenge with SXYQ1860 virus
We studied the pathogenicity of SXYQ1860 virus in suckling mice. After inoculating 20 μL (1 × 105.6 pfu/mL) of SXYQ1860 virus (BHK-21 7th passage) supernatant into the suckling mouse brain, we observed and recorded the morbidity and mortality of suckling mice. No morbidity or mortality occurred in the first or second passage of suckling mice after inoculation and incubation for 14 days. On the 10th day after the third passage (n = 10), onset of symptoms began, such as failing to consume milk, convulsion and rigor of the limbs, lateral lying position, and negative turning reflex. The suckling mice died on the 10th (n = 1), 11th (n = 1), 12th (n = 1), 13th (n = 4), and 14th days (n = 1). The remaining two suckling mice showed no morbidity or death on the 14th day after viral inoculation. All animals (n = 7) at the fourth passage became ill on the 6th day and died successively on the 7th (n = 1), 8th (n = 2), 9th (n = 2), and 10th days (n = 2) (Fig. 3).
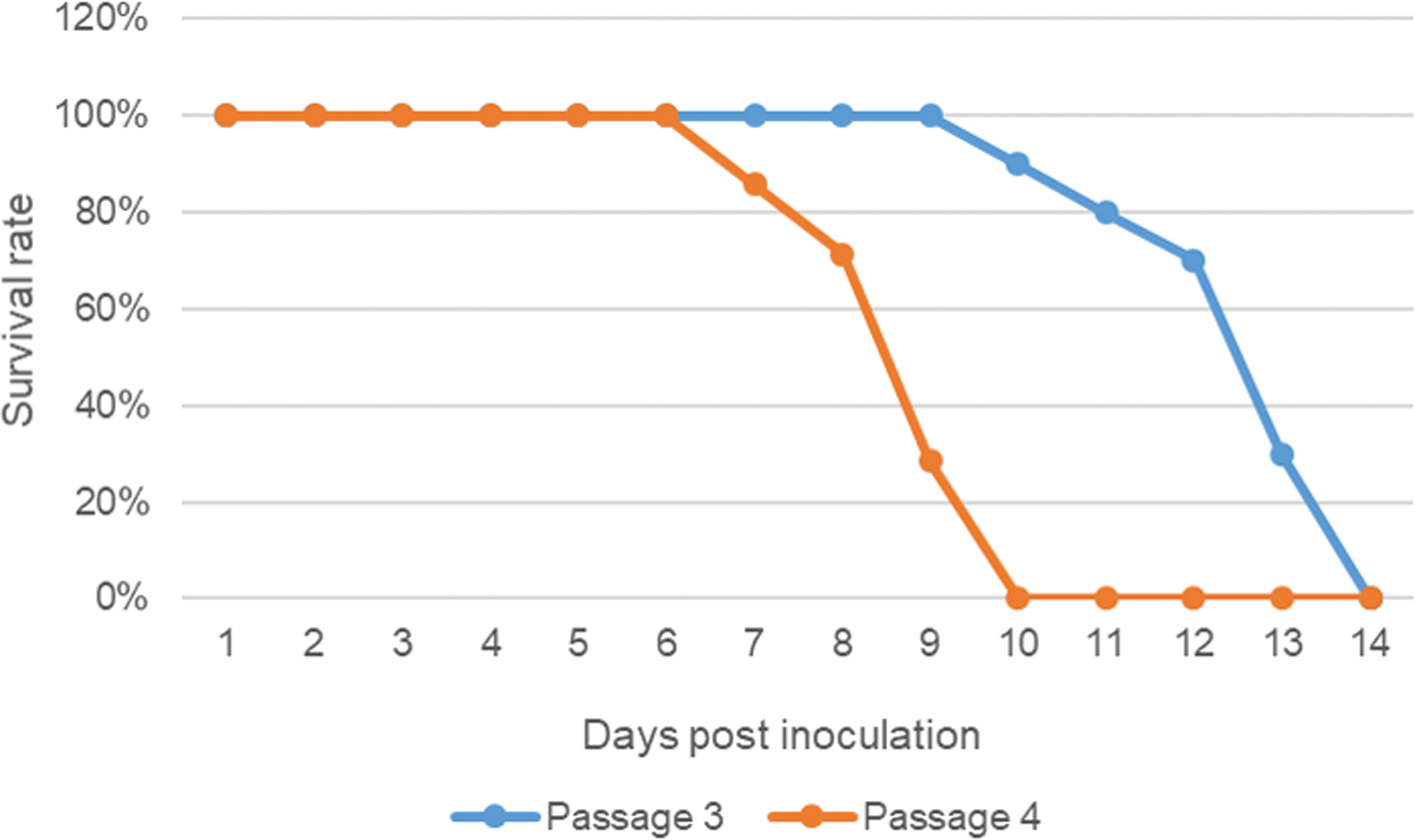
Animal challenge with SXYQ1860. The two polynomial curves shown are the survival rates of suckling mice in the third and fourth passages after inoculation with SXYQ1860, respectively. Color images are available online.
Titers of SXYQ1860 virus in the brain tissues of four consecutive passages of suckling mouse brains were detected through plaque formation experiments. The results showed that the viral titers of suckling mouse brains in the first to fourth passages were 1 × 105.86, 1 × 106.05, 1 × 106.32, and 1 × 106.60 pfu/mL (Fig. 2B1), respectively.
Characteristics of molecular biology
Use the virus cDNA prepared in this study and primers to use gene amplification methods to obtain the whole virus genome. The genome of the SXYQ1860 virus contains three segments. The nucleotide sequences of the coding regions of these three segments showed that the nucleotide and amino acid counts of the L segment (GenBank acc. no. MT767755) were 6273 nt and 2090 aa, encoding RNA-dependent RNA polymerase (RdRp); the M segment (GenBank acc. no. MT767756) contained 4089 nt and 1362 aa, encoding glycoprotein (GP); and the S segment (GenBank acc. no. MT786480) includes 1611 nt of genes and contains two open reading frames (ORFs) encoding nonstructural protein (NS; 260 aa) and nucleocapsid protein (N; 246 aa). The length of nucleotide and amino acid sequences of the S segments of the other nine isolates were the same as those of SXYQ1860.
The nucleotide and amino acid homology results for the SXYQ1860 virus isolated in Yangquan sandflies in this study and the WUXV isolated previously show that the SXYQ1860 virus has a homologous nucleotide (amino acid) sequence of the L gene with WUXV (SXWX1813-2), with similarity of 97.7% (99.3%); the nucleotide (amino acid) homology between SXYQ1860 and WUXV (17 isolates, including SXWX1813-2) M gene, NS gene, and N gene are 96.5–96.9% (97.1–97.8%), 96.6–98% (97.3–98.1%), and 98–99.2% (99.2–99.6%), respectively, and their homology with other viruses isolated from sandflies outside China is significantly lower than that with WUXV.
The viruses isolated from Wuxiang and Yangquan showed the highest levels of homology with Toros virus (TORV) and Corfou virus (CFUV), which had L segment nucleotide (amino acid) homology of 76.5–77.0% (88–88.2%), M segment nucleotide (amino acid) homology of 71.2–71.9% (75.0–75.7%), S segment NS gene nucleotide (amino acid) homology of 74.3–75.4% (84.3–85.1%), and S segment N gene nucleotide (amino acid) homology of 81.9–82.2% (96–96.4%) (Table 3). These results indicate that the viruses isolated from Yangquan sandflies can be attributed to WUXV, representing a newly isolated strain of this virus.
Genome Sequence and Homology of SXYQ1860
“/” Indicates that no sequences were available in the GenBank database.
CFUV, Corfou virus; GP, glycoprotein; N, nucleocapsid protein; NS, nonstructural protein; RdRp, RNA polymerase; TORV, Toros virus.
The results of molecular genetic evolution analysis (Supplementary Table S1) of the nucleotide sequence of the viral genome show that regardless of the L, M, and S (N) genes, Drin virus (Bino et al. 2019), CFUV, TORV, and WUXV, including the 10 isolates isolated in Yangquan in the present study, belong to the mosquito- and sandfly-borne virus group, which shows the closest relationships with TORV, CFUV, and Drin virus (subbranch II) (Fig. 4A–C).
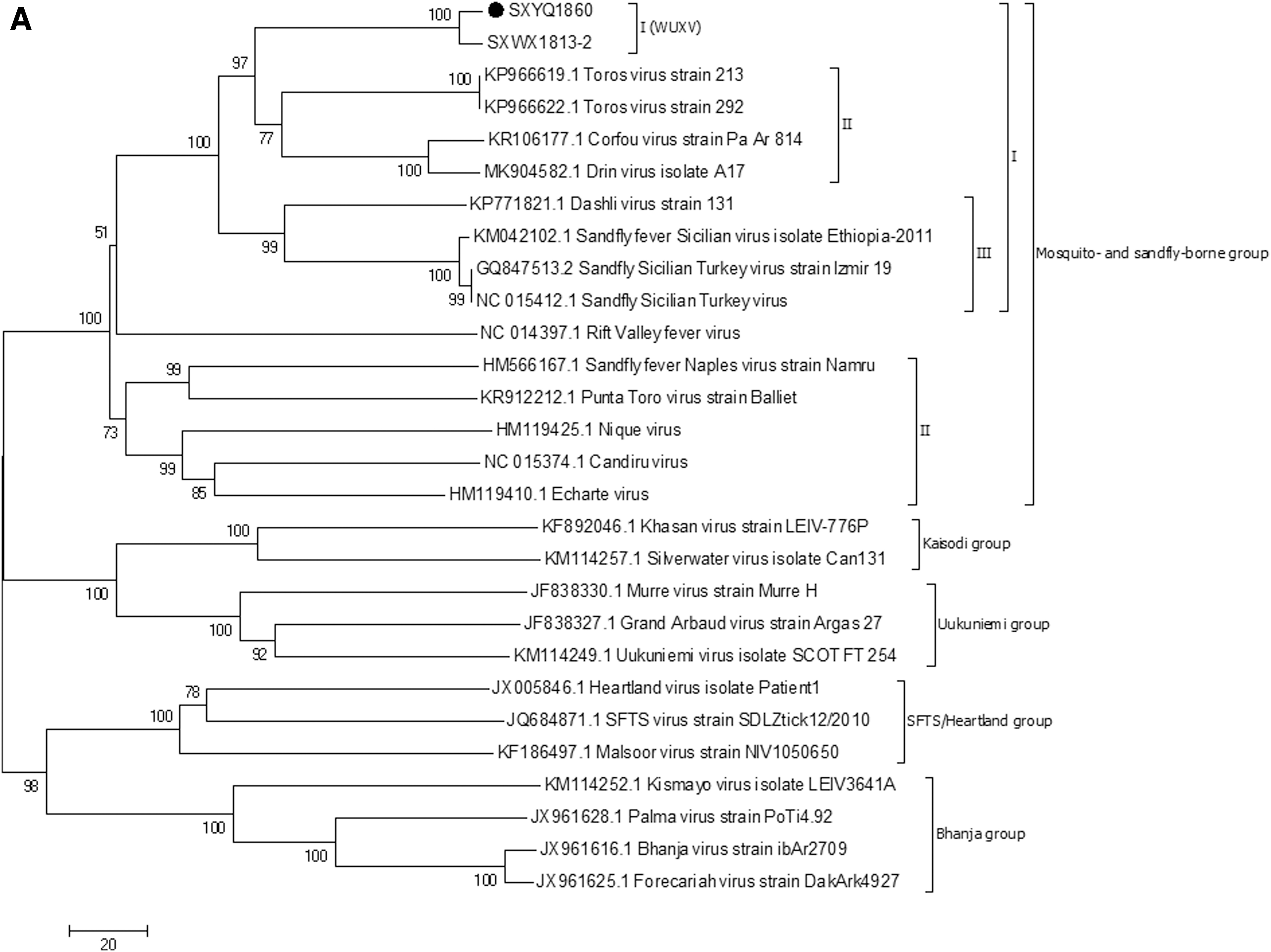
Molecular genetic evolution of the virus isolated from sandflies in Yangquan County.

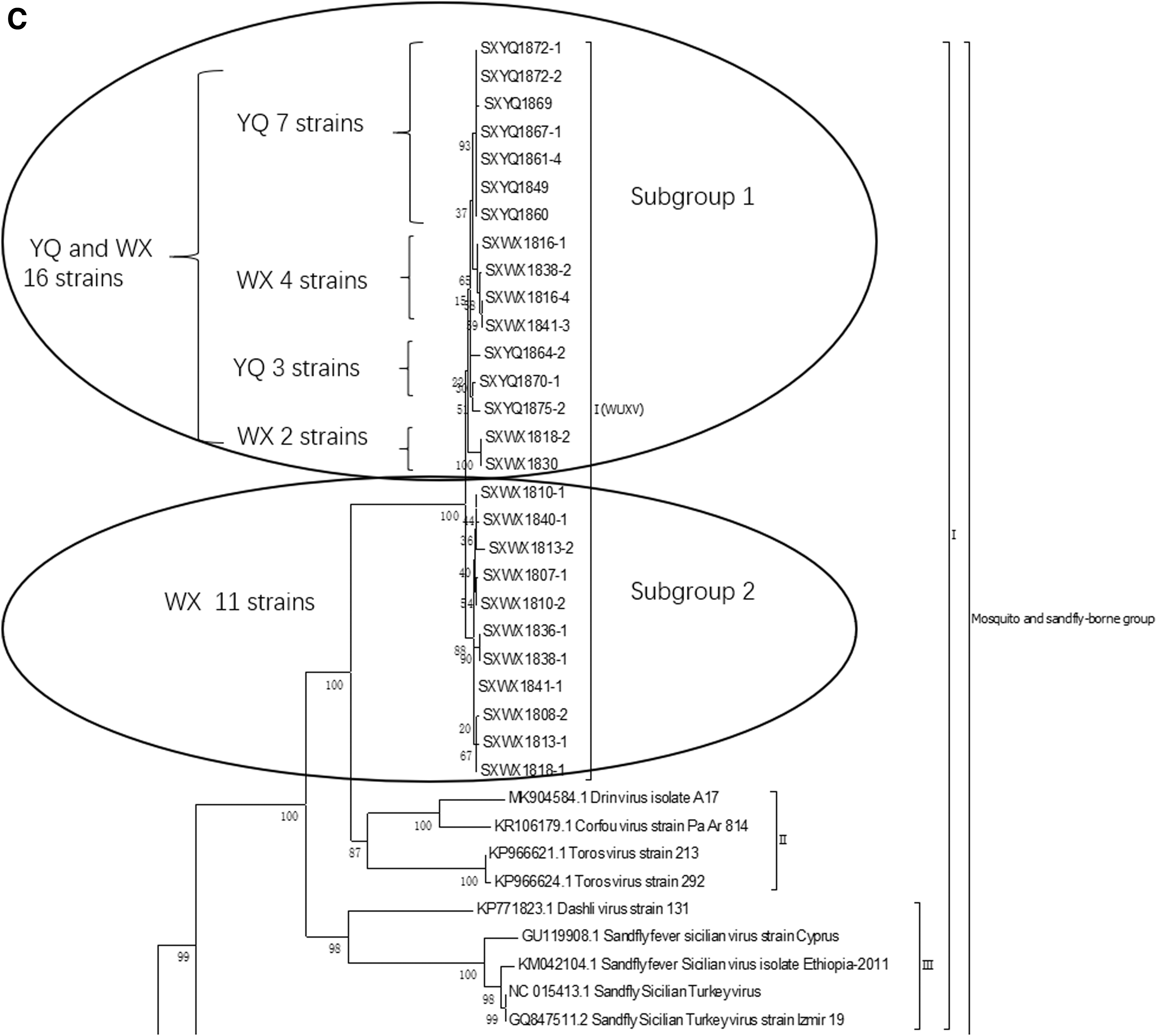

Phylogenetic analysis of the S gene showed that, independently of the phylogenetic analysis results for the NS and N protein gene nucleotide sequences, all 10 isolates obtained from sandflies in Yangquan County, including SXYQ1860, and the 17 strains of WUXV isolated from sandflies in Wuxiang County form a separate evolutionary branch (subbranch I) that is closely related to the Drin virus, TORV, and CFUV (subbranch II). This result suggests that the sandfly-borne virus isolated from Yangquan County and that isolated from Wuxiang County are the same virus, but that the 10 isolates isolated from Yangquan represent new strains of that virus.
Discussion
Previously, our research group reported that a new member of Phlebovirus, WUXV, was isolated from sandfly specimens collected in Wuxiang County, China (Wang et al. 2020). The present study reports that 10 strains of the same virus were isolated from sandflies collected in Yangquan County, Shanxi Province, China. Molecular genetic evolution analysis of the virus showed that these viruses belong to the WUXV group within the genus Phlebovirus and represent different isolates of WUXV from different regions (Wang et al. 2020).
Why can WUXV be isolated from sandflies in Yangquan County?
Yangquan County (112°5′–114°4′ east longitude, 37°40′–38°31′ north latitude) and Wuxiang County (112°26′–113°22′ east longitude, 36°39′–37°8′ north latitude) are located in the central and eastern parts of Shanxi Province, China, and the distance between the two areas is about 200 km (Fig. 1). Yangquan and Wuxiang counties are located in the central and western portions of the Taihang Mountains, and the geographical climate and natural environment of the two places are similar. The average altitude of the two sites is similar (between about 700 and 1700 meters), and both have a warm temperate continental climate with arid conditions and low rainfall (Wuxiang 2019, Yangquan 2019).
In addition, villagers in both places commonly raise sheep and chickens. Our research team used the same type and number of insect collectors in chicken and sheep pen environments in Wuxiang and Yangquan counties as collection sites.
On June 11–12, 3819 sandflies and 250 mosquitoes were collected in Wuxiang (Wang et al. 2020). In this study, we collected sandflies in Yangquan on June 13–14, obtaining 3996 sandflies and 74 mosquitoes. In both Wuxiang and Yangquan, natural populations of sandflies and mosquitoes are breeding in mid-June, and sandflies are denser than mosquitoes. In addition, the number of sandflies breeding in chicken and sheep pens in the two places is very similar. The numbers of breeding sandflies captured in chicken pens were 2665 (Yangquan) and 1604 (Wuxiang), and the breeding populations in sheep pens were 1029 (Yangquan) and 2032 (Wuxiang) (Wang et al. 2020).
Thus, the climate and natural environment of the Yangquan and Wuxiang areas are suitable for the growth and reproduction of sandflies. Therefore, the isolation of Phlebovirus (WUXV) from sandfly specimens collected in Yangquan is not surprising.
Molecular genetic characteristics of phleboviruses isolated in China
The results of molecular genetic evolution analysis of the viral genomes show that the genus Phlebovirus can be divided into five evolutionary groups, namely the mosquito- and sandfly-borne group, severe fever with thrombocytopenia syndrome (SFTS)/Heartland group, Bhanja group, Kaisodi group, and Uukuniemi group (Shen et al. 2018, Kuhn et al. 2020).
The mosquito- and sandfly-borne group can be further divided into two subgroups, of which the second subgroup contains sandfly fever Naples virus, transmitted by sandflies (Verani et al. 1980), and Rift valley fever virus, transmitted by mosquitoes (El Mamy et al. 2014). The first subgroup contains three evolutionary branches.
The first branch includes the SXYQ1860 virus isolated in this study and the WUXV isolated for the first time in China; the second branch contains the Toros (Alkan et al. 2016) virus and Corfou (Rodhain et al. 1985) virus. The third branch comprised sandfly fever Sicilian virus (Carhan et al. 2010) and Dashli virus (Alkan et al. 2017) (Fig. 4A). The SXYQ1860 virus isolated in Yangquan in this study is a member of Phlebovirus and forms a common evolutionary cluster with the WUXV (SXWX1813-2 virus) isolated previously (Fig. 4A: Viral L gene phylogenetic analysis and Fig. 4B: viral M gene phylogenetic analysis). Figure 4B and C show the molecular genetic evolution analysis results for 17 WUXVs (Wang et al. 2020) and 10 viruses isolated from Yangquan sandflies, indicating that the 27 isolates obtained from Chinese sandflies formed an independent evolutionary cluster. These 27 isolates are completely independent of epidemic phleboviruses of the Mediterranean, such as the TORV (Turkey) and CFUV (Greece) in the second cluster and sandfly fever Sicilian virus and Dashi virus in the third cluster. The virus isolated from sandflies in Yangquan and the virus previously isolated in Wuxiang are the same virus.
Although the currently WUXV isolates [17 from Wuxiang (Wang et al. 2020) and the 10 isolated in this study] are all closely related to Drin virus, CFUV, and TORV, and the length of gene sequence of WUXV is the same. However, it is obvious from the evolution analysis of the virus N gene (Fig. 4C) and NS gene (Supplementary Fig. S1) that WUXVs are divided into two subgroups. The number of isolates and the distribution of isolates in the two subgroups were completely different. The second subgroup of N gene analysis contained 11 isolates from Wuxiang County (Fig. 4C), while the second subgroup of NS gene evolution analysis only contained 2 isolates from Wuxiang County (SXWX1816-1 and SXWX1841-3), which were in the first subgroup of N gene evolution analysis (Fig. 4C).
In addition, the branch length of different WUXV isolates in the evolutionary tree was also very different, suggesting that there were evolutionary differences among different virus isolates. In conclusion, WUXV is a group of sandfly-borne virus that is constantly mutating.
The Mediterranean coast is hot and dry in summer, and warm and humid in winter, with an average temperature in the coldest month of 4–10°C. Winter is rainy, whereas summer is dry with bright sun, and annual precipitation is 300–1000 mm, defining a unique geographic climate, namely the Mediterranean climate (Giorgi and Lionello 2008). Wuxiang and Yangquan are located in central China. The winter is cold and dry, with temperatures below 0°C, and summer is hot and rainy. The annual rainfall is about 1000 mm, with two thirds of precipitation occurring in summer. Thus, the Mediterranean area and central Shanxi Province in China have common characteristics of cold winters, hot summers, drought and low rainfall, which are suitable geographic and climatic conditions for the breeding of sandflies. The Mediterranean region and Shanxi Province in central China are nearly 10,000 km apart.
However, WUXV isolated from a sandfly sample from Shanxi Province in China had a very close genetic evolutionary relationship with the TORV (Turkey) and CFUV (Greece), which are epidemic to Mediterranean countries.
Biological characteristics between SXYQ1860 and SXWX1813-2
We analyzed the virus isolated from Yangquan (SXYQ1860) and that virus isolated from Wuxiang (SXWX1813-2) in parallel to examine their viral biological characteristics. These phenotypic characteristics are as follows: they showed the same CPE timing and characteristics in BHK-21 cells; they had similar viral plaque characteristics and the same particle size; and the time between the onset and death after viral infection of suckling mice is not much different (Table 2). Thus, it indicates that the two virus isolates are the same virus and should both be considered isolates of WUXV from different geographic regions.
In this study, WUXV was isolated from sandflies collected in the wild in Yangquan County. Our results suggest that WUXV may have a wide geographical distribution except Wuxiang county. Therefore, it is of great public health significance to advance the detection and monitoring of local sandfly-borne pathogens and determine the potential infection of humans and animals.
Footnotes
Authors' Contributions
J.W., N.F., and S.F. involved in drafting the article. J.W., N.F., S.F., and G.L. involved in revising the article. J.W., N.F., S.F., J.C., B. Wu, Z.X., J.S., X.T., Y.L., Y.H., F.L., and S.X. participated in the collection of sandfly specimens and acquisition of data. J.W., N.F., S.F., and G.L. participated in the analysis of data. J.W., N.F., S.F., and G.L. contributed to the interpretation of data. H.W., B. Wang, and G.L. contributed to conception and design of the article. H.W., B. Wang, J.C., and G.L. supported funding for the study.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This work was supported by the Ministry of Science and Technology of the People's Republic of China (2018ZX10711001, 2018ZX10734-404-003); National Key Research and Development Program (No. 2018YFA0900800); Key Research and Development (R&D) Projects of Shanxi Province, China (201803D31205); Development Grant of the State Key Laboratory of Infectious Disease Prevention and Control (2014SKLID103, 2015SKLID505).
Supplementary Material
Supplementary Figure S1
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.