
Abstract
The family Rhabdoviridae contains diverse viruses, including vector-borne and nonvector-borne viruses, some that are human pathogens, including rabies virus and also nonpathogenic viruses. Bats, which are a known reservoir of viruses with zoonotic potential including coronaviruses, also carry multiple rhabdoviruses such as but not limited to lyssaviruses. We collected samples from 193 insectivorous and frugivorous bats in the Republic of the Congo and tested them for rhabdovirus RNA. Four samples were found positive for viral RNA representing sequences of four different, not previously described rhabdoviruses. Although phylogenetic and taxonomic placement of the novel sequences is uncertain, similarities with previously detected rhabdovirus sequences in bats suggest that these could represent vertebrate viruses. Considering the pathogenic risks some rhabdoviruses pose for humans, these results highlight the need for more research and surveillance regarding rhabdoviruses and bats.
Introduction
The negative sense RNA
Although the biology of many of the rhabdoviruses discovered to date remains largely unknown, some of them, such as rabies virus and other lyssaviruses are well known as pathogens causing fatal disease in humans and other animals (Shope 1982, Banyard et al. 2014, Hayman et al. 2016). One reservoir and known source of zoonotic infections with rhabdoviruses are bats, which are known to harbor and shed a rich set of not only rhabdoviruses, but viruses in general, often without themselves displaying any overt clinical signs of infection (Murphy et al. 1970, Iwasaki et al. 2004, Ghedin et al. 2013, Luis et al. 2013, Blasdell et al. 2014, Binger et al. 2015). Considering the proposed role of bats as reservoirs, we undertook testing of bats in the highly biodiverse Congo Basin for the presence of rhabdovirus RNA to further asses rhabdovirus diversity.
Materials and Methods
Bat “capture and release” specimen collection was conducted by mist-netting at heights of 1–3 meters along trails and streams in lowland forest. Nonlethal sample collection for the Emerging Pandemic Threats (EPT) program's PREDICT project was approved by the Institutional Animal Care and Use Committee (IACUC) of the University of California Davis (UC Davis) and the government of the Republic of the Congo (permit number 018/MRSIT/DGRST/DMAST issued by the Republic of the Congo Ministry of Scientific Research and Technical Innovation). Some samples analyzed in this study were collected as part of a faunal inventory, commissioned in association with a proposed iron ore mine feasibility study (Bates et al. 2013). As part of that study, a limited number of bats (n = 17) were sacrificed as voucher specimens and samples were also collected for the PREDICT project. Bat species were identified based on field guides and verified by Paul Bates of the Harrison Zoological Museum, Sevenoaks, Kent, United Kingdom (Kingdon 2005, Monadjem et al. 2010).
Oral and rectal swabs were collected from live and voucher animals using sterile micro-tipped polyester swabs (Puritan™ Medical Products Company, LLC), placed in 1.5 mL NucliSens® lysis buffer (bio-Mérieux, Inc.) in 2.0-mL cryotubes. Tissue samples from voucher animals were collected in 2.0-mL cryotubes without medium. All were stored at ambient temperature for up to 3 h before being frozen in liquid nitrogen and later transferred to −80°C until further processing.
RNA was extracted either manually using Trizol®, or using a Qiagen Viral RNA Mini Kit and stored at −80°C. Afterward RNA was reverse transcribed into cDNA using a Maxima H Minus First Strand cDNA Synthesis Kit (Thermo Scientific) and stored at −20°C until analysis. A semi-nested consensus touchdown PCR, amplifying ∼260 nucleotides of the conserved DNA polymerase (L) gene, was conducted (Supplement Data S1). PCR products were examined by gel electrophoresis and products excised, cloned, and Sanger sequenced at the UC Davis DNA sequencing facility. Obtained sequences were edited in Geneious and compared with the GenBank database (BLAST).
A phylogenetic tree was constructed based on the amino acid sequences of the DNA polymerase gene, including the PCR amplicons and 48 published rhabdovirus L-gene sequences; cytorhabdoviruses served as outgroup. First multiple sequence alignments were made in Geneious (Muscle Alignment), Bayesian phylogeny was inferred using MRBAYES (version 3.2), with default parameters and four chains of 1,000,000 generations, and tree sampling every 1000 steps during the process to monitor phylogenetic convergence. The final average standard deviation of split frequencies was <0.03. The first 10% of the trees were discarded, and the remaining trees combined using TreeAnnotator (version 2.5.1) and displayed with FIGTREE (1.4.4).
Results
Samples were collected between July 2012 and March 2014 in the Lékoumou and Sangha departments of the Republic of the Congo and included 259 samples from 193 bats belonging to at least 23 different species (Supplementary Data S2 and S3). The collected samples were primarily rectal (140) and oral swabs (93), but also included lung (20), brain (4), liver (1), and feces (1). The female/male ratio was 111/78, with four individuals undetermined. Rhabdovirus RNA was detected in the oral swab samples of four adult animals, accounting for an overall 2.1% apparent prevalence of rhabdovirus RNA in the bats sampled. All rhabdovirus RNA positive samples were collected within 1 week in the same area during the dry season (Supplement Data S2 and S3). Two of these bats were Hipposideros ruber, one a Triaenops afer and one a Neoromicia brunnea. All four obtained sequences were different from each other, sharing no more the 67% nucleotide identities. Closest matches upon protein BLAST shared between 70% and 80% identities with mostly other isolates of unclassified rhabdoviruses isolated from bats. The closest classified matches belonged to Vesicolovirus (ZB12028, ZB12058, and ZB12064) or Perhabdovirus (ZB12070) genera. Upon phylogenetic analysis, the detected sequences largely branch off at the base of classified genera branches, two at the base of the Caligrhavirus branch, and one at the base of the Sigmavirus branch, whereas one showed no close connection with any established genera (Fig. 1). Novel sequences are stored in GenBank with accession numbers MG956843–MG956846 (Supplement Data S4).
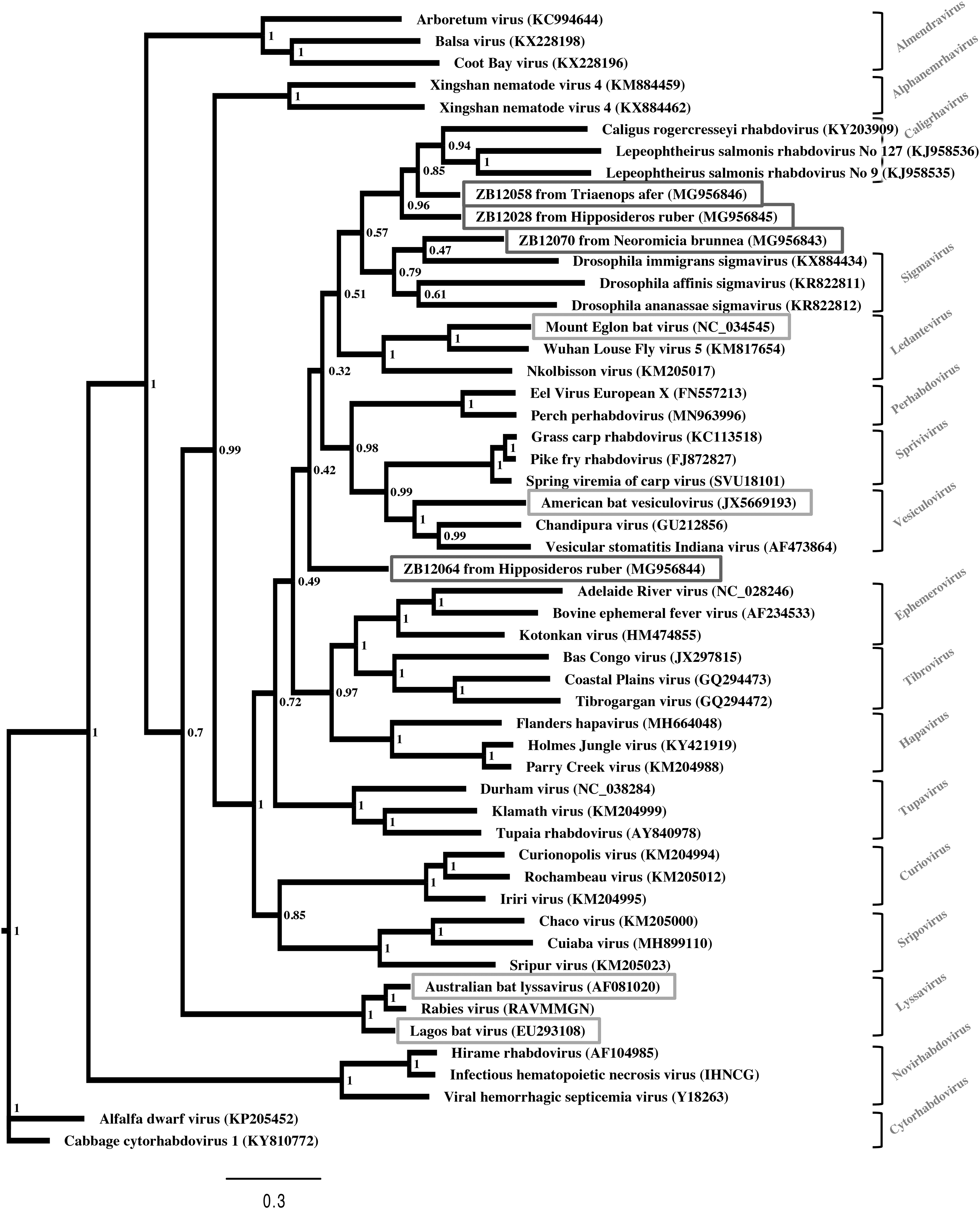
Maximum likelihood phylogenetic tree of rhabdoviruses based on the L gene. The four novel sequences (dark boxes), previously known sequences of 4 other bat (light gray boxes) and 44 non-bat rhabdoviruses are indicated. Virus name and GenBank accession numbers are shown and, genus and host association according to the ICTV indicated. Bootstrap support indicated at nodes.
Discussion
We have detected RNA indicative of four rhabdoviruses in samples collected from bats native to the Congo Basin. None of them matches any available sequence from the 191 previously described rhabdovirus species. Full genome sequencing was not possible, which is a limitation of the study. Given the brevity of the RNA sequences amplified definite phylogenetic and taxonomic placement was, therefore, not possible; however, comparison with the GenBank database and the clustering upon phylogenetic analysis suggest that all of the respective viruses may represent new species. Considering the apparent basal or unclear placement in the phylogenetic tree some or all of them might potentially not be associated with any of the 30 established rhabdovirus genera.
Since all sequences were amplified from oral swabs from insectivorous bats, it is possible that some or all of the detected sequences may either belong to viruses with arthropod vectors or viruses primarily infecting insects. However, virus containing saliva of bats and other animals is a known source of zoonotic lyssavirus transmission. Considering the protein similarity to other rhabdovirus sequences isolated from bats, and similarities to other better known rhabdoviruses, we believe that the detected sequences may belong to vector- or nonvector-borne vertebrate rhabdoviruses.
Although we do not have any information with regard to the biology or possible zoonotic potential of the viruses of which we detected RNA, it is known that not only lyssaviruses but also other types of rhabdoviruses can or are suspected to cause human disease (Gurav et al. 2010, Grard et al. 2012). We consequently believe that further research into, and surveillance of rhabdoviruses, particularly in bats, is warranted to better understand rhabdovirus evolution and to evaluate and mitigate potential risks bat rhabdoviruses (other than lyssaviruses) may pose.
Footnotes
Acknowledgments
The authors thank the government of the Republic of the Congo for the permission to conduct this study; the staff of the Wildlife Conservation Society, especially Ashley Vosper; the staff of the University of California in Davis; the staff of Metabiota, especially Prime Mulembakani; and any other involved members of the PREDICT-1 consortium (
Disclaimer
The contents are the responsibility of the authors and do not necessarily reflect the views of USAID or the U.S. Government.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
The study was undertaken as part of the global USAID-funded Emerging Pandemic Threats (EPT) PREDICT project, which focuses on enhancing the global capacity for the detection and discovery of potentially zoonotic viruses at the human–animal interface. It was made possible primarily by the generous support of the American people through the United States Agency for International Development (USAID) Emerging Pandemic Threats PREDICT program (co-operative agreement number GHN-A-OO-09-00010-00).
Supplementary Material
Supplementary Data S1
Supplementary Data S2
Supplementary Data S3
Supplementary Data S4
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.