
Abstract
Scientists collect fleas (Siphonaptera) to survey for Yersinia pestis, the bacterial agent of plague. When studying fleas parasitizing prairie dogs (Cynomys spp.), two primary methods are used: (1) combing fleas from live-trapped prairie dogs and (2) swabbing fleas from burrows with cloth swabs attached to metal cables. Ideally, burrow swabbing, the cheaper and easier method, would explain flea burdens on prairie dogs and provide reliable information on plague prevalence. In a linear regression analysis of data from 1-month intervals (June–August 2010–2011) on 13 colonies of black-tailed prairie dogs (Cynomys ludovicianus, BTPDs) in New Mexico, flea abundance on swabs explained 0–26% of variation in BTPD flea burdens. In an analysis of data (May–August 2016) from six colonies of BTPDs in Montana, flea abundance on swabs explained 2% of variation in BTPD flea burdens. In an analysis of data from a short-term interval (July 23–27, 2019) on four colonies of BTPDs in Montana, flea abundance on swabs explained 0.1% of variation in BTPD flea burdens. In an analysis of data from 1-week intervals (August–October 2000) on four colonies of white-tailed prairie dogs (Cynomys leucurus, WTPD) in Utah, swabbing data explained 0.1% of variation in WTPD flea burdens. Pools of fleas from two WTPD colonies were tested for Y. pestis by mouse inoculation and isolation; 65% from WTPDs tested positive, whereas 4% from burrows tested positive. Data herein also show that results from burrow swabbing can misrepresent flea species composition and phenology on prairie dogs. Burrow swabbing is useful for some purposes, but limitations should be acknowledged, and accumulated data should be interpreted with caution.
Introduction
Plague is a lethal bacterial disease of mammalian hosts and flea vectors. In the western United States, plague negatively affects a variety of rodents, including multiple species of sciurids (Gage and Kosoy 2006, Mahmoudi et al. 2020, Barbieri et al. 2021). Some sciurids amplify plague, providing conditions for disease spillover to sympatric rodents, lagomorphs, carnivores, and humans (Eskey and Haas 1940, Barnes 1982). Rates of plague transmission are expected to increase as flea burdens increase (Lorange et al. 2005, Bosio et al. 2020). Thus, it is important to acquire a deeper understanding of flea parasitism among sciurids using appropriate, representative sampling.
Many types of sciurids excavate burrows and use the labyrinths as refuge against predators and environmental conditions (Reichman and Smith 1990). Three primary techniques have been used to capture fleas parasitizing burrowing sciurids. First, burrowing sciurids are live-trapped, anesthetized, and combed for fleas (Gage 1999). This method requires traps and, in some cases, intensive effort; that said, it provides a direct measure of flea burdens and, therefore, is highly useful (Gage 1999).
Second, shovels and hydraulic machines have been used to excavate sciurid burrows and collect fleas. This approach is useful because some flea species spend significant amounts of time off sciurids, in burrows (“nest fleas”; Krasnov et al. 2004). Burrow excavations are time and labor intensive, and they destroy valuable shelters for rodents and other organisms. Excavations are typically avoided.
Third, burrows can be swabbed for fleas (Stewart and Evans 1941, Maupin 1970). A flannel cloth is clipped to the end of a metal flexible plumber's snake and inserted into a burrow as deep as possible. The cable is used to shake the swab, which purportedly stimulates host-seeking by fleas. The swab is removed from the burrow and, commonly, placed in a plastic bag and frozen to kill fleas, which are counted in a field station or laboratory (Gage 1999). This technique is relatively rapid and inexpensive, leaves burrows intact, and allows for repeated sampling over time.
Ideally, burrow swabbing would accurately and precisely index the abundance of fleas on sciurids. Swabbing suffers from at least two limitations in this context. First, some flea species spend most (or all) of their time on hosts. These “fur fleas” (Krasnov et al. 2004) are presumably difficult to collect from burrows. Second, biologists sometimes have trouble inserting swabs into the deep components of burrows (Eads 2017) where fleas presumably spend most of their time when off hosts (Krasnov 2008). Depending on the study system, swabbing might fail to index flea burdens on sciurids and might even provide misleading information on seasonal changes in flea parasitism of hosts (Seery et al. 2003).
From a surveillance perspective, it would be useful if burrow swabbing yielded fleas that, when tested, provided accurate information on Y. pestis prevalence. When a rodent dies, fleas leave the host carcass and quest for live hosts (Poland and Barnes 1970). If the rodent died from plague and fleas were infected by Y. pestis, and the timing is right, biologists might collect Y. pestis-infected fleas that test positive when applying standard diagnostic procedures.
In this study, we analyze data from prairie dogs (Cynomys spp., PDs), which are colonial burrowing sciurids. The swabbing technique is commonly used to study fleas and survey for Y. pestis on PD colonies (e.g., Salkeld and Stapp 2008, Griffin et al. 2010, Jones and Britten 2010, St. Romain et al. 2013, Bron 2017, Tripp et al. 2017, Biggins and Eads 2019, Poje et al. 2020). Here, we compare data from live capture of fleas on prairie dogs to coincident burrow swabbing to determine to what extent burrow sampling represents animal burdens, and we provide recommendations for flea collection and plague surveillance on PD colonies.
Materials and Methods
Study sites
In a planned study of data from swabbing and combing, we sampled black-tailed PDs (Cynomys ludovicianus, BTPDs) and their fleas at the Vermejo Park Ranch, New Mexico, from June–August 2010–2011 (Eads 2014). We conducted research on 1.54 or 2.25 ha plots distributed among 13 colonies (n = 19 plots in 2010 and 20 plots in 2011). We categorized the plots into five groups of two to five (based on proximity) and sequentially sampled the groups in random order during 10-d work periods. During a field day, we trapped BTPDs on some plots within a group and swabbed burrows on the remaining plots. The following day, we switched efforts. Each plot was sampled at least twice per month (comb and swab).
In a post hoc assessment, we analyzed data on BTPDs and their fleas on six plots (∼13 to 22 ha) on six colonies at Charles M. Russell (CMR) National Wildlife Refuge, Montana, from May to August 2016. Trapping and swabbing were completed for separate study objectives (trapping: Rocke et al. 2017, Russell et al. 2018, swabbing: Poje et al. 2020), and on different dates (Table 1), but the overall effort allowed for a coarse-scale comparison of combing and swabbing data from summer 2016.
Fleas on Burrow Swabs and on Black-Tailed Prairie Dogs, May–August, 2016, on Six Colonies, Charles M. Russell National Wildlife Refuge, Montana
In another post hoc assessment, we analyzed data from five areas on four BTPD colonies between July and August 2019 at CMR. Sampling areas on three colonies were selected according to suspected plague circulation, indicated by visual observation of BTPD densities and activity from 2018 through 2019. Categories and areas of suspected plague circulation included (1) epizootic in 2018, with subsiding declines and stabilized, low abundance of BTPDs in 2019 (area decline from 197 to 22 ha), (2) active, ongoing epizootic decline during 2019 (from 35 to 5 ha), and (3) stable with no apparent decline in BTPD abundance or occupied area in 2018 or 2019 (3 areas of 28, 67, and 6 ha). Trapping on the first four areas occurred during 23–24 July followed by swabbing on 25 and 27 July. Trapping on the 5th area (6 ha), 9 km away from the other four areas, occurred during 8–9 August followed by swabbing on 10 August. Except for the colony sampled in August 2019, fleas were counted and frozen in vials with saline for future identification and plague testing.
Also post hoc regarding comparisons of swabbing and combing, we sampled white-tailed PDs (Cynomys leucurus, WTPDs) and their fleas at Coyote Basin, Utah, from August to October 2000 (Biggins et al. 2010). Fleas were swabbed from burrows and combed from WTPDs on 16 plots (4 ha each) distributed among 11 colonies. Sampling occurred during ∼1-week intervals: 1–6 August and 6–10 October for one colony; 7–11 August and 6–10 September, and 12–20 October for seven adjacent colonies combined into a single site; 12–18 August and 20–25 October for one colony; and 23–29 August and 20–25 October for two adjacent colonies combined into a single site. For the site with seven adjacent colonies, we combined data from August 7–11 to September 6–10 so all areas were characterized by an early sampling period (August–September) and a later sampling period (October) (with implications for an analysis of temporal changes in flea densities from burrows and WTPDs).
Plague kills PDs and fleas, thus reducing flea habitat and densities; plague transmission therefore confounds studies of flea abundance (Eads et al. 2016) due to consequent changes in flea-host ratios (Biggins and Eads 2019). Plague was likely absent from our sampling areas at Vermejo (Eads et al. 2016). Plague is seemingly maintained at CMR (Matchett et al. 2010), similar to other sites with endemic plague in the western United States (Lowell et al. 2015). Epizootic plague as defined by Biggins and Eads (2019) was suspected in some cases at CMR. Plague is endemic and was detected at Coyote Basin, but epizootic plague was not observed on our sampling areas (Biggins et al. 2010).
Pulicides can be used to suppress fleas and manage plague on PD colonies (Seery et al. 2003, Biggins et al. 2010, Matchett et al. 2010, Hoogland et al. 2018, Eads et al. 2019, 2020, 2021). At Vermejo, DeltaDust® (0.05% deltamethrin; Bayer Environmental Science, Research Triangle Park, NC) had not been used for at least 5 years on the study colonies. Two of the BTPD colonies at CMR sampled in 2016 had never received any flea control treatment and the other four, plus the colony sampled in August 2019, were last treated with Pyraperm (pyrenone, 4–0.05 permanone 0.5 dust; Bayer Environmental Science, Montvale, NJ) 19 years before this study. The three BTPD colonies at CMR sampled in July 2019 were last treated with DeltaDust® 8 years before this study. The effect of DeltaDust® on fleas at PD colonies usually wanes after 1–2 years (Eads and Biggins 2019). We restricted the Coyote Basin data to colonies that had never been treated with DeltaDust®, or to sampling periods before a colony was first treated.
Swabbing burrows for fleas
At Vermejo, the number of swabbing events per plot, per day, was standardized at 16.3 × ha−1. Burrow transects suggested we swabbed
At CMR in 2016, ∼100 randomly selected burrows were targeted per plot during two seasonal “sessions” (May–June and July–August; Poje 2019, Poje et al. 2020). Additional sampling was completed if possible, such that >200 burrows were swabbed on some plots (Table 1). Each burrow was swabbed as described above, except each flannel cloth was 15 × 15 cm. The percentage of burrows swabbed (
On the CMR colonies with areas of suspected differing plague circulation in 2019, 202 randomly selected burrows within the areas trapped were swabbed (each flannel cloth was 28 × 28 cm) a few days after trapping in July 2019. One hundred randomly selected burrows were swabbed on the third (6 ha) no decline area 2 days after trapping was completed. Burrow density data were not collected on these plots; thus, we cannot index the percentage of burrows swabbed. Swabs were bagged and frozen. Fleas were counted in a laboratory and stored frozen in vials with saline for future identification and plague testing (not completed as of article completion).
At Coyote Basin, randomly selected burrows (in areas with observed WTPD activity) were swabbed (20 × 20 cm swabs) on a single day within each of the 1-week intervals noted above. The number of swabbing events per plot (or colony) per interval varied from 21 to 25 (
Trapping and combing prairie dogs for fleas
We combed fleas from live-trapped, anesthetized PDs. At Vermejo, each plot contained 25 or 37 single-door live traps (16.3 traps × ha−1; Tomahawk Live Trap, Hazelhurst, WI). We anesthetized each BTPD with isoflurane in an induction chamber, removed the BTPD, and combed its body as thoroughly as possible for 45 s over plastic tubs (Eads et al. 2013, 2015). We collected and counted the fleas and stored them in vials with saline. We released each BTPD at its trapping location. Fleas were identified to species (Eads 2014).
In 2016, at CMR, each plot contained 80 single-door cage traps (Tru-Catch Traps, Belle Fourche, SD) during the first trapping session and 160 traps during the second trapping session. Fleas were similarly collected from anesthetized BTPDs during 30-s combings. Fleas were counted, identified to species, and tested for plague (Rocke et al. 2017, Russell et al. 2018).
In July 2019, at CMR, 40 traps were placed in each of the 4 areas of varied suspected plague activity. BTPDs observed above ground were scarce in the area of the ongoing decline in 2019. In August 2019, 80 traps were used on the 6-ha plot. Fleas were similarly collected from anesthetized BTPDs during 30-s combings plus an additional 30-s manual body inspection. Fleas were counted, identified to species, and stored frozen in saline for later plague testing (not completed as of article completion).
At Coyote Basin, WTPDs occurred at low densities. Consequently, traps were clustered at active WTPD burrows. Fleas were collected and counted during 30-s combings. Fleas were stored frozen in vials with saline for identification and plague testing through mouse inoculation and isolation (Biggins et al. 2010).
Data analyses
BTPDs in New Mexico
We summarized mean flea abundance (total fleas collected divided by number of BTPDs sampled, Krasnov et al. 2004, 2006a, b) with the Vermejo data separated by month and year. We used least-squares regression, the lm function, and F-tests in Program R ( × 64 version 3.6.1; R Core Team 2019) to evaluate the correlation between paired values of flea abundance on swabs (predictor) and BTPDs (response). The number of animals sampled can affect measures of parasite abundance (Poulin 2011). Thus, we included the number of BTPD combing events as a control. We also included a two-way interaction between year and flea abundance on swabs, due to known data; fleas were much more abundant in 2011 (Eads et al. 2016). We wondered if the swabbing data would provide reliable information on the relative abundance of flea species on BTPDs. We summarize abundance data for two flea species that predominated: Oropsylla hirsuta (a PD specialist) and Pulex simulans (a mammal generalist). Other flea species comprised <0.5% of fleas collected (thus, inference would be limited at best).
BTPDs in Montana
With the data from summer 2016, we summarized and compared mean flea abundance on swabs and BTPDs with the data grouped by plot. We used least-squares regression to evaluate the correlation between paired values, with the number of combing events as a control. All fleas from swabs and BTPDs were identified to species. Oropsylla hirsuta and P. simulans predominated; we summarize their abundance on swabs or BTPDs as percentages of all fleas identified.
With the data from 2019, we summarized and compared mean flea abundance on swabs and BTPDs with the data grouped by suspected plague circulation. We used least-squares regression, as described above, to evaluate the correlation between paired values. We summarized the data in Table 2. We summarized the abundance of flea species as percentages of all fleas identified.
Area Occupied by Black-Tailed Prairie Dogs, Fleas on Burrow Swabs and on Black-Tailed Prairie Dogs, July–August, 2019, in Areas of Suspected Differing Plague Activity, Charles M. Russell National Wildlife Refuge, Montana
WTPDs in Utah
We used least-squares regression to examine the correlation between paired values of flea abundance from swabs and WTPDs during ∼1-week intervals, with the number of combing events as a control. With BTPDs at Vermejo, monthly changes in flea abundance from swabs (June–August each year) were mirrored by temporal changes in flea abundance on BTPDs; regardless of the technique, flea abundance increased from June to July, and declined slightly into August (Eads 2014). To determine if the same consistency applied to data from WTPD colonies, we evaluated temporal changes in flea abundance on swabs and WTPDs from August to September (n = 15 plots on the four sites) into October 2000 (n = 5 plots, only one of which was not sampled during August–September). Finally, in an attempt to survey for Y. pestis among fleas, 26 pools of fleas from individual burrows and 54 pools of fleas from individual WTPDs were tested for Y. pestis through mouse inoculation and isolation. All pools were collected in August 2000 (1–23 fleas per burrow swab pool and 1–32 fleas per WTPD comb pool). Each pool contained either O. hirsuta or P. simulans. For each flea species separately, we used the prop.test in Program R to compare the proportion of flea pools from WTPDs and burrows testing positive for Y. pestis.
Results
BTPDs in New Mexico
In 2010, we combed BTPDs on 650 occasions and collected 1448 fleas. On the same plots, we swabbed 2201 burrows and collected 1183 fleas. The following year, we combed BTPDs 631 times and collected 5233 fleas. We swabbed burrows 2765 times in 2011 and collected 5035 fleas. Oropsylla hirsuta and P. simulans comprised about 46% and 54% of fleas combed from BTPDs (Eads 2014) versus 89% and 11% of fleas swabbed from 350 randomly selected burrows (Eads 2017). This comparison might suggest burrow swabbing underestimates the relative abundance of P. simulans on BTPDs, but only a subset of swabbed fleas was identified.
The 2-way interaction between year and flea abundance on swabs justified separation of data by year (p = 0.057). Using data from 2010, we failed to detect a relationship between flea abundance on swabs and flea abundance on combed BTPDs (p = 0.726). In contrast, using data from 2011, when fleas were more abundant overall (Eads et al. 2016), flea abundance on swabs correlated positively with flea abundance on BTPDs, although the correlation was relatively weak (Fig. 1; r 2 = 0.30, p < 0.001). Explanatory power (r 2) declined to 0.26 when the number of combing events was excluded.
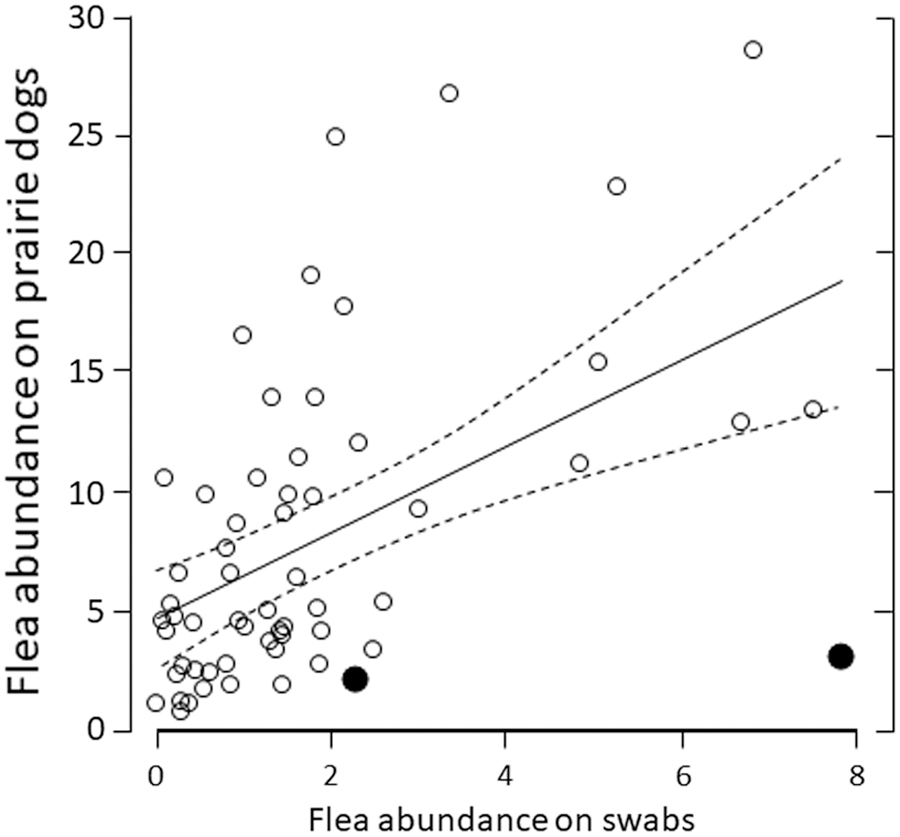
Correlation between flea abundance on burrow swabs (horizontal axis) and flea abundance on black-tailed prairie dogs (vertical axis) during June–August, 2011, Vermejo Park Ranch, New Mexico. Dots represent sampling plots. The unbroken line depicts a relationship derived from linear regression and dashed lines depict 95% confidence limits. In two cases (larger black dots), flea abundance was higher on swabs than on prairie dogs.
BTPDs in Montana
In 2016, we combed BTPDs on 661 occasions and collected 1205 fleas. In the same areas, we swabbed 1597 burrows and collected 139 fleas (Table 1). We failed to detect a relationship between flea abundance on swabs and flea abundance on BTPDs (p = 0.626). Oropsylla hirsuta and P. simulans comprised 64% and 36% of fleas combed from BTPDs, versus 83% and 6% of fleas swabbed from burrows. Thus, burrow swabbing underestimated the relative abundance of P. simulans on BTPDs.
In 2019, we combed BTPDs on 153 occasions and collected 256 fleas (75% O. hirsuta and 24% P. simulans based on 156 flea identifications). In the same areas, we swabbed 302 burrows and collected 64 fleas, all O. hirsuta. We failed to detect a relationship between flea abundance on swabs and flea abundance on combed BTPDs (p = 0.156). Flea abundance varied among areas of colonies with differing degrees of suspected plague circulation in 2019 (Table 2). Fleas were most abundant on small numbers of BTPDs inhabiting the area of observed ongoing decline (60 fleas on 9 BTPDs). In that same area, just 3 fleas were swabbed from 35 burrows. Very few fleas were found in areas where the massive die-off in 2018 appeared to have stabilized in 2019. Fleas were moderately abundant on BTPDs, but scarce in burrows, where no decline was observed from 2018 through flea sampling in 2019 (Table 2).
WTPDs in Utah
We combed WTPDs on 121 occasions; 2338 fleas were found. On the same colonies, we swabbed 455 burrows; 321 fleas were found. With data from 1-week intervals, we failed to detect a relationship between flea abundance on swabs and flea abundance on WTPDs (p = 0.778). As the study progressed from August–September to October, flea abundance increased by 86% on WTPDs (August–September
Sixty-seven percent of O. hirsuta pools (n = 27 with 157 fleas) from WTPDs during August 2000, when epizootic plague was not suspected, tested positive for Y. pestis. In contrast, the bacterium was not detected in O. hirsuta from burrows (n = 12 with 171 fleas, p < 0.001). Similarly, 63% of P. simulans pools from WTPDs tested positive (n = 27 with 280 fleas) compared to 7% of P. simulans pools from burrows (n = 14 with 263 fleas, p = 0.001).
Discussion
Implications for studies of fleas
Our research, conducted over 5 years across three U.S. states with two PD species, provides insight into limitations of burrow swabbing on PD colonies to assess flea population levels, species composition, and Y. pestis prevalence. We failed to detect any meaningful relationship between flea abundance from swabbed burrows and combed PDs. If we combined all data to increase statistical power, flea abundance from burrows was positively correlated with flea abundance from PDs (p < 0.001), but the correlation was relatively weak (r 2 = 0.13). In many cases, we swabbed very few fleas from burrows even though PDs observably harbored large numbers of fleas (Fig. 1, Tables 1 and 2). If fleas are collected from burrows only, fleas can appear relatively scarce, despite large numbers of fleas on some (or many) PDs.
Burrow swabbing data provided different, and potentially misleading, information on the relative abundance of different flea species, compared to on-host assessments. Burrow swabbing data regularly underestimated the on-host relative abundance of P. simulans compared to O. hirsuta. Similarly, in the study of Tripp et al. (2017), O. hirsuta and P. simulans represented 85% and 0.3% of fleas on swabs versus 73% and 27% of fleas on BTPDs. In the study of Holmes (2003), O. hirsuta and P. simulans represented 41% and 44% of fleas on swabs versus 18% and 75% of fleas on BTPDs. Perhaps P. simulans is very much a fur flea, spending little time off hosts when hosts are available. Oropsylla hirsuta is a purported fur flea (Krasnov et al. 2004), although we emphasize additional study is needed for confirmation; if O. hirsuta is more of a nest flea, on-host combings may underestimate true abundance. Pulex simulans is relatively small and is more difficult to comb from PDs than O. hirsuta (Eads et al. 2015); a similar detection limitation may apply to burrow swabbing (Poje et al. 2020).
Swabbing and combing suggested differing trends for temporal changes in flea abundance on WTPDs. The results mirror observations by Seery et al. (2003). In both cases, flea abundance and prevalence increased from summer through fall on combed PDs, but declined in burrow swab samples. This discrepancy, potentially resulting from variation in flea behaviors (e.g., time spent on hosts, questing for hosts, or retreating deeper into burrows during hot/dry conditions and becoming less available for “capture”), reduces confidence that swabbing reliably, or accurately, indexes changes in flea parasitism over time, often considered a key attribute for research, management, and plague risk assessments (Eisen et al. 2019).
One might expect a large degree of variation in flea detection when swabbing PD burrows. Many factors influence flea collection with the swabbing technique, including burrow activity (Poje et al. 2020), ambient temperature, sunlight, size of swab, and depth of swab insertion (Seery et al. 2003, Eads 2017). Fur fleas might commonly be missed by swabs, unless few or no live PDs are available, which motivates fleas to quest for hosts. We found little relationship between burrow swabbing characterizing actual PD flea parasitism rates and dynamics. Thus, if investigators are interested in the abundance of fleas on PDs, we recommend combing PDs.
Implications for plague ecology and management
Rates of plague transmission fall along a continuum (Ramakrishnan 2017, Biggins and Eads 2019). In our study of WTPDs, during a period of enzootic plague (Biggins et al. 2010), 67% of O. hirsuta pools from WTPDs tested positive for Y. pestis. Yet Y. pestis was not detected in O. hirsuta from burrows (similar results were found for P. simulans). If swabbing were the only technique used, it would have seemed as if Y. pestis was absent, when in fact it was present and infectious, as demonstrated by mouse inoculation and suggested by improved WTPD survival on colonies where deltamethrin was used for flea control (Biggins et al. 2010).
In many, but not all cases, Y. pestis-positive fleas are rarely swabbed from burrows or combed from PDs or other mammals during enzootic phases of plague circulation (Holmes et al. 2006, Biggins et al. 2010, Matchett et al. 2010, Kosoy et al. 2017, Russell et al. 2018, but see Hanson et al. 2007, Griffin et al. 2010, St. Romain et al. 2013). At least four nonexclusive explanations might explain why. First, diagnostic tests might provide false-negatives with fleas collected during enzootic periods because populations of Y. pestis are reduced (compared to populations during epizootic “recharge” events; Baltazard 1964). Detection rates might increase if fleas with visible blood meals are available (Stevenson et al. 2003) and large numbers of fleas or large pools are tested (Griffin et al. 2010, St. Romain et al. 2013). It should be remembered, if a large flea pool tests positive, few of the fleas may have actually harbored Y. pestis (Engelthaler and Gage 2000), and those fleas might not necessarily be infectious.
Second, during enzootic periods, Y. pestis is presumably sequestered in space and time (but still causes host mortality at local scales; Matchett et al. 2010, Biggins et al. 2021). Large numbers of swabbing events across PD colonies might be needed to gain sufficient numbers of fleas, and in particular to detect Y. pestis-positive fleas (but see Hanson et al. 2007), which might tend to occur in patches of habitat where plague transmission is active (Kosoy et al. 2017, Biggins and Eads 2019). During enzootic periods in Colorado, Griffin et al. (2010) completed 2498 swabbing events on WTPD colonies. In total, 3083 fleas were collected; only 50 of 1071 flea pools tested positive for Y. pestis. All the positive flea pools came from a single site where the swabbing effort was intensive (1816 swabbing events, compared to a range of 130–302 events at sites where Y. pestis was not detected).
Third, even if surveyors swab burrows or comb hosts harboring infected fleas, the timing of sampling is critical. Some flea species spend most of their time on hosts, and some species might inhabit the same host throughout most (or all) of their lives (Krasnov et al. 2004). If a host dies from plague in a burrow inhabited by other live hosts (likely for colonial PDs during enzootic periods), fleas on a perished host might simply shift from the carcass to a live host. Presumably, surveyors rarely swab burrows or comb hosts during such a short window of opportunity for optimal collection.
Fourth, and related, if live PDs are abundant enough to harbor most of the local population of mature adult fleas, or most mature adult fleas have recently acquired bloodmeals (e.g., because they feed often), surveyors might tend to collect adult fleas that recently emerged from pupal cocoons and are questing for a host. Surveyors may collect few to no infectious adult fleas that recently fed on bacteremic hosts. Upon emergence from cocoons, newly emerged adult fleas would (presumably) not harbor Y. pestis; the alimentary canal of larvae is nearly autolysed and replaced during pupation, and the adult digestive tract is mostly built anew (although we caution that some intact segments of organs such as the intestine might be retained through metamorphosis, functioning as a starting site for growth of adult organs; Linser and Dinglasan 2014).
If infected fleas tend to remain on live PDs during enzootic periods and, perhaps, the initial stages of plague epizootics, scientists may commonly fail to collect Y. pestis-positive fleas on swabs, but succeed, although rarely (Russell et al. 2018), in collecting Y. pestis-positive fleas from PDs. Biologists might collect Y. pestis-positive fleas from burrows during epizootics (Kartman et al. 1962, Lechleitner et al. 1968) and from PD carcasses found opportunistically if the carcasses are found before many or all fleas abandon the carcass (Engelthaler and Gage 2000). By that time, however, the utility of plague surveillance is greatly diminished as the window of opportunity for effective mitigation has narrowed or closed.
Biologists often fail to collect Y. pestis-positive fleas during epizootics on PD colonies, even on colonies with recent, near-total population collapses (e.g., Thiagarajan et al. 2008). Factors noted above, and the lethal effects of Y. pestis on fleas themselves (Y. pestis can “block” blood-feeding; Lorange et al. 2005, Eisen et al. 2009), may lead surveyors to collect noninfected fleas during most of the plague cycle (see Jennelle et al. 2007 for a related example). Sometimes fleas (infected or not) are simply hard to find in burrows regardless of building or subsiding rates of plague circulation (e.g., as suggested herein with the CMR data). Proliferating populations of Y. pestis may kill large numbers of fleas. After many or all surrounding PDs have succumbed to plague, noninfected fleas may quickly perish with reduced or no access to blood (Wilder 2007). If at least some PDs and fleas survive an epizootic, as was presumed to have happened at CMR (e.g., Table 2), or PDs and fleas recolonize extirpated sites (Antolin et al. 2006) and plague transmission subsides to an enzootic level, flea populations can rebound. If all BTPDs die and no fleas remain, BTPDs may later recolonize the site, and fleas could arrive on those BTPDs and found a new flea population (Eads 2014).
Conclusions
We documented important limitations of flea data, inferences, and plague detection from burrow swabbing on PD colonies. Detection of Y. pestis-positive fleas (or rodent carcasses) should not be the sole criterion for determining if plague is circulating on PD colonies; the absence of evidence is not evidence of absence. If massive numbers of PDs have disappeared, even if Y. pestis is not detected in fleas, hosts, soil, or other venues of Y. pestis persistence, plague is the likely culprit. No other disease or biological factor is known to cause such large mortality events among PDs, including tularemia (caused by Francisella tularensis; Barnes 1993, Cheery et al. 2019; although very small PD colonies may dwindle due to recreational shooting, or during droughts; for an example of the later, see Stephens et al. 2018). That said, we caution that burrow swabbing for fleas on PD colonies might be useful in some cases, especially during controlled experiments (Biggins and Eads 2019). Swabbing has been used to evaluate host-seeking by fleas at local scales (Bron 2017, Biggins and Eads 2019), to collect nest fleas, and fleas that occur in PD burrows from time to time, but rarely parasitize PDs (Ubico et al. 1988, Cully et al. 1997, Bron 2017), and to collect fleas for laboratory studies of plague transmission (Wilder et al. 2008).
Footnotes
Acknowledgments
We are indebted to CDC, who identified flea species from WTPD colonies and tested flea pools for Y. pestis. We thank many technicians for their hard work, and the Bureau of Land Management (Vernal, UT) and CMR staff for logistical support. We thank D. Long and Turner Endangered Species Fund for facilitating the research at Vermejo Park Ranch. We thank M. Antolin, K. Gage, J. Montenieri, T. Rocke, J. Wimsatt, and many other colleagues for discussions on flea and plague ecology. We thank J. Hughes and three anonymous reviewers for constructive comments on the article. Data are available in Eads et al. (![]() ). Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
). Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
Funding and logistical support were provided by the U.S. Geological Survey; Turner Endangered Species Fund; Turner Enterprises Incorporated; Colorado State University; Centers for Disease Control and Prevention (CDC); U.S. Fish and Wildlife Service; Charles M. Russell (CMR) National Wildlife Refuge; Shortgrass Steppe Long-Term Ecological Research Project (DEB 021763 and 0823405); National Science Foundation under grant no. 1027319; and the U.S. Department of Defense Strategic Environment Research and Development Program Number 16 RC01-012.