
Abstract
Introduction:
Many SARS-CoV-2 variants of concern (VOC) have been reported recently that were linked to increased transmission. In our earlier study using VOC 202012/01 (U.K. variant) and D614G variant in the hamster model, we observed higher viral RNA shedding through nasal wash in the case of U.K. variant with lower pathogenicity in lung. In this study, we have studied transmission of these two variants by direct contact, aerosol, and fomite routes in Syrian hamsters and compared the viral load and body weight changes in hamsters exposed by both variants to understand the transmission efficiency.
Methods:
Nasal, throat, and rectal swabs were collected sequentially to assess viral load till 14 days.
Results:
Transmission could be established by direct, aerosol, and fomite contact in Syrian hamsters. Body weight loss or viral load in the contact animals exposed did not show any statistical significance.
Conclusion:
The study demonstrated comparable transmission of both U.K. and D614G variants of SARS-CoV-2 in Syrian hamsters in the given conditions. Provided these data, it seems that all the routes of exposure are effective leading to higher transmission.
SARS-CoV-2 has accumulated numerous genetic changes on circulation all over the world. Some of these mutations have the potential to change the virus characteristics such as infectiousness, transmissibility, and severity of disease and can have impact on diagnostics, vaccines, and therapeutics (WHO 2021). SARS-CoV-2 variant with a D614G substitution that emerged in the first quarter of the year 2020 supplanted the initial SARS-CoV-2 strain, showing its increased fitness to become the dominant strain circulating globally.
Recently, many variants of concern (VOC) were reported from the United Kingdom, South Africa, Brazil etc., which were linked to increased transmission, disease severity, and vaccine escape mutants (Davies et al. 2020). To categorize a variant as VOC, various risk elements such as increased transmissibility, morbidity, mortality, and immunity escape factors needs to be studied. Scanty information is available on these aspects about the variants.
In our earlier study on virus shedding using VOC 202012/01(U.K. variant) and D614G variant in Syrian hamster model, we observed significantly higher viral RNA shedding through nasal wash in case of the U.K. variant (Mohandas et al. 2021a). Direct contact, aerosol, and fomite routes of transmission of SARS-CoV-2 have been established in hamster model (Sia et al. 2020). Here we have compared the transmission of both the U.K. and D614G variants by various routes in Syrian hamster model to understand whether the high viral RNA shedding through nasal cavity in hamsters infected with the U.K. variant could enhance the transmission efficiency of the variant.
The study was approved by the institutional animal ethics (No. NIV/IAEC/2021/MCL/01) and biosafety committee (No. NIVIBSC/05.01.2021/02) of Indian Council of Medical Research (ICMR)-National Institute of Virology (NIV), Pune, and all the experiments were performed as per the institute ethical guidelines (CPCSCEA guidelines, Government of India).
The study was performed in the containment facility with a total of 36 male hamsters of 6–8-week age housed in individually ventilated cages. Nine hamsters each were intranasally infected with two SARS-CoV-2 variants, that is, U.K. variant (hCoV-19/India/NIVP1 20203522/2020, GISAID identifier: EPL_ISL_825088) and a SARS CoV-2 variant isolated during March 2020 from Italian tourists that possess D614G mutation (hCoV-19/India/2020770/2020, GISAID identifier: EPL_ISL_420546), with 0.1 mL of virus of 105.5 TCID50/mL under isoflurane anesthesia and were used as donor hamsters for studying transmission through direct contact, aerosol, and fomite routes (Sarkale et al. 2020, Yadav et al. 2021). All the experiments were performed in triplicates for both the variants.
SARS-CoV-2 genomic RNA (gRNA) load was tested in the nasal wash, throat, and fecal swab samples collected from the contact hamsters on every alternate day till 14 days postexposure (DPE) using E gene quantitative real-time RT-PCR as described earlier (Choudhary et al. 2020). Virus titration was also performed for the nasal wash samples in Vero CCL81 cells.
The donor hamsters infected with the U.K. variant showed progressive weight loss with the maximum average weight loss of 11% ± 2.82% (mean ± standard deviation) on day 6, whereas the donor hamsters infected with D614G variant showed a maximum average weight loss of −6.66% ± 1.36% on day 8. The donor hamsters showed regain of weight thereafter. All the donor hamsters, bedding, cage surfaces, and water bottle nozzle samples from cages used for fomite transmission study were tested for gRNA to ensure the presence of the virus before exposure (Supplementary Table S1).
Twenty-four hours postinfection, three donor hamsters were cohoused with a naive hamster (referred to as contact hamster further) in 1:1 ratio in a new cage to study direct transmission and were observed till 14 DPE for body weight change and any respiratory signs. The contact hamsters exposed with the U.K. variant showed maximum average weight loss of −2.93% ± 0.34% and with D614G variant showed −4.8% ± 3.13% on 8 DPE (Fig. 1A). The contact hamsters exposed with both variants showed viral gRNA positivity in the throat swab, nasal wash, and fecal samples from 2 DPE and peak average viral gRNA load by 2 to 4 DPE (Fig. 1B–D). This is similar to the pattern of detection reported in intranasal inoculated hamsters with both variants (Hou et al. 2020, Mohandas et al. 2021). Titration of nasal wash samples showed consistent presence of virus till 10 DPE with comparable titer in hamsters exposed with both variants (Fig. 1E).
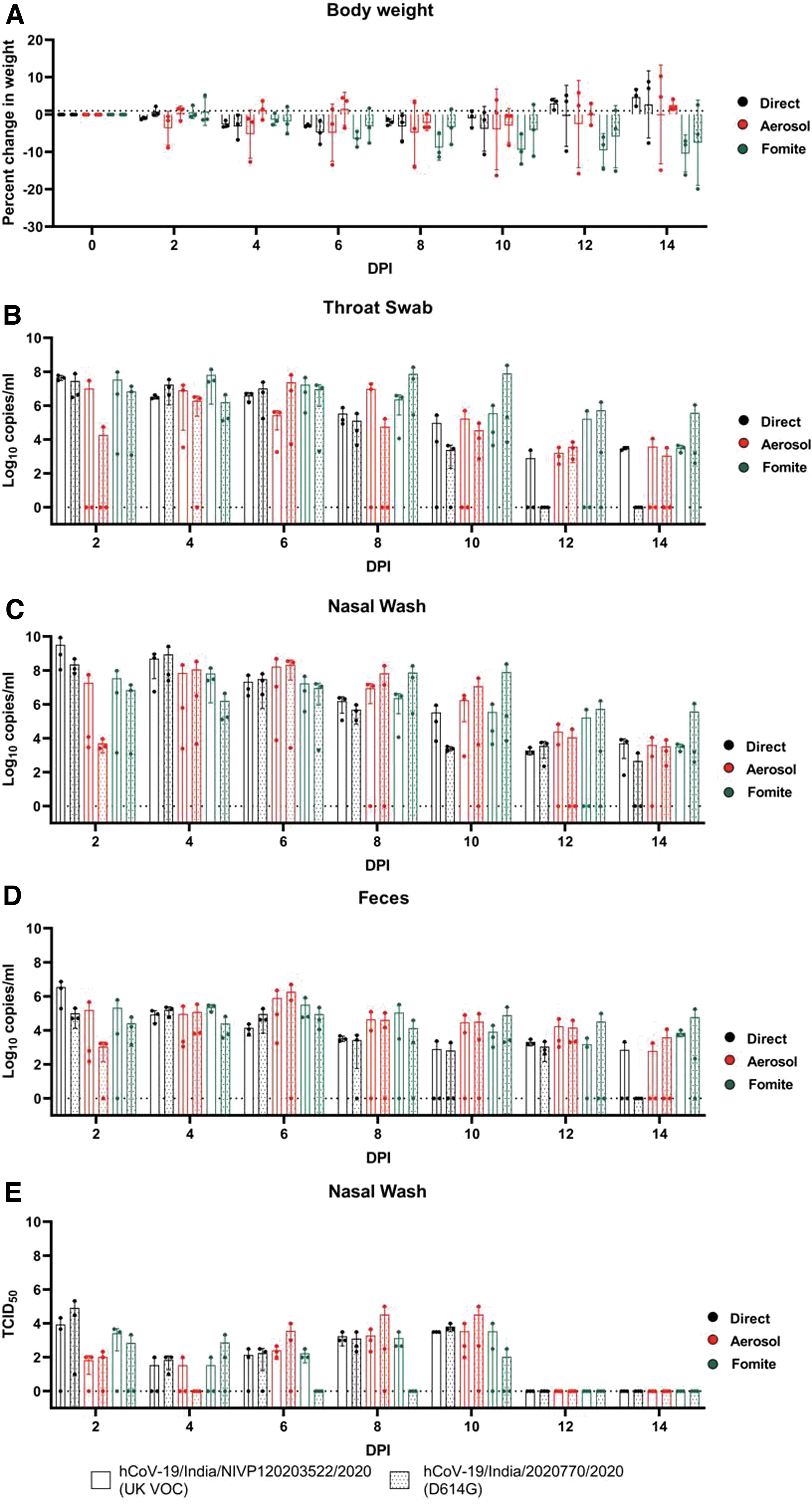
Percentage body weight change and SARS-CoV-2 viral load in the contact hamsters postexposure.
As the direct contact transmission could be contributed by aerosol and fomites, we assessed these routes of transmission alone. To assess the aerosol transmissibility, SARS-CoV-2–infected hamster after 24 h postinfection was cohoused in a modified individual ventilated cage with partition (which allows airflow) with a naive hamster for 8 h. The contact hamsters were housed in new cages and were observed for 14 days. One hamster from the aerosol contact group of D614G variant did not show any body weight loss and showed very low or negligible amount of viral RNA load in the samples. The other hamsters of the aerosol contact group of the U.K. variant and D614G variant showed a peak average weight loss of −8.9% on 8 DPE and −4.2% on 10 DPE, respectively (Fig. 1A). The viral gRNA detection was observed from 2 DPE and the peak average viral gRNA detection in the nasal wash and fecal samples was observed on 6 DPE for hamsters exposed with both variants and higher viral load till 10 days (Fig. 1B–D). The TCID50/mL of the nasal wash samples from 6 to 10 DPE showed higher titers (Fig. 1E).
For the fomite transmission study, three naive hamsters were housed in different cages with soiled bedding of SARS-CoV-2 infected hamsters housed for 48 h after infection. Progressive body weight loss was also observed in fomite contact hamsters of both the variants till 14 days (Fig. 1A). The peak average viral gRNA in fomite contact hamsters varied from 4 to 8 DPE in case of the U.K. variant and 8 to 10 DPE with the D614G variant (Fig. 1B–D). This is contrary to an earlier study that reported less efficient fomite transmission by SARS-CoV-2 in hamsters (Sia et al. 2020). Even though viral gRNA could be detected till 14 DPE, live virus particles could not be detected from 6 DPE in case of the D614G variant contacts (except in one animal on 10 DPE) in contrast to the U.K. variant contacts where it could be detected till 10 DPE (Fig. 1E).
Anti-SARS-CoV-2 IgG antibodies by anti-SARS-CoV-2 IgG ELISA as described earlier by Mohandas et al. (2021b) could be detected in all the donor and contact animals on 21 DPE except one contact hamster of the D614G variant contact of the aerosol group that showed negligible viral gRNA load. The day-wise comparison of the body weight loss and the viral shedding pattern in contact hamsters by both variants on Mann–Whitney test did not show any statistical significance. Also the comparison of different routes of transmission by each variant on Kruskal–Wallis test did not show any statistical significance.
The virus detection in contact hamsters was seen as early as on 2 DPE for both variants, indicating efficient transmission and a maximum infectious period of 10 days by all routes. The peak viral gRNA levels were comparable among different transmission routes, whereas the peaks of detection by the aerosol and fomite route were found extended and with more variations among contact animals. The percentage body weight loss also varied among different routes of transmission. This could be due to difference in the amount of virus dose exposure by these routes. Earlier research have shown that lower and higher doses of virus inoculums show comparable viral gRNA loads in hamsters, but the lung lesions and body weight loss vary with virus dose (Mohandas et al. 2020, Ryan et al. 2021).
The limitation of the study is that the lung viral load or pathology induced in the contact hamsters were not estimated. Also transmission studies with shorter virus exposure times of contact hamsters should be explored.
Conclusion
The transmission of SARS-CoV-2 variants could be established in Syrian hamsters by direct, aerosol, and fomite routes as evident by the body weight loss, detection of viral gRNA/live virus in the samples, and anti-SARS-CoV-2 IgG antibodies in the contact hamsters. The study demonstrated comparable transmission of both the U.K. variant and D614G variant of SARS-CoV-2 in Syrian hamsters.
Footnotes
Authors' Contributions
P.D.Y. and S.M. conceived, designed, and performed the animal experiments. P.D.Y., A.S., K.L., and S.H. performed the laboratory investigations and standardizations. D.N. performed the statistical analysis. P.S. performed the titration experiments. S.M. wrote the draft of the article. P.D.Y., S.K., and K.L. reviewed and substantively revised the article. All the authors have read the article and agree to its contents.
Acknowledgments
The authors acknowledge the support received from Prof. (Dr.) Priya Abraham, Director, ICMR-NIV, Pune, and the laboratory team of Maximum Containment Facility, ICMR-NIV, Pune, which includes Mr. Manoj Kadam, Mr. Annasaheb Suryawanshi, Mr. Abhimanyu Kumar, Dr. Rajlaxmi Jain, Mr. Rajen Lakra, Mr. Shreekant Baradkar, Mrs. Ashwini Waghmare, Ms. Manisha Dudhmal. Mr. Kundan Wakchuare, Ms. Tejashri Kore, Ms. Shilpa Ray, Ms. Priyanka Waghmare, and Ms. Poonam Bodke. The authors also thank Dr. SSYH Qadri, Head, Animal Facility, and Dr. M. Satya Vani, ICMR-National Institute of Nutrition, Animal Facility, for their support.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
The authors acknowledge the funding received from Indian Council of Medical Research as intramural COVID-19 research funding under the project “Propagation of SARS-CoV-2 variant isolate and characterization in cell culture and animal model.”
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.