
Abstract
Rodents are known reservoir hosts for a number of pathogens that can spillover into humans and cause disease. These threats are likely to be elevated in informal urban settlements (i.e., slums), where rodent and human densities are often high, rodents live in close proximity to humans, and human knowledge of disease risks and access to health care is often limited. While recent research attention has focused on zoonotic risks posed by urban rodents in major cities around the world, informal urban settlements have received far less attention. Here we report on a study in which samples were collected from 195 commensal rodents and 124 febrile human patients in the Kibera informal settlement in Nairobi, Kenya (one of the largest informal urban settlements in the world). Using immunofluorescence assays, samples were screened for antibodies against common rodent-borne zoonotic virus groups, namely orthopoxviruses, arenaviruses, and hantaviruses. We detected antibodies against orthopoxviruses in rodents (4.1% positive) and antibodies in humans against orthopoxviruses, arenaviruses, and hantaviruses (4.8%, 3.2%, and 8.1% positive, respectively). No rodents had antibodies against arenaviruses or hantaviruses. These results provide strong evidence for the circulation of zoonotic viruses in rodents and humans in Kibera urban settlement, but discordance between viruses detected in host groups indicates that other species or taxa may also serve as reservoirs for these zoonotic viruses or that humans testing positive could have been exposed outside of the Kibera settlement. More broadly, this study highlights the threat posed by zoonotic viruses in informal urban settlements and the need to mitigate human exposure risks.
Introduction
Rodents are known reservoir hosts for a number of zoonotic viruses that can have severe consequences for human health. Examples include orthopoxviruses, arenaviruses, and hantaviruses, which are all globally widespread and cause considerable morbidity and mortality in humans (Charrel et al. 2011, Moss 2013). A unique feature of some rodent species, compared with many other wildlife taxa, is their often-close associations with humans, which can create regular opportunities for human exposure to pathogens they carry. Exposure can occur directly through bites and other close contacts, or indirectly via contamination of food, water, and through virus aerosols shed into the environment in rodent urine and feces (Meerburg et al. 2009). Furthermore, there is increased tendency for exotic pet rodents, which are sometimes also able to spread zoonoses (CDC 2003).
Commensal rodents are among the best adapted mammalian species to urban areas (Battersby et al. 2008). Their rapid reproductive cycles and fast maturation rates allow them to capitalize on favorable environments. Moreover, urban environments often have readily available shelter and food sources for rodents (Colvin and Jackson 1999, Cevidanes et al. 2017), enabling them to maintain high abundances and quickly increase following population disturbances such as human control efforts (Lambropoulos et al. 1999, Fernández et al. 2007). These high-density reservoir host populations can also result in greater diversity and prevalence of pathogens, through mechanisms such as density-dependent transmission and density thresholds for pathogen persistence (Kosoy and Bai 2019), enhancing pathogen exposure and subsequent disease risks for humans.
The number and extent of urban slum settlements have increased dramatically over the past several decades, predominantly in low- and middle-income tropical regions (UN-HABITAT 2015), creating favorable environments for urban rodents. While several studies have investigated urban rodents and their zoonotic pathogens in major cities such as New York (Firth et al. 2014, Williams et al. 2018), informal urban settlements have received less attention despite their often-greater abundance of shelter and food sources for rodents. Moreover, public health knowledge of inhabitants and access to health facilities are often lacking in informal settlements (Ezeh et al. 2017, Lilford et al. 2017), creating ideal conditions for the initiation of zoonotic disease outbreaks that can then spread to other areas.
To address these deficiencies, here we report on a study to investigate zoonotic viruses associated with commensal rodents in the Kibera informal urban settlement in Nairobi, Kenya; one of the largest urban informal settlements in the world (Glass et al. 1989, Childs et al. 1991). While Kibera settlement is known to harbor large populations of rodents (Halliday et al. 2013), information on the viruses they carry and whether they spillover into humans is lacking. We screened both rodents and febrile humans inhabiting Kibera settlement for antibodies against selected major zoonotic virus groups carried by rodents, namely: orthopoxviruses, arenaviruses, and hantaviruses.
Methods
Study site
Kibera informal settlement is located in Nairobi, Kenya, about 5 km south of Nairobi city center (1° 17′ 11.0004″ S, 36° 49′ 2.0028″ E). It has an estimated total population of 283,024 people, spread over 2.5 km2 of land (Ren et al. 2020). The majority of residents have poor or unreliable income, and either provide unskilled labor at the nearby industrial area of Nairobi or are self-employed with small businesses such as food or grocery shops, hair salons, repair shops, and used goods shops. Most human dwellings in Kibera are basic, measuring about 12′ × 12′ each and built with mud walls, corrugated tin roof, and have concrete or dirt floors. Each dwelling houses an average of three inhabitants (APHRC 2014). There is no functional sewerage system in Kibera settlement, with one latrine (hole in the ground) shared among ∼50 dwellings.
In Kibera informal settlement, undifferentiated febrile illnesses are common with an average of 2.7 cases per person/year for children <5 years and 0.58 cases per person/year for people older than 5 years of age (Feikin et al. 2011). Despite this, the causative agents are in general not known and nothing is known about rodent-borne viruses and their prevalence in Kibera settlement.
Rodent trapping and sample collections
Rodent trapping was conducted for 10 consecutive days from February 3 to 13, 2019. Kibera settlement was divided into five areas on the basis of the preexisting administrative zones to ensure geographic spread in trapping sites (Fig. 1).
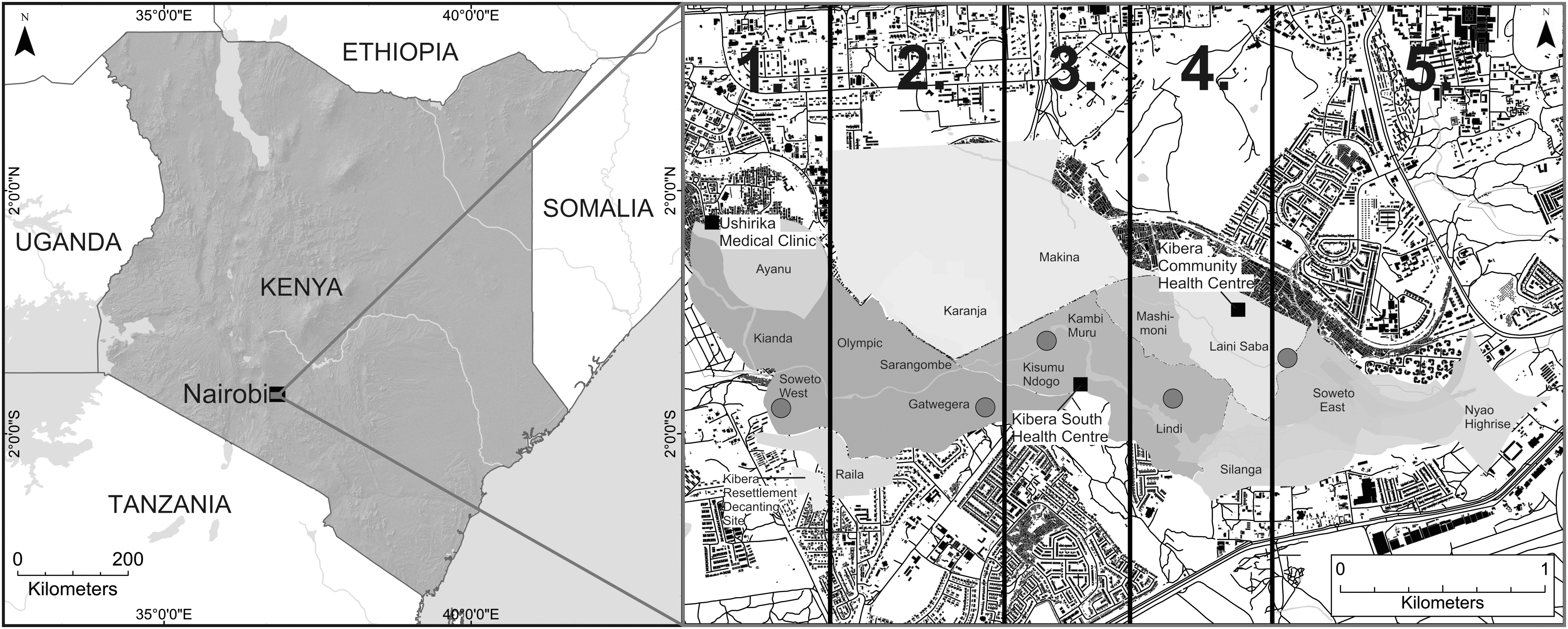
Map of Kenya with the Kibera settlement and its preexisting administrative boundaries (1–5) used to define the five study zones. Red circular points indicate rodent trapping areas in each zone, and black squares indicate the three health care centers that were used for the collection of blood samples from febrile patients. For this map, a 30-meter DEM 2015 from SRTM was obtained from the Regional Centre for Mapping of Resource for Development, and Nairobi land use data (2010) were acquired from Columbia University's Center for Sustainable Urban Development. DEM, Digital Elevation Model; STRM, Shuttle Radar Topography Mission.
A total of 300 trap nights were conducted across the 5 focus areas (60 trap nights/area). Traps were distributed to households around a central point in each area (∼3 traps per household). During each trapping night, Sherman live traps (H.B. Sherman Traps, Inc., Tallahassee. FL) baited with small, dried fish were set indoors on the floor against walls and under furniture. Traps were set late in the afternoon and checked the following morning. Additional traps were set in places frequented by inhabitants and known to have large rodent populations, such as a communal dump site and dry sewers. In these sites, traps were set at dusk and periodically checked until midnight by researchers and then removed to prevent theft.
Early the following morning, traps with live rodents were transported to the University of Nairobi laboratory for processing. Trapped rodents were anesthetized using isoflurane gas and then euthanized via cervical dislocation. Biometric data were recorded and were used for taxonomic identification. They included body weight, sex, reproductive status, body length, and tail length; rodents were immediately dissected for the collection of terminal samples. Heart samples were stored in phosphate-buffered saline (PBS) in preparation for serology assays and frozen at −20°C. They were later shipped on dry ice to the University of Helsinki in Finland for diagnostic assays.
Human sample collections
Human serum samples were collected from febrile patients seeking medical care at three local health facilities located within Kibera settlement from February to June 2017. These facilities predominately serve residents living in the Kibera settlement. Any patient with a temperature of 37.5°C or higher was eligible for inclusion in the study. Adult participants provided informed written consent, while adult guardians provided written informed consent for anyone aged <18 years.
Patient's demographic information, including sex and age, was captured with age categorized into two groups; younger than 18 years (children) and older than 18 years (adults). Consenting patients were sent to a nearby hospital laboratory where blood was collected by technicians into 5 mL serum tubes using Vacutainer butterfly needles 21G and 23G and a butterfly needle adaptor. Blood samples were allowed to clot and then centrifuged at 1000 × gravitational units (g) for 10 min to separate serum. Serum samples were then transferred aseptically to sterile vials labeled with the patient's identifiers and stored at −80°C at the University of Nairobi before being shipped on dry ice to the University of Helsinki for diagnostic assays. Only one sample was collected per patient.
Laboratory diagnostics
Samples were screened for antibodies against orthopoxviruses, arenaviruses, and hantaviruses using immunofluorescence assays, as previously described (Kallio-Kokko et al. 2006, Kinnunen et al. 2011, Forbes et al. 2014). Briefly, the process involved thawing the human serum and rodent heart samples, diluting with PBS at a ratio of 1:20, and incubating with specific antigens, followed by several wash steps. Slides were dried and goat anti-human IgG and goat anti-mouse IgG conjugates were added to human and rodent samples, respectively. Slides were then incubated and washed again, and lastly, examined under a fluorescence microscope.
These assays are not virus-specific and cross-react with other closely related viruses. This is especially useful when we do not know which particular orthopoxvirus, arenavirus, or hantavirus, we are searching for in samples. Puumala orthohantavirus (PUUV) and Dobrava-Belgrade orthohantavirus (DOBV) represent different hantavirus serogroups, and by using assays for both of them, we can detect all possible rodent-borne hantaviruses. For arenaviruses, we used the lymphocytic choriomeningitis virus (LCMV) that cross-reacts with all Old-World arenaviruses, and so, we are not only detecting LCMV but any Old-World arenavirus as well. Similarly, cowpox virus is highly cross-reactive and was used to detect orthopoxviruses.
Results
A total of 195 rodents were trapped and sampled, comprising 149 Mus musculus and 46 Rattus rattus. Although Rattus norvegicus is also known to be present in Kibera settlement, none was captured. Eight rodent blood samples were positive for anti-orthopoxvirus IgG antibodies; three M. musculus and five R. rattus representing seropositivity of 2.0% and 10.9%, respectively (Table 1). All rodents were seronegative for arenaviruses and hantaviruses.
Proportion of IgG Seropositive Rodents Captured in Kibera Informal Settlement in February 2019
Total positive/total screened (percentage positive).
The overall rodent seroprevalence for orthopoxviruses in Kibera urban settlement was 4.1% and varied among sites. Of the five focus areas, a communal dump in Lindi (zone 4) had the highest seroprevalence with 33.3% (3/9) positive, followed by households in the same area of 5.1% (2/39) (Table 2). No rodents with orthopoxvirus antibodies were detected in the two trapping zones (zone 2 and 3), despite relatively high sample numbers.
Proportion of Seropositive Rodent Samples for Orthopoxviruses Across the Different Trapping Sites and Rodent Hosts in Kibera Informal Settlement
Total positive/total screened (percentage positive).
A total of 124 serum samples were collected from febrile patients and screened for the focus virus groups. Of these, 4 (3.2%) were seropositive for arenaviruses, 5 (4.8%) were seropositive for orthopoxviruses, and 10 (8.1%) were seropositive for hantaviruses (Table 3). Of hantaviruses, 4 (3.2%) were seropositive for DOBV-like viruses and 6 (4.8%) were seropositive for PUUV-like viruses. There was only one patient (male child) who was seropositive for more than one virus (LCMV and PUUV).
Seroprevalence of Rodent-Borne Zoonoses in Patients Attending Health Care Facilities in Kibera Informal Settlement from February to June 2017
DOBV, Dobrava-Belgrade orthohantavirus; PUUV, Puumala orthohantavirus.
Seropositivity distribution across the health facilities varied; the Kibera Community Health Centre and the Ushirika Medical Clinic had relatively high orthopoxvirus and PUUV-like virus seroprevalence, while the Ushirika Medical Clinic and Kibera South Health Centre reported relatively high seroprevalence for DOBV-like virus (5.9%) and arenavirus (10.0%), respectively. A breakdown of positive results by patient sex and age group is available in Table 4.
Seroprevalence of Rodent-Borne Zoonoses Among Patients Attending Health Care Facilities in Kibera Informal Settlement Segregated by Age and Sex
Discussion
Little is known about zoonotic viruses carried by rodents in urban slums in Africa and in other informal settlements across the world, despite high densities and close associations between humans and rodents in these settings and their potential to seed disease outbreaks that spread to other areas. Here we used serological assays to investigate evidence of common rodent-borne zoonotic virus groups in humans and rodents. We report the presence of antibodies against all three virus groups—orthopoxviruses, arenaviruses, and hantaviruses—and also note mismatches between their presence in rodents and humans. These results highlight the risk that rodent-borne zoonotic viruses pose in informal urban settlements and the need for efforts to mitigate human exposure risks.
Our results show evidence of circulation of orthopoxviruses in both rodents and humans in Kibera urban settlement. The overall seroprevalence for orthopoxviruses in captured rodents and human patients was similar. However, clear differences were observed in rodent seroprevalence among sites, ranging from all negative to >30% positive (although sample numbers were sometimes low). These differences could be due to the trapping environment, with the only communal dump site trapped in Lindi (trapping zone 4) having particularly high prevalence. Both Lindi and Soweto West (zone 1) trapping sites, which had high seroprevalence of orthopoxviruses in rodents, also recorded high seroprevalence in human serum samples in the Kibera Community Health Centre and Ushirika Medical Clinic.
The difference in site seroprevalence is similar to studies by Lederman et al. (2007) in the Republic of Congo, where variation among people living in different villages was also observed. The seroprevalence in this study is roughly consistent with a study in the Democratic Republic of Congo, which reported 2% anti-orthopoxvirus antibodies in mammals in rural settings (Doty et al. 2017). Similarly, low human seroprevalence of orthopoxviruses has also been reported in a rural population serosurvey in Sierra Leone (1.3%) (MacNeil et al. 2011).
We detected antibodies against hantaviruses and arenaviruses in humans, but not in rodents. Our rodent trapping focused on urban settings where M. musculus and R. rattus dominate. However, it is possible that other rodent species not encountered in our study, but carry these viruses, are present at lower densities or in specific areas within the informal settlement. Another possible explanation is that residents of Kibera were exposed to these viruses outside of Kibera settlement.
Examples of such exposure opportunities include Tigray virus (TIGV), a hantavirus previously reported in East Africa from Ethiopian white-footed mouse (Meheretu et al. 2012), two novel hantaviruses, Kilimanjaro virus (KMJV) and Uluguru virus (ULUV), that were detected in Tanzanian shrews (Kang et al. 2014), and a sister lineage of Sangassou virus that was detected from the African wood mouse in Kenya (Těšíková et al. 2017). Similarly, a survey of rodent-borne viruses in 2016 reported a novel arenavirus in Grammomys macmillan rodents that were trapped in Kitale, western Kenya (Onyuok et al. 2019). Many Kibera residents originate from the western parts of Kenya (Amis 1984) and therefore might have been exposed to viruses when they travel to visit family members and friends in the rural villages.
The observed reaction of human serum samples to PUUV hantavirus and DOBV-Belgrade hantavirus is an indication of exposure to multiple different African hantaviruses since no samples positive to one hantavirus type also reacted to the other. Although our study reports seroprevalence of hantaviruses in urban slums, the seroprevalence is within the range of those previously reported in human population-based surveys in close proximity to national parks in both Côte d'Ivoire (3.9%) and the Democratic Republic of Congo (2.4%) (Witkowski et al. 2015). In contrast, our seroprevalence was higher than levels reported in Guinea (1.2%) and South Africa (1.0%) (Klempa et al. 2010, Witkowski et al. 2014).
Although sample numbers were small, our study also identified demographic-based trends for hantavirus disease epidemiology that could suggest disparity in infection risk; consistent with studies in the Netherlands and Brazil (Sane et al. 2014, Vieira et al. 2016), we identified more infected females than males, which could be linked to occupation (Vieira et al. 2016). Clearly, more research is needed to provide stronger insights into the role of gender on infection risk in Kibera informal settlement and other urban slums.
The serological evidence of orthopoxviruses in rodents and orthopoxviruses, arenaviruses, and hantaviruses in humans, but no reported diseases in the study population, is an indication that zoonotic rodent-borne human illnesses may be occurring but are not reported or detected. For example, it is suggested that hantavirus infections may not be diagnosed in sub-Saharan Africa due to weak surveillance, lack of laboratory capacity, lack of disease awareness among clinicians and because symptoms of hantavirus might overlap with those of other common febrile illnesses (Klempa et al. 2010, Witkowski et al. 2014). All these factors worsen the burden of undifferentiated febrile illnesses in these settings (Feikin et al. 2011) leading to underestimation of rodent-borne pathogens. Previous studies have also documented the presence of Leptospira and Bartonella species among rodents in Kibera informal settlement (Halliday et al. 2013), increasing the zoonotic risks among the residents.
Conclusion
Although the serological assay results reported here do not identify the specific viral species due to broad cross-reactivity, this study provides strong evidence for infection by orthopoxviruses, arenaviruses, and hantaviruses in humans and in commensal rodents in one of the largest urban informal settlements in the world. This research highlights the risks rodent-borne viruses pose to humans in urban slum settlements in low- and middle-income countries, and more generally, the potential for informal urban settlements as a source of zoonotic disease outbreaks that can spread to other areas.
Ethics Statement
The research was conducted under permits from Kenyatta National Hospital-University of Nairobi Ethics and Research Committee (permit No. P707/11/2015), Biosafety, Animal Use and Ethics Committee-University of Nairobi (permit No. FVM BAUEC/2018/180), and the National Commission for Science, Technology and Innovation (permit No. NACOSTI/P/18/76501/22243). This study was approved by the University of Arkansas IACUC committee.
Footnotes
Acknowledgments
We thank Joseph Ondieki and the village elders in Kibera for their participation as guides during the rodent field sampling in Kibera. We also thank the staff of Ushirika Medical Centre, Amref Kibera Community Health Centre, and Kibera South Health Centre. We are also indebted to all the people of Kibera informal settlement who participated in the research. We also thank the technical staff at the Department of Veterinary Pathology, Microbiology and Parasitology, University of Nairobi, Kenya, for assistance during sample taking from the rodents and the Chairman of the same department for allowing use of the laboratory facilities for this work.
Authors' Contributions
J.G.O., K.M.F., T.S., O.V., M.M., and P.N. designed the human and rodent sampling. J.G.O., K.M.F., T.S., M.M., O.A., and P.N. performed fieldwork. R.U. prepared the map of the study site. J.G.O., H.A., and E.K. performed the serological analysis. J.G.O. drafted the article. All authors contributed to the revisions and approved the final version.
Author Disclosure Statement
The authors declare no conflict of interests.
Funding Information
Funding for this study was provided by the Finnish Cultural Foundation, the Jenny and Antti Wihuri Foundation, the Academy of Finland (grant No. 318726), Helsinki University Hospital Funds, the Jane and Aatos Erkko Foundation, and the Arkansas Biosciences Institute, and NSF grant DEB 1911925.