
Abstract
Babesiosis is a tick-borne disease caused by intraerythrocytic protozoa belonging to the genus Babesia that is primarily transmitted through Ixodid ticks and is highly pathogenic to ruminants, horses, pigs, dogs, cats, and in some cases, even humans. Babesia is considered an increasing threat to animal and human health. In this study we report Babesia capreoli infection in Korean water deer (Hydropotes inermis argyropus). Blood samples of 77 wild animals (62 Korean water deer and 15 Korean raccoon dogs [Nyctereutes procyonoides koreensis]) were analyzed to detect Babesia infection. First, PCR was performed to investigate Babesia infection using the partial 18S rRNA gene. Among the 77 wild animals examined, 11 Korean water deer were positive for infection as per the results of sequence analysis. Next, these 11 samples were reanalyzed by PCR using newly designed primers to differentiate between B. capreoli and Babesia divergens. Finally, only one Korean water deer sample was found to be positive for B. capreoli. The nucleotide sequence of this protozoan showed 99.6 − 99.7% identity to those of B. capreoli reported previously. Phylogenetic analysis revealed that the sequence belonged to the B. capreoli clade and diverged from the B. divergens clade. Three nucleotides at positions 631, 663, and 1637 were analyzed to differentiate between B. capreoli and B. divergens. The nucleotides at positions 631 and 1637 were identical to those of B. capreoli, whereas that at position 663 was different. The sequence detected in Korean water deer also exhibited differences at other four positions (669, 704, 711, and 1661) compared with other B. capreoli sequences. This study indicates that Korean water deer serve as a reservoir for B. capreoli. In addition, the primers designed in this study can be used to detect B. capreoli. Future studies should explore the tick vectors that transmit B. capreoli and its pathogenicity. This study was approved by the Institutional Committee of Graduate Studies and Research at Jeonbuk National University, ROK (IACUC decision no.: CBU 2014-00026).
Introduction
Babesia is a tick-borne intraerythrocytic parasite transmitted by Ixodid ticks that infect a wide range of domestic and wild animals, and occasionally humans (Vannier and Krause 2009). In particular, babesiosis is primarily of veterinary importance and leads to significant economic losses in the cattle industry worldwide (Zintl et al. 2003, Bock et al. 2004). Several species of Babesia have zoonotic potential and have been reported to infect humans in Europe and North America (Hildebrandt et al. 2007, Schorn et al. 2011). Among these species, B. divergens is the primary agent of bovine babesiosis (Uilenberg 1995) and has been identified as the causative agent of human babesiosis in Europe (Hildebrandt et al. 2013).
In the United States, Babesia microti causes human babesiosis through blood transfusion (Ord and Lobo 2015). Various wild animals also act as reservoirs for zoonotic babesiosis, and B. divergens, B. capreoli, and Babesia venatorum have been reported to be transmitted through wild cervids in Europe (Razanske et al. 2019). Furthermore, B. divergens is found in cattle (Zintl et al. 2003), whereas B. capreoli is found in roe deer (Capreolus capreolus) (Michel et al. 2014). The clinical signs of babesiosis range from asymptomatic infections to hemolytic anemia with icterus and hemoglobinuria, cardiovascular shock, multiorgan failure (depending on factors such as age and immunocompetence), and even death (Homer et al. 2000, Bos et al. 2017).
According to a recent study conducted by our group, B. capreoli is identified in Korean water deer (Hydropotes inermis argyropus) (Shin et al. 2020). Babesia capreoli infection was first reported in roe deer in 1962 in Germany (Enigk and Friedhoff 1962); subsequently, fatal cases in wild roe deer with parasitemia as high as 20 − 25% were attributed to B. capreoli throughout Europe (Hinaidy 1987, Cancrini et al. 2008, Malandrin et al. 2010). In addition, B. capreoli is considered the causative agent of fatal hemolytic anemia in chamosis (Rupicapra rupicapra) (Schmid et al. 2008). Whether B. capreoli is a zoonotic pathogen remains unclear, but its potential threats to animal and human health cannot be ruled out.
Most importantly, B. capreoli exhibits high sequence identities in the 18S rRNA gene with B. divergens, and the two species may have only three differences in this gene at the nucleotide level (Malandrin et al. 2010). Babesia capreoli and B. divergens have not been clearly differentiated based on morphology or serology; thus, identification of these species requires sequencing analysis (Andersson et al. 2016).
Changes in human behavior, that is, the increased leisure activities have led to closer proximity to tick habitats, resulting in the emergence of zoonotic tick-borne diseases (TBDs). Of note, wild animals serve as the primary reservoirs of various tick-borne pathogens (TBPs) and have been demonstrated to be essential in maintaining and amplifying tick populations in the Republic of Korea (ROK) (Han et al. 2017, Shin et al. 2020).
In the warmer season, people who enjoy leisure activities and farmers may come in direct contact with infected animal blood by inadvertently touching an infected game animal with bare hands as well as tick infestation, increasing chances for the direct transmission of TBPs. Wild animals play an important role in disease transmission from domestic animals to nearby human populations (Daszak et al. 2000). Unlike other TBPs, the distribution and public health relevance of Babesia spp. have been relatively neglected in the ROK. Therefore, this study sought to investigate B. capreoli infection from wild animals in the ROK.
Materials and Methods
Ethics statement
All samples were collected from the study animals after obtaining consent from the provincial Wildlife Rescue and Conservation Centers.
Sample collection
Blood samples were collected from a total of 77 animals found at 6 different Wildlife Rescue Centers: Chungnam, Chungbuk, Jeonbuk, Gyeongnam, and Gyeongbuk Provinces as well as Ulsan city in the ROK. These included 62 Korean water deer (Hydropotes inermis argyropus
DNA extraction, PCR, and nucleotide sequencing
DNA was extracted from the blood samples using the DNeasy Blood and Tissue Kit (Qiagen, Inc., Valencia, CA) according to the manufacturer's instructions and was stored at −20°C until further use. Primers were designed using the Primer3 software (version 4.0.0). Babesia spp. were first screened from rescued wild animals targeting the partial 18S rRNA gene (Schorn et al. 2011). Although B. capreoli was identified through primary sequencing, the amplicon obtained after PCR was short (∼450 bp), thus making it difficult to differentiate between B. capreoli and B. divergens. Therefore, positive samples for the 18S rRNA gene were re-tested using B. capreoli-specific primers designed in this study.
This procedure was used to differentiate among three nucleotides at positions 631, 663, and 1637, and facilitate the identification of B. capreoli and B. divergens and amplification of the 18S rRNA gene with an expected amplicon size of 1699 bp using two pairs of primers (S1F, 5′−CTGGTTGATCCTGCCAGTAG −3′; S1R, 5′−CAAGACAAAAGTCTGCTTGAAAC −3′; S2F, 5′−AGATTGCACTTCGCTTTTGG −3′; S2R, 5′−CGGAAACCTTGTTACGACTTC −3′) (Fig. 1). The expected sizes of S1 and S2 were 711 and 1065 bp, respectively. The cycling conditions were as follows: 94°C for 5 min; followed by 35 cycles of 94°C for 30 s, 60°C for 30 s (S1), or 55°C for 30 s (S2), and 72°C 45 s; and a final extension at 72°C for 10 min.
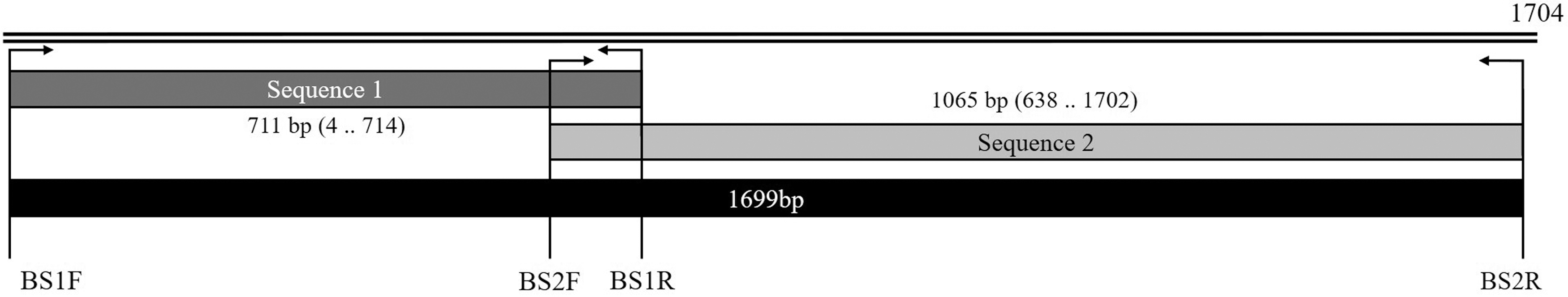
Primers designed in this study. S1F and S1R as well as S2F and S2R were used for the first and second rounds of PCR, respectively. These sequences were joined through overlapping sites.
The S1–S2 sequences were attached. In addition, the specificity of these primers was evaluated using the DNA of a parasite, such as Theileria orientalis, present in the blood and DNA from a healthy animal. Only B. capreoli DNA was amplified but not B. divergens, B. venatorum, and Babesia odocoilei DNAs. In all PCR experiments, distilled water was used as a negative control. The PCR products were separated through electrophoresis on 1.5% agarose gels and visualized by staining with ethidium bromide. The products were purified using the AccuPrep® PCR Purification Kit (Bioneer, Daejeon, ROK) in accordance with manufacturer's instructions and then sequenced directly (Macrogen, Inc., Daejeon, ROK).
Phylogenetic analysis
The nucleotide sequences obtained in this study were analyzed using BioEdit (version 7.2.5) and the Basic Local Alignment Search Tool available from the National Center for Biotechnology Information database. To investigate the homology among B. capreoli genes, nucleotide sequences were aligned using ClustalX and then analyzed by direct comparison with reference sequences from GenBank. A phylogenetic tree was constructed based on nucleotide alignments using the neighbor-joining method. Bootstrap analysis was performed with 1000 replicates using the MEGA X program (Kumar et al. 2018).
Results
Of the 77 rescued wild animals, 11 Korean water deer were positive for the partial 18S rRNA gene of Babesia spp. Babesia DNA was not detected in Korean raccoon dog. These 11 amplicons (450 bp in length) were sequenced, and B. capreoli DNA was identified. These samples identified as B. capreoli in the first round of PCR were reanalyzed using the newly designed primers to further differentiate B. capreoli from B. divergens. Among the 11 samples tested, one sample was poorly sequenced and the remaining 10 sequences were analyzed. Finally, only one sample was positive for B. capreoli and was deposited in GenBank under the accession number of MZ150580, although the remaining nine amplicons were very similar to B. capreoli. Our sequence (MZ150580) had 99.6 − 99.7% identity to those of B. capreoli reported from Europe.
The 18S rRNA gene of B. capreoli differed from that of B. divergens at only three nucleotide positions, namely 631, 663, and 1637, which were GTT for B. capreoli and AAC for B. divergens according to a previous study (Supplementary Fig. S1) (Malandrin et al. 2010). To clarity whether the sequences obtained in this study corresponded to B. capreoli, we analyzed the nucleotides at three positions. In these positions, our sequence (MZ150580) carried the GCT nucleotide (Supplementary Fig. S2 and Table 1), whereas the others had the GCC nucleotide.
Positions of Nucleotide Changes Within Babesia capreoli by the 18S rRNA Gene
“−”: the same nucleotide.
Korean water deer obtained in this study.
The nucleotides present at positions 631 and 1637 of our sequence (MZ150580) were identical to those of B. capreoli, which confirmed the protozoan B. capreoli; however, the nucleotide at position 663 was different from that of B. capreoli. The nine sequences differed by two of three nucleotides. To ensure accuracy, we did not consider these as B. capreoli. The sequence identified in this study showed differences at five positions (663, 669, 704, 711, and 1661) from those of B. capreoli reported in other countries (Table 1). This indicated that these positions are differentiating factors for B. capreoli from the ROK from those of other countries.
Finally, one B. capreoli amplicon (1699 bp) was included in the phylogenetic tree. Phylogenetic analysis based on the 18S rRNA gene revealed that our sequence belonged to the B. capreoli clade and diverged from the B. divergens clade. As given in Fig. 2, B. capreoli and B. divergens are similar. B. capreoli, which includes our sequence, diverged from B. odocoilei and B. venatorum (Fig. 2). Thus, to accurately identify B. capreoli, the difference among the abovementioned three nucleotides should be considered and phylogenetic tree analysis should be performed.
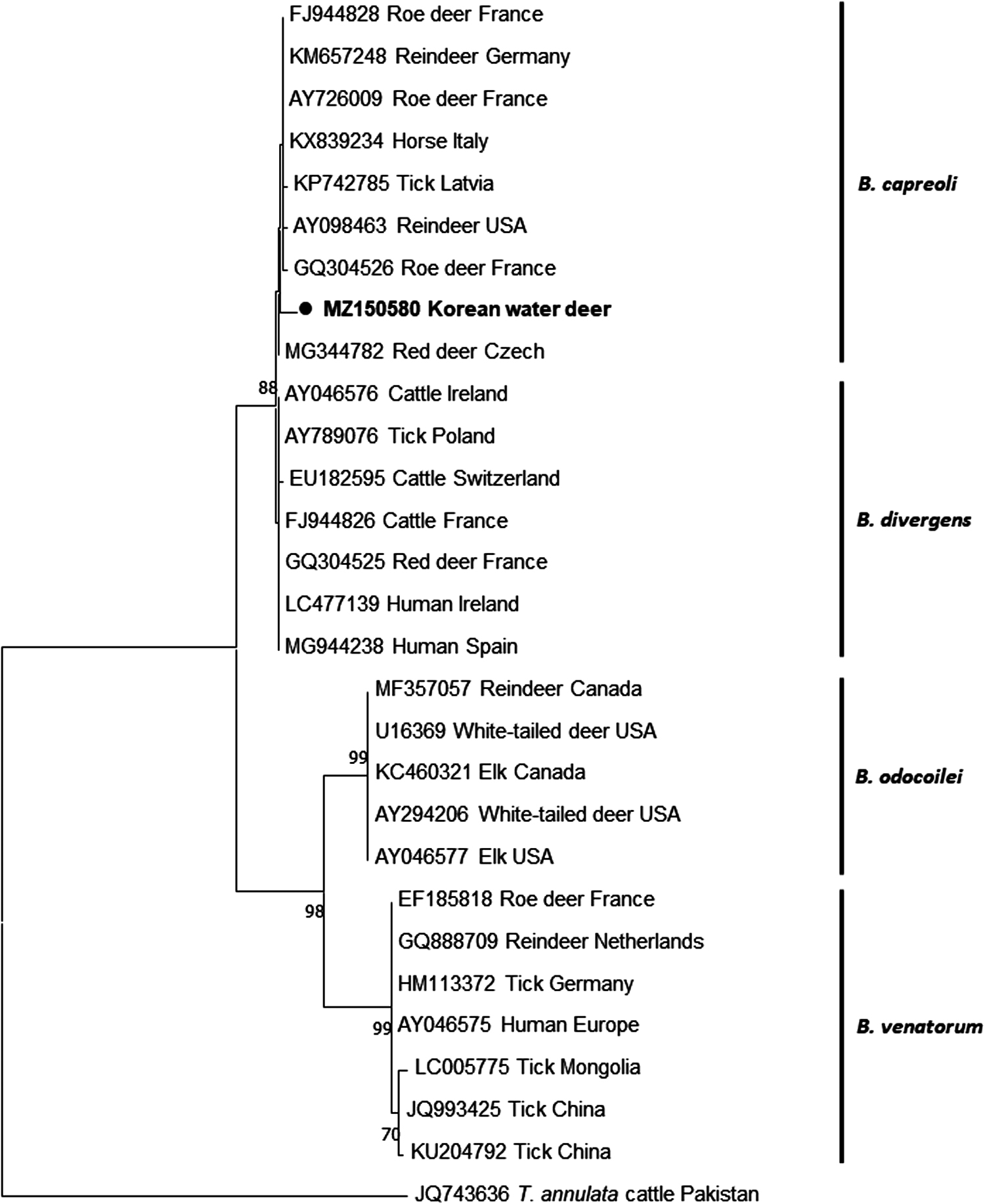
Phylogenetic analysis based on Babesia capreoli 18S rRNA gene sequences (1699 bp). A tree was constructed using the MEGA X software following the maximum likelihood method; the numbers over the branches indicate the bootstrap values as the percentage of 1000 replicates that support each phylogenetic branch. The sequence identified in this study is indicated in bold with a circle symbol. Bootstrap values (1000 replicates) of <70 are not shown.
Discussion
This study reports the incidence of B. capreoli infection in Korean water deer following a PCR-based molecular approach. Babesia capreoli has mainly been reported in wild ruminants in Europe (Hoby et al. 2007, Lempereur et al. 2012, Michel et al. 2014, Zanet et al. 2014, Andersson et al. 2016, Bos et al. 2017) and it is generally nonpathogenic to humans and livestock but occasionally causes fatal diseases in some hosts (Hoby et al. 2007, Malandrin et al. 2010, Wiegmann et al. 2015, Bos et al. 2017). Several studies have reported that B. capreoli causes clinical disease in younger animals more frequently than in adults (Michel et al. 2014, Wiegmann et al. 2015, Bos et al. 2017). Moreover, the clinical signs of B. capreoli infection are more severe in immunocompromised animals, suggesting that this might pose a risk to naive animals.
Because the animals used in this study were rescued from the roadside, predicting the association between the clinical symptoms and B. capreoli infection was difficult. Recently, wild animals in the ROK have been frequently spotted in urban areas because of food scarcity and escape from predators. Moreover, because these animals share the same habitat, there is an increased risk for interspecies transmission of TBDs (Han et al. 2017). The results suggest that Korean water deer serve as a reservoir for B. capreoli. Although the clinical importance of B. capreoli was not evaluated in this study, the possibility of transmission to domestic animals, livestock, and humans cannot be ruled out in the future. Moreover, it is estimated that B. capreoli has existed in the ROK for a long time.
In our previous study, B. capreoli infection in Korean water deer was detected through partial amplification (Shin et al. 2020). Because the amplicon size (450 bp) was inadequate to differentiate between B. capreoli and B. divergens, we used newly designed primers for more accurate identification of B. capreoli in this study. The sequence identified in Korean water deer was confirmed as B. capreoli sequence and exhibited differences at five positions compared with B. capreoli reported in Europe.
Although it is challenging to make a conclusive remark, this may be attributed to the difference of tick vectors that transmit B. capreoli in Europe and the ROK or to the geographical location. Thus, PCR-based studies using sequence analysis are warranted for the accurate identification of B. capreoli. More epidemiological investigations using various wild animals are needed to confirm the presence of B. capreoli and verify whether these differences are distinctive of a Korean lineage or are accidental.
Ixodid ticks are the primary vectors of Babesia spp. Ixodes ricinus is the most prevalent tick species and is known as the main vector of zoonotic infectious disease agents in Europe (Casati et al. 2006, Hilpertshauser et al. 2006, Schorn et al. 2011, Lempereur et al. 2012, Welc-Faleciak et al. 2013, Hamsikova et al. 2016). Babesia capreoli has been detected also in I. ricinus (Schorn et al. 2011, Lempereur et al. 2012, Hamsikova et al. 2016). However, to our knowledge, no studies have explored the presence of I. Ricinus in the ROK yet, whereas I. nipponensis (Japanese hard tick) is commonly distributed and transmits potential zoonotic TBPs (Shin et al. 2013, Kang et al. 2016, Suh et al. 2016).
According to previous studies, I. persulcatus harbors various species of Babesia (Rar et al. 2011, Xia et al. 2020). To the best of our knowledge, there is currently a lack of studies on the association of I. nipponensis or I. persulcatus with the spread of B. capreoli in the ROK. These tick vectors may be of public health concern and pose a threat to humans, domestic animals, and livestock through tick bites. How this pathogen was introduced in the ROK remains unknown. In Europe, roe deer and captive reindeer (Rangifer tarandus tarandus) have been proposed as maintenance hosts for B. capreoli (Michel et al. 2014, Bos et al. 2017). Although B. capreoli was detected in Korean water deer, whether these animals are the primary hosts of B. capreoli remain to be explored. However, it is undeniable that Korean water deer serve as significant hosts for the maintenance and amplification of various tick populations.
Conclusions
Our results showed that B. capreoli infection was detected in Korean water deer. Our sequence (MZ150580) had a 99.6 − 99.7% strong identity to those of B. capreoli reported in Europe. Although the three positions that identify B. capreoli did not match completely, the PCR primers used in this study were suitable for the detection of B. capreoli. However, there had not been many reports of Babesia, it is speculated that its importance has been overlooked in the ROK. Although B. capreoli is not considered a zoonotic species of Babesia, its prevalence and pathogenicity in wild and domesticated animals should be investigated to better understand the importance of B. capreoli as an infectious agent. This study provides new information on B. capreoli, which has not been reported to date in the ROK.
Footnotes
Authors' Contributions
K.S.C. designed the research and wrote the article. Y.J.P., E.M.K., H.C.C., and S.U.S. performed the experiments. K.S.C., J.S.C., and J.H.P. analyzed the data. All authors have read and approved the final version of the article.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This work was supported by the Government-wide R&D Fund for Infectious Diseases Research (HG18C0021).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.