
Abstract
Teschen disease is an acute fatal enterovirus encephalomyelitis of pigs, characterized by a range of central nervous system disorders. The cause of porcine enterovirus encephalomyelitis is the picornavirus porcine teschovirus-1 (PTV-1). There are at least 12 disctinct serotypes of PTVs, where PTV-2 to PTV-12 serogroups are associated with other forms of disease (Talfan disease or poliomyelitis suum) or benign enzootic paresis. Combined, PTVs have been found to have a high seroprevalence, up to 65%, in healthy pig populations in Europe. PTVs have also been detected in wild boar, including the divergent PTV-13 serogroup; wild suids may represent a sylvatic reservoir capable of carrying the virus long distances. In Ukraine, Teschen disease is widespread and causes lethal disease in domestic pigs. To understand temporal and geographical distribution of Teschen disease virus (PTV-1) in wild boar in Ukraine (2001–2013), we analyzed seroprevalence of 6840 blood serum samples from hunted suids using a virus microneutralization assay. A total of 1364 samples (19.9%) were seropositive, with average antibody titer ratios 5.89 ± 0.03 log2 (range 5–12 log2). Teschen seroprevalence was temporally and geographically concentrated in the northern and western regions of Ukraine, corresponding to forested regions (polissya) and overlapping with wild boar populations and habitats, suggesting endemicity in wild boar. The virus sporadically emerged in central, southern, and eastern forested regions, suggesting long-distance movement of infected wild suids. Thus, wild boar should be monitored for potential transboundary spread in forested and mountain regions and spillover of PTVs to domestic swine populations.
Introduction
Research on zoonotic viruses has increasingly focused on wild animal species, with study of viral ecology in wild fauna from a number of perspectives (e.g., biology and ecology of host species, disease susceptibility, virus genomic variation in evolutionary niches) (Daszak et al. 2000).
Epizootological (epidemiological) monitoring of zoonotic diseases in wild boar (Sus scrofa) has received considerable attention, especially in the European Union (EU) countries with large domestic pig populations. In the EU, World organisation for animal health (OIE) data on reportable swine infectious diseases include African swine fever (ASF), classical swine fever virus (CSF), porcine reproductive and respiratory syndrome (PRRS) virus, Aujeszky's disease, and others (Drolet et al. 1999). As vectors of zoonotic pathogens, Eurasian and European wild boar populations have been responsible for the transboundary spread of infectious diseases, between wild and domestic pig populations (Boadella et al. 2012, Guinat et al. 2016, Malmsten et al. 2018).
Zoonotic pathogens of concern in wild boar and domestic pigs are porcine teschoviruses (PTVs), enzootic Picornaviridae comprising at least 13 serotypes, including the etiological agent of Teschen disease (Boros et al. 2012). Teschen disease is an acute fatal enterovirus encephalomyelitis of pigs, characterized by a range of central nervous system (CNS) disorders, first identified in Czechoslovakia by Dr. Trefni in 1929 (Romanenko 2009a,b). Porcine enterovirus encephalomyelitis is caused by the picornavirus PTV-1 (previously PTV). There are few scientific reports on Teschen disease in wild boar compared with Aujeszky's disease, PRRS, porcine circovirus-2 (PCV-2), and swine influenza.
However, both domestic and wild animals are susceptible to Teschen disease (Kolomytsev 2005, Romanenko 2007). In Ukraine, the disease was first recorded by Romanenko in Lviv and Zhytomyr oblasts (1974), Vinnytsia and Kyiv (1980), and subsequently in Rivne and Volyn oblasts of Ukraine (Kolomytsev and Dubrovin 2001, Romanenko 2009a,b, Vabishchevych 2009). Teschen disease occurs in wild boar in Europe and water hogs in Madagascar (Romanenko 2009a,b, Deng et al. 2012), with the most recent major epizootic outbreak occurring in Haiti in 2009 (Romanenko et al. 1992).
Multiple clinical manifestations of Teschen disease in pigs exist and range from severe-to-mild signs and outcomes. Highly virulent strains cause diseases in pigs of all ages, but most often in immunologically naive pigs less than 3 months of age (mortality between 70% to 90%) (FAO 2002). Severe cases result in porcine enteroviral encephalomyelitis. The incubation period for severe infection is 15 days, but can range up to 60 days (Romanenko et al. 1992, Syurin 1998, Romanenko 2007, Romaniuk 2011). Teschen disease affects the CNS, with symptoms including the following: apyetous encephalomyelitis, hypertension, muscle tremor, nystagmus, opisthotonus, tonic and clonic spasm, unsteady gait, paralysis of the extremities, neck muscles, and pharynx (Syurin 1998, Romanenko 2009a,b, Romaniuk 2011).
Encephalomyelitis is acute, and results in death due to respiratory paralysis after 1 to 3 days of symptomatic disease in 80% to 95% of cases (Romanenko 2009a,b, Romaniuk 2011). A myelitic form of Teschen's disease is observed in 20–50% of cases, and results in death due to paralysis of the respiratory system after 1–2 weeks. Teschen's disease can also present chronically (Syurin 1998, Bobruiko 2000, Romanenko 2007, 2009a,b) in adult pigs and can persist up to several months. Subacute infection lasts 6–8 days and is characterized by a low mortality rate (Romanenko et al. 1992). Finally, asymptomatic disease is rarely recorded, but animal stress can contribute to clinically expressed manifestation of Teschen disease (Vlasov et al. 2001a,b).
PTV-1 is a picornavirus of ∼7.1 kb of positive (+)-sense, single-stranded RNA (Zell et al. 2001, Cano-Gómez and Jiménez-Clavero 2016). There are at least 13 distinct serotypes of PTVs. PTV-1 is associated with Teschen disease, while PTV-2 to PTV-12 are associated with milder disease (e.g., Talfan disease or poliomyelitis suum) or benign enzootic paresis (OIE Terrestrial Manual 2017). PTV-13 was recently identified in central Europe (Buzun et al. 2001). PTVs have a high seroprevalence (up to 65%) in healthy pig populations in Europe, and have been detected in wild boar (including PTV-13). Wild suids may represent a sylvatic reservoir capable of carrying the virus long distances and supporting transboundry transmission.
Diagnostics for porcine teshovirus include isolation and growth in a permissive cell line and identification via neutralization by reference serum (Vlasov et al. 2001b), and electron microscopy (Romanenko 2010). Rapid diagnosis can be performed using a fluorescent antibody test (FAT), however, this assay can give a false-negative result (Syurin 1998). If infection is suspected, but negative results are obtained via FAT, cytopathic effect in cell culture (Romanenko et al. 1992) or infection-based bioassays on piglets (Romanenko 2009a,b) have been used to verify infection. PTV-1 has also been identified by enzyme-linked immunoassay (ELISA) (Romanenko et al. 1992), PCR (Buzun et al. 2001, Vlasov 2002, Kaku et al. 2007, Sytiuk et al. 2014), and real-time PCR (Kaku et al. 2007) in pathological samples.
A partial genome of PTV-1 from wild boar sampled in 2013, encoding the VP1 gene, was sequenced (GenBank acc. no. KJ081865.1); this reference has provided a positive control for PCR analyses. However, as preserved viral RNA in serum samples was not consistently available for this study, PCR analysis for PTV-1 has not been not done systematically (including this study). Although PCR analysis of samples from PTV-1 viremic pigs would provide information about incidence, it could underestimate the prevalence of PTV-1 exposure in a swine population if the window of infection is missed. However, PCR and serological assays capable of distinguishing serotypes, coupled with the advent of virus genome sequencing, should contribute to understanding the transmissibility of PTVs.
Thus, this study focused on serological analyses, to understand Teschen disease exposure in wild swine populations in Ukraine. Seroprevalence in 6840 wild boar hunted in Ukraine from 2001 to 2013 was analyzed. Antibodies to PTV-1 were found in 1364 samples (19.9% seropositivity), with significant variation in geographic prevalence, indicative of the movement of the virus in wild boar populations.
Materials and Methods
Serum samples
Six thousand eight hundred forty blood serum samples were collected from wild boar killed in hunting areas in 375 of 490 administrative raions (county-level districts) of all oblasts (province-level administrative divisions) of Ukraine from 2001 to 2013. Each hunting season spanned the winter months (e.g., 2001/02). Serum was collected by an Order of the State Veterinary and Phytosanitary Service of Ukraine (2001–2013, under state-level monitoring for CSF and ASF), under the supervision of the Section for Major Epizootics from the State Regional and District Veterinary Medicine Departments of Ukraine. With the permission of the State Veterinary and Phytosanitary Service of Ukraine and the State Research Institute of Laboratory Diagnostics and Veterinary and Sanitary Expertise, samples were transferred to IVM NAAN for analysis.
Serologial analysis
Specific antibodies against PTV-1 were identified via a viral microneutralization assay using culture-transduced SNEV cells (Romanenko et al. 1995, FAO 2002, Vlasov 2002, Romanenko 2009a,b), ELISA (FAO 2002), and control monoclonal antibodies (Vlasov et al. 2001b). The positive serological control was PTV-1 Perechynskyi-642 strain (1972–74 isolate). Specific humoral antibodies against PTV-1 in serum were evaluated via a virus microneutralization reaction (Romanenko 2009a,b).
Passaged CHEB cell culture was established from fresh primary embryonic pig kidney. CHEB cells were maintained in Dulbecco's modified Eagle's media (DMEM, PAA, Austria) supplemented with 0.02% Versene (PanEco, Russia) and 10% preservative-free intact bovine serum (Bio-Test-Laboratoriia R&D LLC). Cells were passaged using 0.25% Trypsin in phosphate-bufferred saline (PBS, Bio-Test-Laboratoriia R&D LLC). Control PTV-1 was isolated from a symptomatic pig (109.5 lg TCID50/cm3) with a diagnostic blood serum antibody titer of 1:1000 (9.9 log2). This virus was stored at IVM NAAS (Romanenko, personal communication). No cross-reactivity with PTV-2 to PTV-13 has been noted. Serial dilution of the control PTV-1 serum set a titer of 5.0 log2 as the threshold for a seropositive sample.
Statistical and mapping analyses
Serological data at the oblast (provincial) and raion (county-sized district) levels were collated, and longitudinal prevalence maps generated and analyzed using ArcGIS (ESRI). Statistical analyses of serological data by a single-predictor logistic analysis of variance (ANOVA) model using JMP Pro (Version 15) to estimate significance of correlation between seropositivity and hunting season, for the broad regions of Ukraine (northern, eastern western, southern, central, and the country as a whole).
The single-predictor logistic model analysis yielding a whole model test result, which is a likelihood ratio of the chi-squared test calculated by comparing the whole-model fit to the model that omits all the regression parameters except the intercept parameter, was used to determine statistical significance within each of the five individual regions and across Ukraine. This test is appropriate as it approximates ANOVA for continuous responses, with negative log-likelihood corresponding to the sums of squares, and the chi-squared test corresponding to the F test, but for categorical measures. Log/exponential regression (curve fitting) was performed (MS Excel) to estimate distribution of actual titers obtained in serological titer data, with 5.0 log2 determined as the seropositive cutoff.
Results
We studied Teschen disease in the wild boar population in Ukraine by serological analyses. A microneutralization test detected and measured titers of humoral antibodies against PTV-1 in wild boar serum samples to estimate virus seroprevalence. These data were used to map PTV-1 seropositive wild boar across administrative raions (county-level districts) of Ukraine.
Collection and serological analysis of wild boar samples in Ukraine
To understand Teschen disease seroprevalence rate in wild boar, sera were collected from hunted wild boar spread geographically across raions and oblasts (province-level districts), and temporally over 12 hunting seasons, in Ukraine. Serological monitoring covered all 24 oblasts, plus Kyiv and AR Crimea, inclusive of 375 (76%) of 490 raions in Ukraine (Table 1).
Blood Sera Collection in Hunted Wild Boar in Ukraine, by District (Raion)
Serological testing for PTV-1 antibodies identified 1364 positive samples (overall seroprevalence, 19.9%, Table 2), with antibody titers ranging from 5 log2 to 12 log2. The average level of PTV-1 antibodies in wild boar sera was 5.89 ± 0.03 log2, with the dominating titer (mode) as 5 log2. Considerable variation in prevalence was observed in the spatial distribution of infected animals in the subprovincial-level administrative raions of Ukraine. Overall, raions containing PTV-1-positive samples are geographically concentrated in the forested regions in the northern, western, and eastern regions of Ukraine, and with fewer positive raions in the territories of the south-central and southern regions (Fig. 1).

Teschen disease seropositive wild boar in Ukraine. Distribution map of the administrative raions (county-level districts), with locations where Teschen disease virus-seropostive wild boar were detected in 2001–2013 marked. Darker red color indicates maximum detection events. Color images are available online.
Teschen Disease Seroprevalence in Hunted Wild Boar in Ukraine, 2001–2013
“—” means no tested and positive blood serum samples.
Seroprevalence of PTV variation by hunting season
During hunting seasons, wild boar sera were collected in forest and hunting areas geographically located in a designated hunting district, or districts (within raions). Since 2001, the number of samples collected increased across the key forested regions, reaching maximum in 2012–2013 (268; Table 2). The average percentage of raions submitting sera was 76.5% [western (90.6%), southern (62.9%), northern (93.2%), eastern (71.4%), and central (62.6%)]. Serological monitoring covered all raions of Volyn, Khmelnytskyi, Odesa, Zhytomyr, and Chernihiv oblasts.
To reduce geographic sampling biases, we conducted a sampling trend analysis at the subprovincial raion (district) level. Trend analysis showed that the number of raions from which sera were collected varied by hunting season: 2001–2002 (6.7% of raions were surveyed), 2002–2003 (6.9%), 2003–2004 (17.3%), 2004–2005 (22.0%), 2005–2006 (10.4%), 2006–2007 (28.2%), 2007–2008 (23.7%), 2008–2009 (32.9%), 2009–2010 (47.6%), 2010–2011 (17.1%), 2011–2012 (54.3%), and 2012–2013 (54.7%). Over 12 seasons, serological monitoring covered all 24 administrative oblasts, as well as Kyiv and AR Crimea, of Ukraine.
Spatial distribution of PTV-1 seroprevalence in wild boar
The spatial distribution of administrative districts (raions), where antibodies against PTV-1 were detected during the 2001–2013 hunting seasons indicated that wild boar were exposed in all Ukrainian oblasts (Fig. 1). The raion-level trend analysis suggested that the intensity of PTV-1 detection varied considerably, overlapping with wild boar habitats, particularly forested and mixed forest (pollisiya) regions. Seropositive samples were geographically concentrated in the northern, western, and eastern regions, and less localized in the central and southern regions (Fig. 1). A general trend of increasing seroprevalence was observed, with variation among regions (Table 2).
Spatial and temporal seroprevalence of Teschen disease in wild boar in Ukraine
To better understand regional patterns of Teshcne disease emergence, the 6840 sera samples collected with a varying number across hunting seasons (Table 1) were analyzed across time in broad regions of Ukraine. The highest number of samples tested was in 2011–2012 (1849 samples), and the lowest in 2001–2002 (68 samples). In total, 1364 samples (19.94%) were positive (Table 2). PTV-1 seroprevalence was 19.94%: western region (25.81%), southern (16.88%), northern (18.38%) eastern (19.90%), and central (16.84%). A single-predictor logistic model was used to test whether the hunting season was a correlated predictor of the proportion of seropositive PTV-1, for each region of Ukraine.
In western Ukraine, the proportion of wild boar that tested positive for PTV-1 was marginally significant across hunting seasons, from 2001 to 2013 (chi-squared = 16.90, p = 0.0511). However, the wild boar seropositive for PTV-1 showed significant variation across hunting seasons (2001–2013) in the northern (chi-squared = 38.23, p < 0.0001), southern (chi-squared = 40.82, p < 0.0001), eastern (chi-squared = 33.25, p = 0.0002), and central (chi-squared = 30.73, p = 0.0012) regions, and across Ukraine for all seasons as a whole (chi-squared = 43.48, p = < 0.0001). Seroprevalence trended highest in the 2008/2009-2009/2010, and 2011/2012-2012/2013 hunting seasons.
Geographic patterns of Teschen disease seropositivity varied across Ukraine but were dominated by forested and mixed forest regions (north-central and west) that are wild-boar habitat (Fig. 1). In the western region, average rates of PTV-1 seroprevalence were higher than those in all other regions (19.94%), by oblast: Volyn (21.54%), Zakapattia (26.67%), Ivano-Frankivsk (35.76%), Lviv (20.34%), Rivne (30.90%), Ternopil (32.76%), Khmelnytskyi (23.77%), and Chernivtsi (24.19%) (Table 2). In southern Ukraine, seroprevalence was only higher than the average in Zaporizhia Oblast (21.20%), with lower rates in other oblasts: AR Crimea (18.56%), Mykolaiv (9.59%), Odesa (16.06%), and Kherson (8.0%). In northern Ukraine, average rates were higher than average in only Chernihiv Oblast (21.67%).
In the remaining oblasts, rates were lower than average: Zhytomyr (18.80%), Kyiv (16.57%), and Sumy (14.15%). In eastern Ukraine, average rates were higher than average in Donetsk (20.78%) and Kharkiv (22.47%), and lower in Luhansk Oblast (15.86%). In central Ukraine, average rates were higher than average only in Vinnytsia Oblast (20.13%); lower rates were observed in Dnipropetrovsk (11.11%), Poltava (9.2%), Kirovohrad (9.92%), Poltava (18.33%), and Cherkasy (17.71%).
Temporal and geographical Teschen disease trends in wild boar
The number of PTV-1 seropositive samples varied by year (Table 1), with upward and downward periods of seropositivity (Fig. 2). An increase in positive samples (from 4.41% to 23.41%) was recorded during the 2001–2002 and 2003–2004 seasons. Subsequently, in the 2004–2005 season, the seropositivity rate fell to 15.84%, and then grew to 27.51% (maximum) in 2006–2007. During the following three seasons, seroprevalence rates decreased and remained practically unchanged: 2007–2008 (19.3%), 2008–2009 (18.29%), and 2009–2010 (19.25%). During 2010–2011, the seroprevalence rate fell to 14.71%. Over subsequent seasons, seroprevalence rates were higher in 2011–2012 (20.23%) and 2012–2013 (21.0%) than the average rate (Fig. 2). In geographic regions of Ukraine, seropositivity varied significantly (4.4–27.5%) across all hunting seasons (2001–2013).
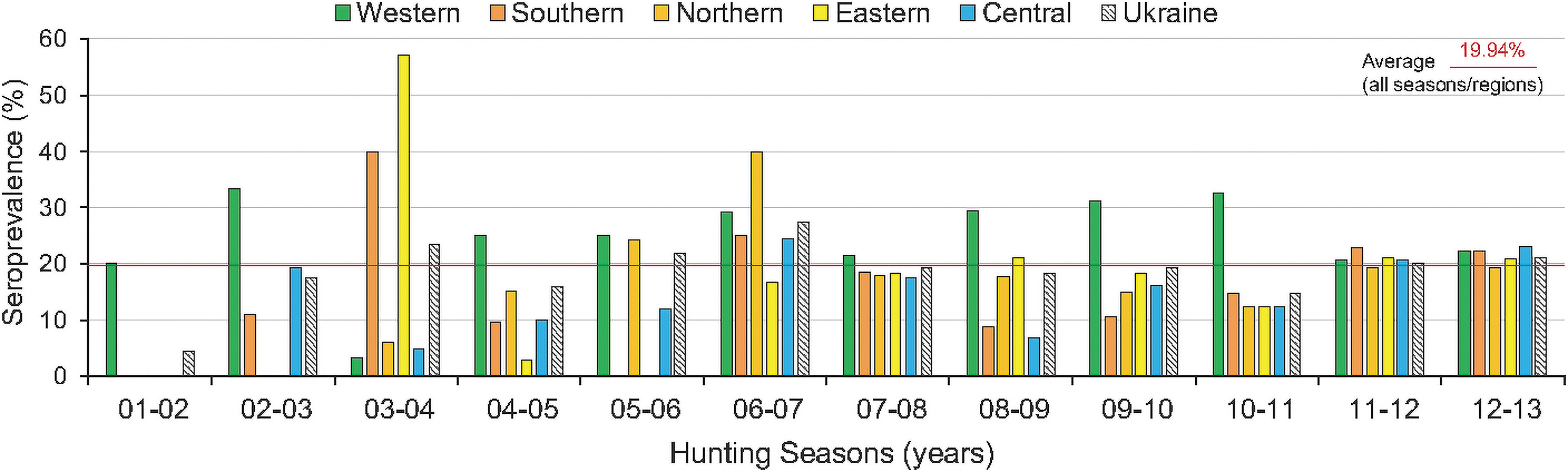
Seroprevalence and geographic distribution of Teschen disease in wild boar in Ukraine. Teschen disease seroprevalence in hunted wild boar in Ukraine over 12 hunting seasons, 2001–2013. Seroprevalence was determined from the 6840 sera samples collected, of which 1364 samples (19.94%, average seroprevalence, indicated) were positive among the western, southern, eastern, northern, and central regions of Ukraine (colors indicated); all Ukraine (cross-hatched). Color images are available online.
A general trend was observed in that PTV-1 seropositive samples were found in a decreasing number of regions over time, suggesting that the geographic appearance of the disease was sporadic (Fig. 1). However, seropositive samples were found in most regions of Ukraine over all the sampled hunting seasons (Fig. 2). To understand the finer distribution of seropositive wild boar, we examined PTV-1 seropositivity in subprovincial-level districts (raions) of Ukraine. Across all seasons (2001–2013), 292 of 375 (78%) raions sampled contained positive samples (Table 2). No PTV-1-positive samples were detected in 83 (22%) of sampled raions.
Quantitative ratios between raions and the number of hunting seasons when positive animals were detected in them were calculated: 99 raions with detection in 1 hunting season (99/1), 75/2, 51/3, 31/4, 21/5, 7/6, 5/7, and 3 raions with detection in 8 hunting seasons, a trend that follows a logarithmic decay pattern (50lnx, R = 0.98). These results suggest that PTV-1 exhibits enzootic circulation in the wild boar population across the geographic span of Ukraine, with more focused localization and geographic concentration in the northern and western regions, and less localization in oblasts of the eastern, central, and southern regions (Fig. 1).
Seroprevalence and antibody titer against Teschen disease virus in wild boar
To identify levels of humoral antibodies against PTV-1 in wild boar sera, antibody titers were analyzed. In previous studies (Romanenko et al. 1992), a 5 log2 or higher antibody level was used to incidicate a positive diagnostic antibody titer. Of the total number of sera samples tested, 1364 samples (19.9%) reached this seropositive level (range: 5 log2 to 12 5 log2 neutralizing antibody titer) (Fig. 3).
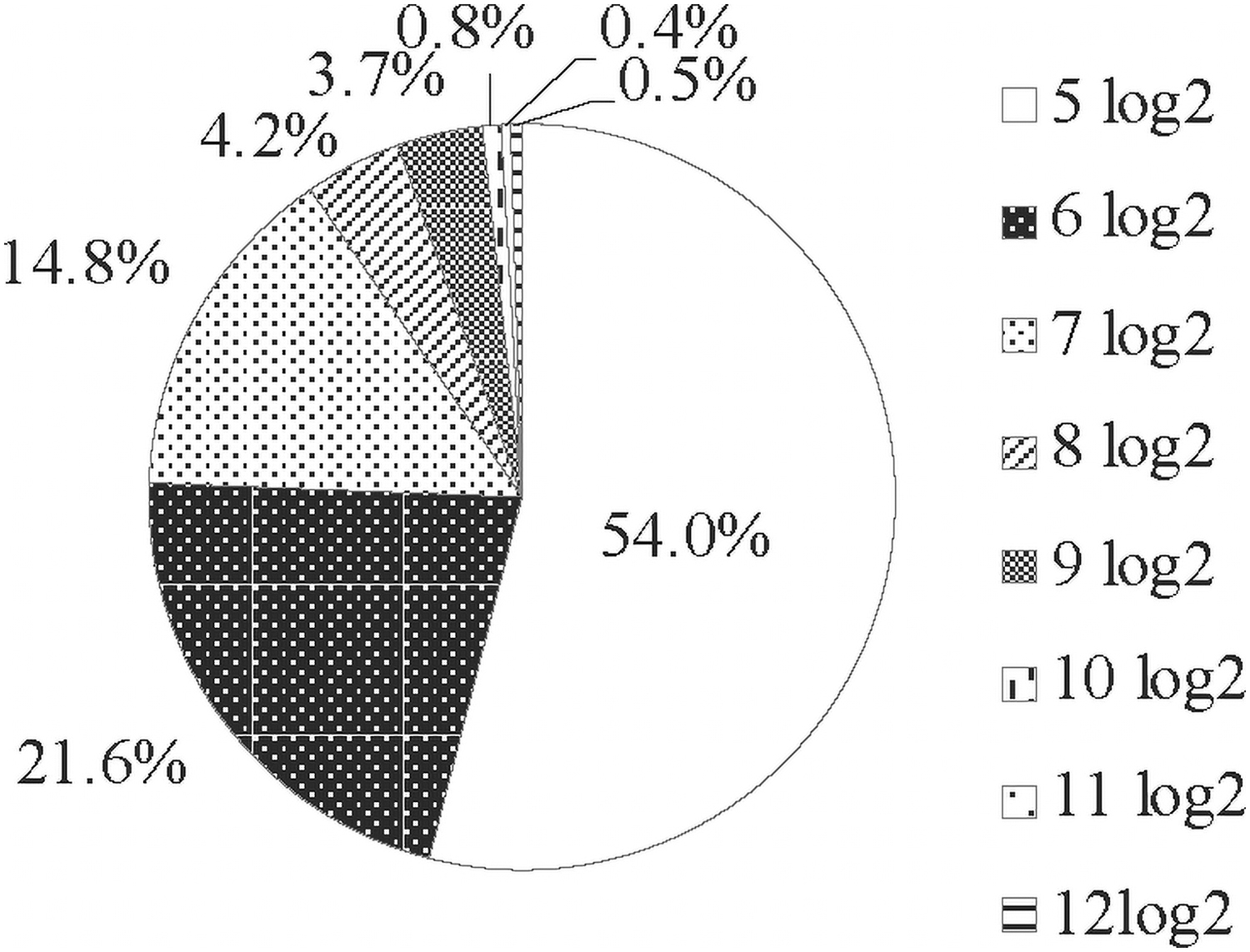
Distribution of antibody titers against Teschen disease in wild boar. Distribution of antibody titers against Teschen disease virus in 1364 seropositve wild boar across the study (2001–2013).
The distribution of antibody titers across all the 1364 positive samples was as follows: 54.0% (736 samples) with 5 log2 antibody titers; 21.6% (295 samples) with 6 log2; 14.8% (202 samples) with 7 log2; 4.2% (57 samples) with 8 log2; 3.7% (50 samples) with 9 log2; 0.8% (11 samples) with 10 log2; 0.4% (6 samples) with 11 log2; and 0.5% (7 samples) with 12 log2. While the titer distribution followed an exponential decay (fraction = 19.3e−0.7[titer], R = 0.96), the average titer of antibodies against PTV-1 among positive samples was 5.89 ± 0.03 log2, illustrating the dominance of the 5–6 log2 titers.
The highest level of postinfection antibodies (12 log2) against PTV-1 may reflect a very recent and/or severe infection; however, virus titer data were not analyzed in our study. The negative exponential distribution may indicate that variations in antibody levels in wild boar are associated with the the dynamics of PTV-1 infection in a field setting, with variations in time elapsed since exposure, infectious dose, and individual susceptibility of animals.
Discussion
Teschen disease is a contagious enzootic disease that impacts both domestic and wild pigs, caused by PTV-1, a virus that circulates in Eastern Europe, Haiti, and Madagascar. Our study shows that Teschen disease is present in wild boar in Ukraine, a population of wild swine that frequently crosses forested borders to neighboring countries (e.g., Moldova, Russia, Belarus, and Poland). The emergence of virulent ASF in this region has highlighted the role of wild boar transboundry vectors for spread of swine virues, and the interactions between wild and domestic swine in central and eastern Europe (Kovalenko et al. 2019, Mazur-Panasiuk et al. 2019).
Teschen disease seroprevalence in Ukraine cannot easily be compared with data from other countries, since there are considerably fewer longitudinal studies in Teschen disease in wild boar compared with Aujezky`s disease, PRRS, ASF, and other pathogens that impact commercially important domestic swine.
Serological analysis of wild boar for Teschen disease in Ukraine was conducted using microneutralization surveillance of 6840 sera samples obtained from hunted animals across a 12-year period (2001–2013). With an overall proportion of positive blood serum samples of 19.94%, and variation by regions (dominated by “hotspots” in western Ukraine), these yearly records of seropositivity wild boar suggest that PTV-1 continuously circulates in the wild boar population. Teschen seroprevalence was geographically focused in the northern and western forested regions (polissya) of Ukraine, overlapping with wild boar populations and habitat. The virus sporadically occurred in central, southern, and eastern forested regions, suggesting long-distance movement of infected wild boar or interactions with infected domestic swine.
The geographic dispersion of seropositive wild boar is stark, suggesting that PTV-1 infection is temporally and geographically dynamic. However, the sporadic nature and geography of outbreaks year-to-year suggest that additional ecological and serological data analyses are needed to understand the role of wild boar populations in the emergence and persistence patterns of Teschen disease in Ukraine. In other words, as a semiresident or migratory subpopulation of wild boar becomes immune, neutralizing antibody titers may decrease (to the lower end of the titer range, near the average of 5.89 ± 0.03 log2), and the virus locally “burns out.”
Meanwhile, wild boar in neighboring districts may suffer zoonotic outbreaks among susceptible or immunologically naive animals. Thus, as a whole, wild boar appear to be enzootic carriers of PTV-1 in Ukraine, with regionally and temporally specific epidemic outbreak patterns.
We have presented a seasonal and geographic picture of Teschen disease seroprevalence in wild boar in Ukraine, highlighting locations that may be useful for future modeling of wild boar in the emergence of swine diseases. Additional studies of the geography of PTV-1 distribution, and immune titers coupled to wild boar age, population distribution, and habitats, are warranted to understand the changing landscape of Teschen disease in Ukraine.
Footnotes
Acknowledgments
The authors extend special thanks to the scientific staff at Metabiota and Battelle for help with preparation of the article, including Yaryna Masniuk, Andrii Pavlenko, Irina Dorochek, Nataliya Mikhalovska, and David Mustra. They also would like to express their profound gratitude to NAAS member V.P. Romanenko for providing the strain and the positive blood serum for the neutralization test; and to IVM scientists Natalia Gutz and Ganna Kovalenko for helpful discussions.
Authors' Contributions
The contents of this publication are the responsibility of the authors and do not necessarily reflect the views of DTRA, BTRP, or the U.S. Government, the Battelle Memorial Institute, or the Government of Ukraine.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
The research here was supported by funding from the State Veterinary and Phytosanitary Service of Ukraine (2001–2013, under state-level monitoring for CSF and ASF), and the Section for Major Epizootics from the State Regional and District Veterinary Medicine Departments of Ukraine. General institutional support for research staff from NSC IVM, Ukraine, and the University of Alaska Anchorage, is also acknowledged.
The U.S. Department of Defense, the Defense Threat Reduction Agency (DTRA), and the Biothreat Reduction Program (BTRP) supported the development of this article. While DTRA/BTRP did not support the research activities described in this publication, the Program supported the presentation of this research in an international forum, and grantsmanship training related to the development of this article through the BTRP (CTRiC) Scientific Writing Mentorship Program.