
Abstract
Objectives:
The emergence of SARS-CoV-2 lineage B.1.617 variants in India has been associated with a surge in the number of daily infections. We investigated the pathogenic potential of Kappa (B.1.617.1) variant in Syrian golden hamsters.
Methods:
Two groups of Syrian golden hamsters (18 each) were inoculated intranasally with SARS-CoV-2 isolates, B.1 (D614G) and Kappa variant, respectively. The animals were monitored daily for the clinical signs and body weight. Throat swab, nasal wash, and organ samples (lungs, nasal turbinate, trachea) were collected and screened using SARS-CoV-2-specific RT-qPCR. Histopathologic evaluation of the lung samples was performed.
Results:
The hamsters infected with the Kappa variant demonstrated increased body weight loss compared to the B.1 lineage isolate. The highest viral RNA load was observed in the nasal turbinate and lung specimens of animals infected with both variants. A significantly higher sgRNA load was observed in the nasal swabs (7 DPI), trachea (3 DPI), and lungs (3 DPI) of hamsters infected with the Kappa variant. Neutralizing antibody response generated in the B.1 lineage-infected hamster sera were comparable against both B.1 and Kappa variant in contrast to Kappa variant-infected hamsters, which showed lower titers against B.1 lineage isolate. Gross and microscopic evaluation of the lung specimens showed severe lung lesions in hamsters infected with Kappa variant compared to B.1.
Conclusions:
The study demonstrates pathogenicity of Kappa variant in hamsters evident with reduced body weight, high viral RNA load in lungs, and pronounced lung lesions. Both Kappa variant- and B.1-infected hamsters produced neutralizing antibodies against both variants studied.
Introduction
Since, the first report of severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2) in Wuhan, China in 2019, the virus has constantly evolved leading to the emergence of new variants (Baric 2020, Chen et al. 2020). The first variant of SARS-CoV-2 with D614G mutation belonging to the B.1 lineage became dominant and was prevalent worldwide since March 2020. Later mutations at different amino acid positions were reported with regional dominance and were designated as variants under investigation (Mercatelli and Giorgi 2020). A few of these were later designated as variant of concerns (VOC) considering their increased risk to public health with increased transmissibility, pathogenicity, and decrease in the effectiveness of vaccines and therapeutics.
The VOCs B.1.1.7 (Alpha), B.1.351 (Beta), P1 (Gamma), and B.1.617.2 (Delta) have seriously affected many countries and posed major public health challenges (WHO 2021). The second wave of the SARS-CoV-2 has hit India hard and affected the population in large numbers. The number of new cases reported in India per day reached ∼0.4 million cases during May 2021(CSSEGIS 2021). The state of Maharashtra shared 21% of the active cases among the reported cases from the country (PRS legislative research 2021). The majority of these cases in Maharashtra have reported to be caused by the SARS-CoV-2 B.1.617 lineage and its sublineages (Cherian et al. 2021). B.1.617 variants have been associated with a surge of cases spreading rapidly not only in Maharashtra but also have been identified in other states of India and abroad.
The B.1.617 variant has eight amino acid changes in the spike region. Multiple sublineages evolved within B.1.617 lineage-like B.1.617.1 (Kappa), B.1.617.2 (Delta), and B.1.617.3 with a different set of unique substitutions and deletions (Cherian et al. 2021). The Kappa variant and B.1.617.3 are characterized by the presence of L452R, E484Q, and P681R amino acid changes, while the B.1.617.2 lacks the E484Q mutation. The presence of L452R, E484Q, and P681R is linked with increased hACE2 receptor-binding affinity and possible NAb escape (Starr et al. 2020, Tchesnokova et al. 2021).
The origin of the Kappa variant is still unknown and as mentioned above, the presence of this variant has been identified in many countries (WHO 2021). Our recent study has shown that Covaxin (a whole virion inactivated vaccine) and Covishield (ChAdOx -1 nCoV-19) immunization generates neutralizing antibodies against the B.1.617 variant (Yadav et al. 2022a, 2022b). Other than a few studies on the neutralization efficiency of vaccinated individuals against the Kappa variant, the other properties of this variant is not yet studied.
Syrian hamsters are a widely used animal model for SARS-CoV-2 and the clinical signs/or endpoints of infection are weight loss and viral RNA/infectious virus titer in target organs (Brocato et al. 2020, Mohandas et al. 2020). In an earlier study, we investigated the pathogenicity of the SARS-CoV-2 B.1 and B.1.1.7 lineage isolates in the Syrian hamster model (Mohandas et al. 2021a). It was observed that the B.1 lineage isolate of SARS-CoV-2 produced interstitial pneumonia with marked alveolar damage and type-II pneumocyte hyperplasia in hamsters (Mohandas et al. 2021a, 2021b). In the present study, we investigated the viral RNA load and pathogenicity of the Kappa variant in the Syrian hamster model and compared it with that of a B.1 lineage isolate.
Materials and Methods
Ethics statement
The animal experiments were performed with the approval of the Institutional Animal Ethics committee and all the experiments were performed as per the guidelines of Committee for the Purpose of Control of Experimentation on Animals (CPCSEA), Government of India (NIV/IAEC/2021/MCL/01).
Experimental design
Male, Syrian golden hamsters (Mesocricetus auratus) of age 6–8 weeks procured from a CPCSEA-approved facility were used. The hamsters were housed in individual ventilated cages with feed and water ad libitum in the containment facility. Eighteen hamsters each were inoculated intranasally with 0.1 mL of 3.16 × 104/mL TCID50 dose of SARS-CoV-2 isolates that is, B.1 (GISAID number: EPI_ISL_420545) and B.1.617.1 (Kappa) (GISAID number: EPI_ISL_8327860) lineages propagated at ICMR-NIV, Pune as described earlier under isoflurane anesthesia (Sarkale et al. 2020, Supplementary Table S1). Clinical signs and body weight of animals were observed daily and six hamsters each were euthanized on day 3, 5, and 7 days postinfection (DPI) to perform necropsy.
Throat swab, nasal wash, and organ samples (lungs, nasal turbinate, trachea) were collected following the necropsy. Throat swabs were collected using sterile nylon flocked swabs (Himedia, Mumbai) in 1 mL viral transport medium (Himedia, Mumbai). Nasal washes were collected using 0.1 mL sterile PBS and were transferred to 1 mL viral transport medium. Organ samples, swabs, and nasal washes were stored at −80°C till further use. A portion of the lungs sample was also collected in 10% neutral buffered formalin.
RT-qPCR
Nasal wash and throat swab collected in viral transport medium (Himedia, Mumbai) and weighed organ samples (lungs, nasal turbinate and trachea) homogenized in 1 mL sterile tissue culture media (Sigma Aldrich) were used for RNA extraction using MagMAX™ Viral/Pathogen Nucleic Acid Isolation kit as per the manufacturer's instructions.
RT-qPCR was performed for SARS-CoV-2 E genomic RNA (gRNA) and N subgenomic RNA using_published primers as follows. The primers and probes used for gRNA were 5′-ACAGGTACGTTAATAGTTAATAGCGT-3′ (forward), _ 5′-ATATTGCAGCAGTACGCACACA-3′ (reverse) and FAM-ACACTAGCCATCCTTACTGCGCTTCG- BHQ and for sgRNA were 5′-CGATCTCTTGTAGATCTGTTCTCT-3′ (forward), 5′-TCTGGTTACTGCCAGTTGAATCTG-3′ (reverse) and 5′-FAM-ACCCCGCATTACGTTTGGTGGACC-3 BHQ′. Primers and probes used for the reference gene HPRT are 5′-TGCGGATGATATCTCAACTTTAACTG-3′ (forward), 5′-AAAGGAAAGCAAAGTTTGTATTGTCA-3′ (reverse) and FAM-AAAGAATGTCTTGATTGTTGAAGGTAAAACTGACATTGG-BHQ.
RT-qPCR was performed using 4 μL of 4 × PCR Master Mix (Life Technologies, New York, NY), 0.5 μL of primer, 0.5 μL of probe, 5 μL RNA, and nuclease-free water to make up a 20 μL reaction volume. Separate reaction was set for each gene. Thermal cycling conditions followed were for E gene were as follows: 50°C for 5 min for reverse transcription, 95°C for 20 s (initial denaturation), 95°C for 5 s, and 60°C for 30 s for 40 cycles in a Real-Time PCR machine (Bio-Rad CFX96). For N sgRNA and HPRT, the thermal cycling conditions used were similar except the annealing temperature, which was 55°C for 30 s. In vitro transcribed RNA was used as positive controls (Choudhary et al. 2020, Perera et al. 2020).
Anti-SARS-CoV-2 IgG ELISA
The antibody response was assessed using an in-house qualitative ELISA assay as described earlier (Shete et al. 2021).
Histopathology
Lung samples collected during necropsy were fixed in 10% neutral buffered formalin (Thermo Scientific). The tissues were processed by routine histopathological techniques for hematoxylin and eosin staining (Culling 2013). The slides were coded before being examined and the pathologist was blinded. The lung lesions were graded from 0 to +4 as nil, minimal, mild, moderate, and severe based on the vascular changes like congestion and hemorrhages, bronchial pathology, degenerative changes, loss of epithelium, hyaline changes, pneumocyte hyperplasia, alveolar septal thickening, and inflammatory cell infiltration.
Plaque reduction neutralization test
The assay was performed against both the isolates. Four-fold serially diluted hamster serum samples were mixed with an equal amount of virus suspension and incubated at 37°C for 1 h. Further 0.1 mL of the mixture was inoculated in a 24-well tissue culture plate (Corning) containing a confluent monolayer of Vero CCL-81 cells (ATCC) and was incubated at 37°C for 60 min and overlay medium containing 1% carboxymethyl cellulose in 2 × MEM (Sigma-Aldrich) was added to the cell monolayer, which was further incubated at 37°C in 5% CO2 incubator for 4–5 days and PRNT50 titers were calculated.
Statistical analysis
GraphPad Prism version 8.4.3 software (GraphPad Software, Inc., San Diego, CA) was used to analyze the data. A nonparametric two-tailed Mann–Whitney test was used to compare between two groups and p-values of <0.05 were considered to be statistically significant.
Results
Kappa variant infection leads to weight loss in hamsters
Weight loss was observed in a higher percentage of hamsters infected with the Kappa variant in comparison with the B.1-infected hamsters (Fig. 1). It was observed that both the B.1- and Kappa variant-infected hamster groups demonstrated a decline in the body weight starting from the 3rd day postinfection (DPI). The mean value for the percent change in bodyweight of B.1-infected hamsters was −0.045 ± 2.54 n = 18) at 3 DPI. The mean body weight decreased further by 1.23 (n = 12) and 0.13 (n = 6) percent on the 5th and 7th DPI. In the Kappa variant-infected hamsters, a significant mean body weight loss of −2.98 ± 2.65 (n = 18, p = 0.0075), −10.690 ± 3.71 (n = 12, p = 0.0001), and −14.877 ± 3.18 (n = 6, p = 0.0022) were observed on 3, 5, and 7 DPI.
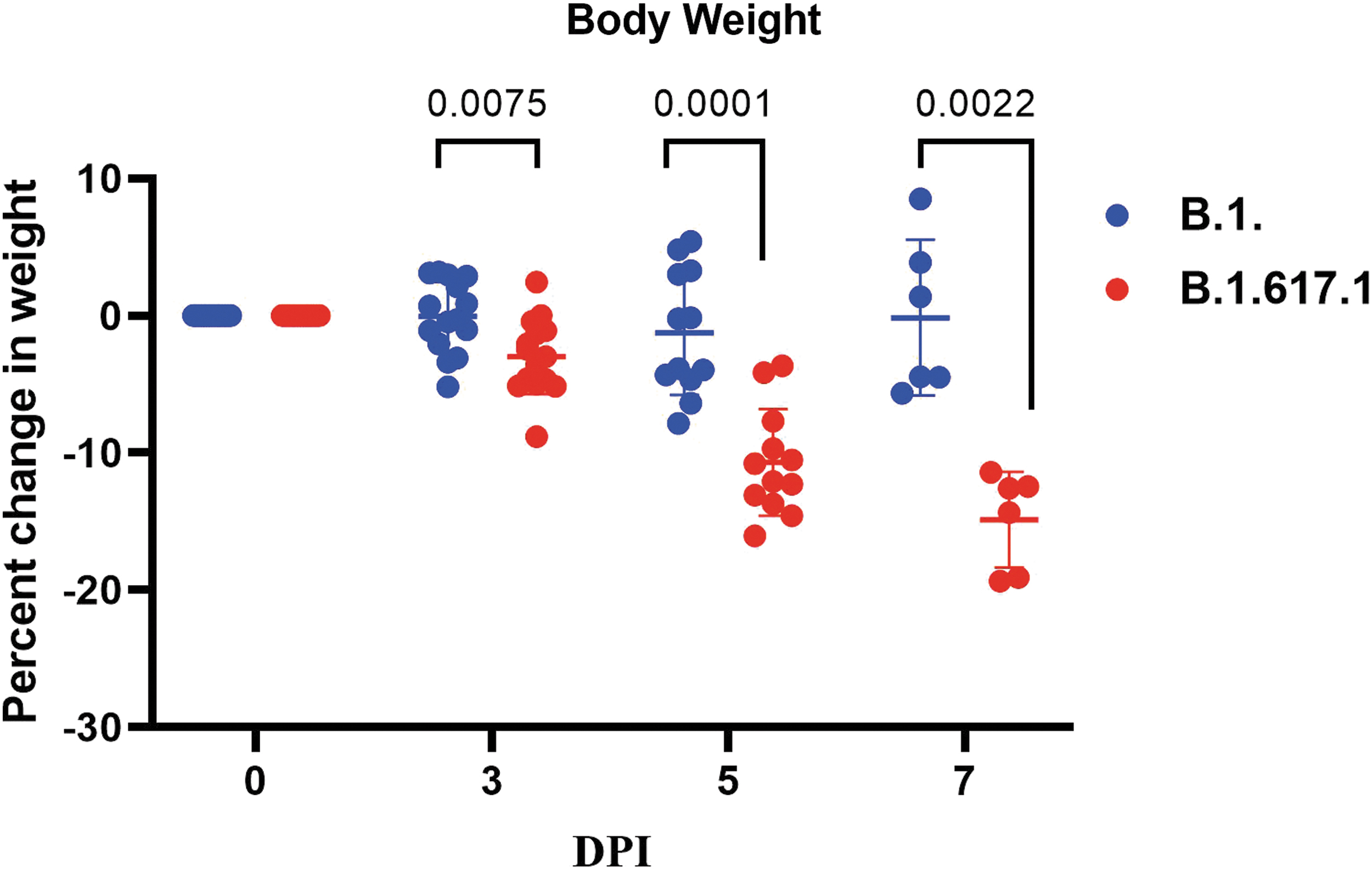
Percent body weight change in the hamsters infected with SARS-CoV-2: The body weight change in hamsters following exposure among hamsters to SARS-CoV-2 variants B.1 and Kappa. The statistical significance of difference in weight loss between the SARS-CoV-2 variants B.1 and Kappa was assessed using the two-tailed Mann–Whitney test between two groups; p values of <0.05 were considered to be statistically significant. Mean and standard deviation is depicted for animals in each group.
Genomic and subgenomic SARS-CoV-2 viral RNA loads in hamster infected with B.1 and Kappa variant
The viral gRNA loads in the throat swab, nasal wash, nasal turbinates, and trachea samples were found comparable and did not show any statistical significance between the two variants on 3 DPI. The lung sample showed significantly high gRNA (p = 0.0152) and sgRNA load (p = 0.0411) on 3 DPI. In the nasal turbinates, the gRNA load was comparable in hamsters of both groups except on 7 DPI, where the viral gRNA load was found significantly higher in case of B.1 variant. Subgenomic RNA (sgRNA) could be detected in all the samples up to 7 DPI. The sgRNA levels of trachea (p = 0.0411) and lungs (p = 0.039) were found significantly higher in Kappa variant group on 3 DPI. The viral RNA load showed a decreasing trend on the 7th DPI in all the samples of hamsters infected with both the variants (Fig. 2A–J).

Scatter plot for the SARS-CoV-2 genomic and subgenomic viral RNA loads in hamsters using RT-qPCR: Genomic viral RNA titers in the swab and organ samples of hamsters infected with SARS-CoV-2 variants B.1 and Kappa.
Antibody response in hamsters postinfection
Neutralizing antibody (NAb) response in hamsters was observed from the 5th DPI against B.1 and Kappa variants in infected hamsters, whereas the anti-SARS-CoV-2 IgG antibodies could be detected by ELISA at the 7th DPI except in one of the hamsters infected with B.1 (Fig. 3A). The variation of antibody response was found to be not significant between the groups both by ELISA and neutralization. NAb geometric mean titer (GMT) for the Kappa variant-infected hamsters against B.1 was 200.4, while against Kappa was 982 on 7 DPI. In case of B.1-infected hamsters, GMT was 391.8 against B.1 and 307.1 against Kappa (Fig. 3B, C).
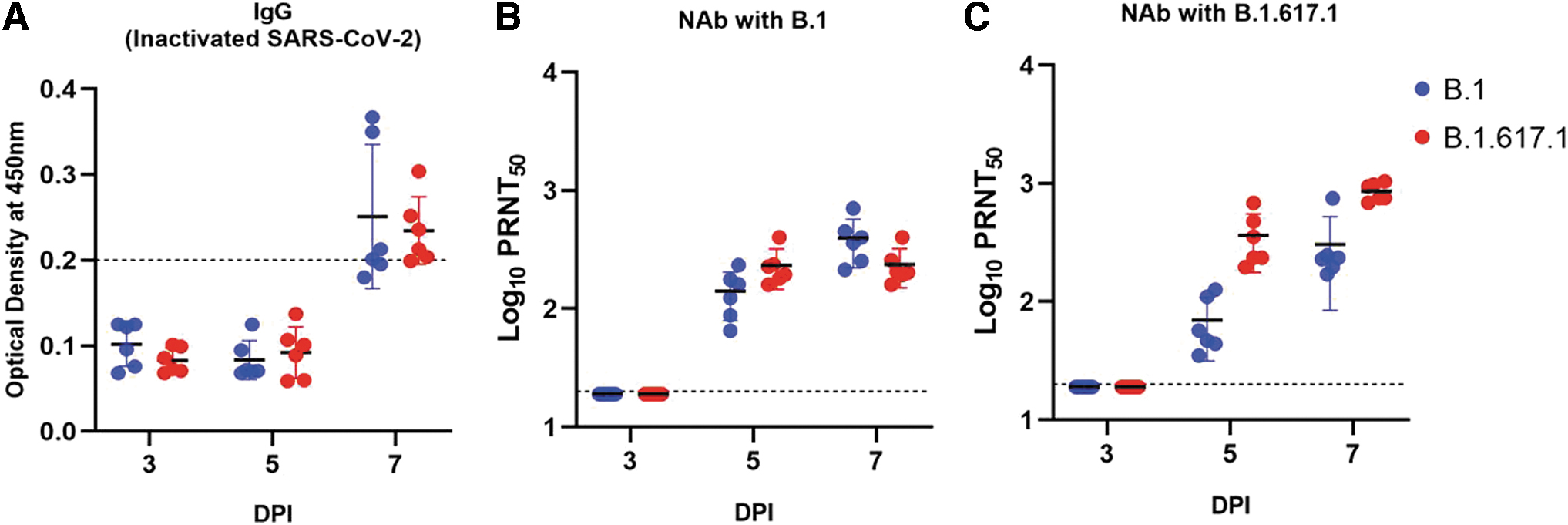
Anti-SARS-CoV-2 antibody response:
Kappa variant induced more severe lung lesions in hamsters
Gross examination of lung specimens showed pronounced congestion and hemorrhages on 5 and 7 DPI in the case of the Kappa compared with the B.1 isolate (Supplementary Fig. S1). The lung lesions in hamsters infected with the B.1 variant ranged from minimal to mild, whereas the Kappa variant developed moderate lesions with significantly high lung histopathological score on 3 (p = 0.0498), 5 (p = 0.0022), and 7 (p = 0.0346) DPI (Fig. 4). In the case of the B.1 variant, the lung lesions observed were minimal to mild, which included inflammatory cell infiltration, focal consolidation, and mild congestion (Fig. 5A–C). The pronounced changes (moderate to severe) with mononuclear infiltration in the alveolar interstitial space, interstitial septal thickening, consolidation, and pneumocyte hyperplasia were observed with Kappa variant consistently till 7 DPI (Fig. 5D–G).

Cumulative histopathological score in hamsters infected with Kappa and B.1 variant. The statistical significance between the SARS-CoV-2 variants B.1 and Kappa was assessed using the two-tailed Mann–Whitney test between two groups; p values of <0.05 were considered to be statistically significant. Mean and standard deviation is depicted for six animals in each group.
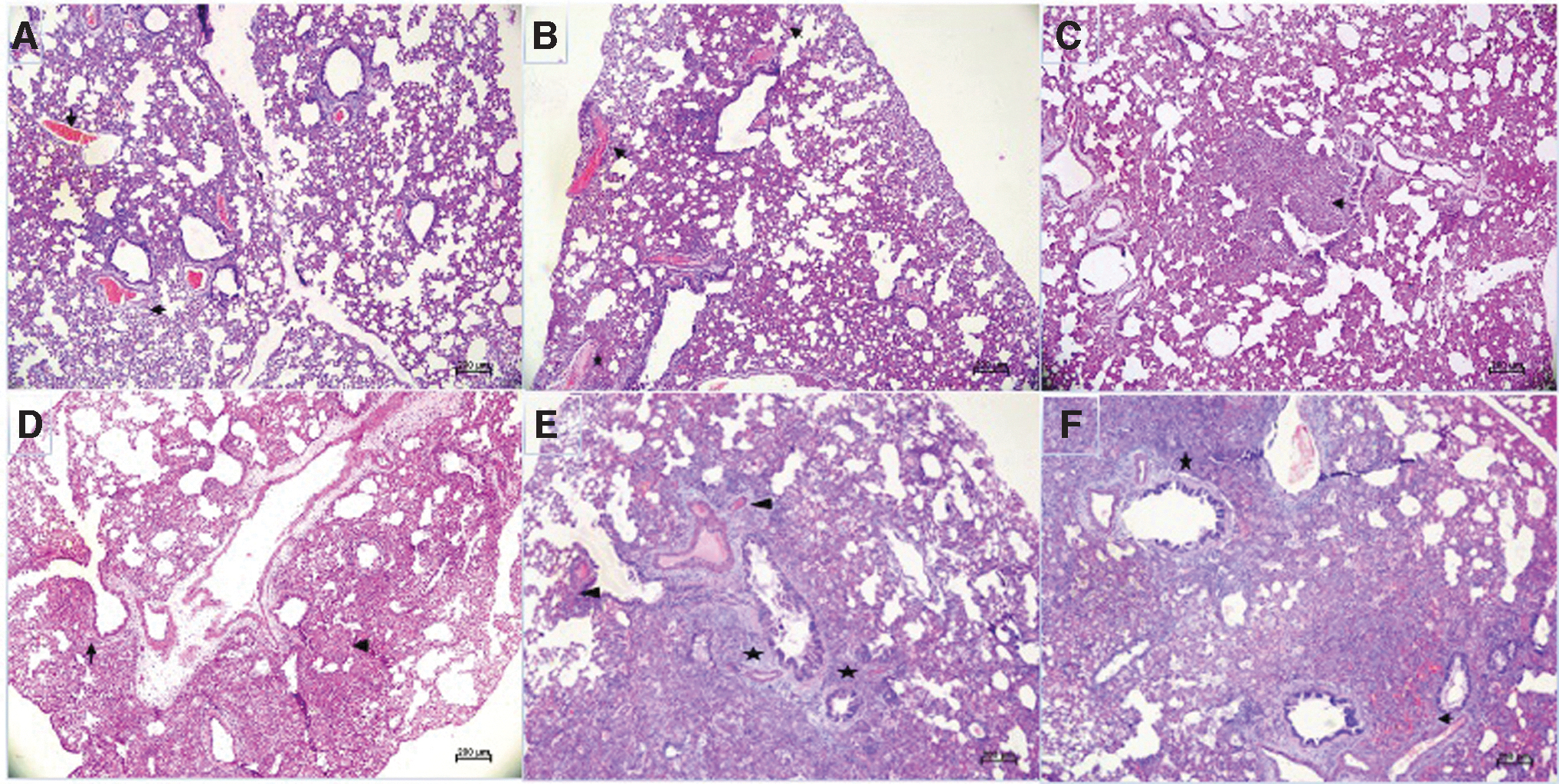
Histopathological changes in hamster's lungs infected with SARS-CoV-2 variants: Lung section of hamsters infected with B.1 showing
Discussion
The emergence of B.1.617 lineage of SARS-CoV-2 has raised concerns among public health experts. The massive surge of cases in India necessitated the study of this variant in the hamster model. Using the Kappa variant virus isolate, we demonstrated the pathogenic potential of this variant in the hamster model in comparison to B.1. The increased severity of Kappa variant infection in hamsters was demonstrated by the higher bodyweight reduction and more severe lung histopathological changes compared to that of B.1. The increased receptor-binding affinity attributed to the characteristic mutations of the Kappa variant isolate might have contributed to the disease severity.
Although the B.1 variant is known to enhance the virus infectivity and transmissibility, it has been reported not to cause severe disease in humans (Korber et al. 2020, Lorenzo-Redondo et al. 2020). Hou et al. (2020) have also demonstrated that the B.1 does not enhance the pathogenicity in hamster models. Besides this, Abdelnabi et al. reported a very efficient infection of B.1.1.7 and B.1.351 variants in the respiratory tract of hamsters (Abdelnabi et al. 2021). Increased cell entry efficiency, a key determinant of viral infectivity has been reported with the Kappa variant in comparison to the wild type Wuhan SARS-CoV-2 (Mlcochova et al. 2021). This could have contributed to the increased lung pathology observed in hamsters.
The protective efficacy of the immunity generated from an initial SARS-CoV-2 infection against subsequent reinfection is puzzling. Studies in animal models have shown that prior infection results in protective immunity (Chandrasekhar et al. 2020). The effect of emerging variants on the ongoing immunization program practiced worldwide needs to be studied in a timely manner. In the present study, we have evaluated the neutralization efficacy of the sera of hamsters infected with the Kappa variant against B.1 variant and vice versa and found NAb response. But the neutralization titer was found reduced in case of the Kappa variant-infected hamster sera against that of B.1. Reinfection studies need to be performed to understand the extent of the protection by these neutralizing antibodies in decreasing the viral RNA load and severity of the disease.
The earlier detection of antibodies by neutralization test compared to the IgG antibody ELISA could be due to the greater sensitivity of the former. We recently reported that sera from Covaxin (whole virion inactivated COVID-19 vaccine) recipients and recovered cases of COVID-19 produced robust neutralizing antibodies against the B.1.617 variant (Yadav et al. 2022a). Studies have shown resistance to neutralization of the Kappa variant by convalescent sera of COVID-19-infected and Moderna and Pfizer vaccines (Edara et al. 2021, Liu et al. 2021). Serum neutralization studies of Covishield (ChAdOx1 nCoV-19 Corona virus vaccine) vaccines with the Kappa variant also have shown NAb response (Yadav et al. 2022b).
Overall in this study, the increased pathogenicity of the Kappa variant lineage was observed in Syrian hamster model with greater loss of body weight and more severe lung lesions in comparison to the B.1 lineage isolate.
Footnotes
Acknowledgment
Authors gratefully acknowledge the team members of Maximum Containment Facility, ICMR-NIV, Pune, including Mr. Prasad Sarkale, Mr. Shreekant Bardkar, Ms. Pranita Gawande, Mrs. Ashwini Waghmare, Ms. Kaumudi Kalele, Ms. Jyoti Yemul, and Ms. Manisha Dhudhmal for providing excellent technical support.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This work was supported by the Indian Council of Medical Research (ICMR), New Delhi at ICMR-National Institute of Virology, Pune, under intramural funding of “COVID-19”.
Supplementary Material
Supplementary Figure S1
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.