
Abstract
Range expansion of the vector tick species, Ixodes scapularis, has been detected in Ontario over the last two decades. This has led to elevated risk of exposure to Borrelia burgdorferi, the bacterium that causes Lyme disease. Previous research using passive surveillance data suggests that I. scapularis populations establish before the establishment of B. burgdorferi transmission cycles, with a delay of ∼5 years. The objectives of this research were to examine spatial and temporal patterns of I. scapularis and its pathogens from 2017 to 2019 in southwestern, eastern, and central Ontario, and to explore patterns of B. burgdorferi invasion. Over the 3-year study period, drag sampling was conducted at 48 sites across Ontario. I. scapularis ticks were tested for B. burgdorferi, Borrelia miyamotoi, Anaplasma phagocytophilum, and Babesia species, including Babesia microti and Babesia odocoilei, and Powassan virus. I. scapularis was detected at 30 sites overall, 22 of which had no history of previous tick detection. B. burgdorferi was detected at nine sites, eight of which tested positive for the first time during this study and five of which had B. burgdorferi detected concurrently with initial tick detection. Tick and pathogen hotspots were identified in eastern Ontario in 2017 and 2018, respectively. These findings provide additional evidence on the range expansion and population establishment of I. scapularis in Ontario and help generate hypotheses on the invasion of B. burgdorferi in Ontario. Ongoing public health surveillance is critical to monitor changes in I. scapularis and its pathogens in Ontario.
Introduction
Lyme disease is the most common vector-borne disease in North America (Hamer et al. 2010, Kilpatrick et al. 2017). In eastern and central North America, it is primarily caused by Borrelia burgdorferi sensu stricto and transmitted by the blacklegged tick, Ixodes scapularis (Burgdorfer et al. 1982, Ogden et al. 2009). In Canada, reported human Lyme disease cases have increased from 144 cases in 2009 to 2636 cases in 2019 (Public Health Agency of Canada 2021). Notable changes have also been documented in canine populations as well, with a 144% increase in seroprevalence from 2008 to 2015 (Evason et al. 2019).
The increased incidence of this emerging infectious disease in Canada is associated with range expansion of I. scapularis (Ogden et al. 2009, Nelder et al. 2018). Within the last 30 years, a northward expansion of I. scapularis has been identified in eastern and central Canada (Leighton et al. 2012, Ogden et al. 2015, Clow et al. 2017a). Ontario, specifically, has seen an increase in I. scapularis populations in areas that had little or no evidence of tick establishment decades ago (Ogden et al. 2010, Public Health Ontario 2021). Passive tick surveillance data have identified the presence of I. scapularis throughout southern Ontario from 2008 to 2012 (Nelder et al. 2014). Active tick surveillance has confirmed the continued range expansion, particularly in eastern Ontario (Nelder et al. 2014, Clow et al. 2017a).
The spread of I. scapularis can be attributed to several factors, including climate change and wildlife host movement (Ogden et al. 2008a, Clow et al. 2017b, Bouchard et al. 2019). Climate change has led to prolonged periods of warmer temperatures in more northern areas throughout the year, which has generated suitable conditions for I. scapularis survival and reproduction (Bouchard et al. 2019). Migratory birds regularly introduce I. scapularis ticks into these northern habitats, providing a seed for population expansion (Ogden et al. 2013). As temperatures increase over time, migratory birds may venture further north in shorter amounts of time, thus reinforcing the process (Ogden et al. 2008a, 2013).
In addition to B. burgdorferi, Anaplasma phagocytophilum, Babesia microti, Powassan virus (deer tick virus lineage), and Borrelia miyamotoi have been identified in I. scapularis ticks in Ontario (Nelder et al. 2016). The mechanisms driving I. scapularis expansion and pathogen invasion is an area of ongoing research; tick-first, coinvasion, and spirochete-first hypotheses have been proposed (Ogden et al. 2008b, Hamer et al. 2010). The “tick-first” or host-mediated invasion scenario involves uninfected I. scapularis ticks being introduced to an area by longer distance movement on hosts, followed by a delayed secondary invasion of a pathogen. In the coinvasion or dual-invasion theory, I. scapularis ticks infected with the pathogen are introduced via hosts simultaneously, allowing for the establishment of both the tick and the pathogen. The “spirochete-first” scenario occurs when a pathogen is maintained in a habitat through cryptic (i.e., nonhuman biting) vectors before I. scapularis establishment (Hamer et al. 2010).
In Lower Michigan, all three processes were detected in different geographic areas for B. burgdorferi invasion (Hamer et al. 2010). Previous analyses of passive surveillance data from eastern Canada illustrated a delay of 5 years for B. burgdorferi establishment after tick establishment, suggesting that a “tick-first” scenario is the driving mechanism (Ogden et al. 2013). However, preliminary field sampling data have illustrated that different patterns may exist and ongoing field-based research is required to continue to investigate these processes in Ontario (Clow et al. 2017a).
Given that Lyme disease is a threat to public and animal health, ongoing research to investigate expansion patterns and the underlying mechanisms of invasion of both the tick and pathogen is crucial. The objectives of this study are to assess the spatial and temporal patterns of I. scapularis and the associated pathogens in southwestern, eastern, and central Ontario from 2017 to 2019, and to explore the patterns of invasion of B. burgdorferi.
Methods
Site selection
Field sampling for ticks previously conducted by Clow et al. (2016, 2017a, 2017b) were used as a baseline for site selection for this study. This study was chosen as a baseline to build a longer temporal sequence. Previously visited and new sites were selected if I. scapularis had been detected but not B. burgdorferi, or if the site was located within a 100 km buffer around the significant spatial cluster for I. scapularis identified by Clow et al. (2016). Selected sites also had to have suitable habitats for I. scapularis (e.g., forest cover) and be accessible for the field team. Additional sites were added in 2018 and 2019 to capture ongoing range expansion of ticks.
All study sites were located within the region of southern Ontario covering three subregions: southwestern, central, and eastern (Government of Ontario 2019). Central Ontario encompasses the Greater Toronto Area and areas north toward Lake Simcoe and Georgian Bay. Southwestern Ontario includes all areas west of Central Ontario and eastern Ontario includes all areas east of Central Ontario. The study region is within a temperate climate zone and spans the deciduous biome (southwestern Ontario) and the Great Lakes-St. Lawrence forest region that comprised a mix of coniferous and deciduous trees (central and eastern Ontario) (Government of Ontario 2021).
Field sampling
Tick dragging was performed at 38, 45, and 46 sites in 2017, 2018, and 2019, respectively. All sites were visited at least once on dry days from May to November, capturing the time of year, in which larval, nymphal, and adult ticks are most active (Lindsay et al. 1995, 1999b, Clow et al. 2018). Longitude and latitude values were measured at each site visit using a Global Positioning System (GPS). To collect ticks, a 1 m2 white flannel cloth was dragged over the forest floor and vegetation at each site for three-person hours. Every 3 min, the timer was stopped so the cloth could be examined for ticks. Larval, nymphal, and adult ticks were removed and stored in 70% ethanol.
Laboratory analysis
Ticks were sent to the National Microbiology Laboratory (NML) of the Public Health Agency of Canada (PHAC) in Winnipeg, Manitoba, and identified to species using standard identification keys (Lindquist et al. 2016). Adult and nymphal I. scapularis were tested for Borrelia spp. including B. burgdorferi, B. miyamotoi and Borrelia kurtenbachii, A. phagocytophilum, and Babesia spp. including B. microti and Babesia odocoilei using PCR. RNA was extracted using commercial kits as per manufacturer's protocol (QIAGEN RNeasy Kit, Toronto, Canada) (Ogden et al. 2006, Dibernardo et al. 2014). A specific multiplex real-time PCR assay was used to simultaneously detect Borrelia spp. and A. phagocytophilum using 23S ribosomal RNA (rRNA) and msp2 genes as amplification targets, respectively (Courtney et al. 2004).
Samples positive for Borrelia spp. were subsequently tested by a duplex confirmatory ospA and flaB real-time PCR assay to differentiate and confirm B. burgdorferi and B. miyamotoi (Tokarz et al. 2017). Positive A. phagocytophilum samples were tested again by the same assay. The detection of Babesia spp. was conducted using a triplex assay targeting the 18S (rRNA) gene of Babesia spp., B. microti, and B. odocoilei. Samples positive for B. microti in the screening assay were confirmed using primers and probes specific for the CCTη gene of B. microti (Nakajima et al. 2009), while samples positive for Babesia spp. and B. odocoilei in the triplex were reported as B. odocoilei. Detection of Powassan virus was carried out using a real-time RT-PCR assay targeting the NS5 region of the Powassan virus genome (Smith et al. 2018).
Statistical analysis
A purely spatial retrospective Bernoulli probability model was used to identify spatial clusters of high prevalence of sites with (1) I. scapularis and (2) B. burgdorferi from 2017 to 2019 using SaTScan version 9.6 (Martin Kulldorff and Information Management Services, Inc., Boston, MA). A flexible scanning window was set at a maximum spatial cluster of 50% of the population at risk. Monte Carlo replications were set to 999. Gini indexes were reported if statistically significant secondary clusters with no geographical overlap were present (Kulldorff 2015). A significance level of α = 0.05 was used for identifying statistically significant spatial clusters. Spatial data was projected using ArcMap version 10.7.1 (Esri, Redlands, CA).
Mixed univariable exact logistic regression was performed to assess the relationship between the outcome, B. burgdorferi detection, and the main effect, years of tick presence until B. burgdorferi detection. For this portion of the analysis, sites were only eligible for inclusion if they had at least 1 year of baseline data where no I. scapularis ticks were detected. Additional years of field sampling data for sites that had previously been visited by Clow et al. (2016, 2017a, 2017b) in 2014, 2015, or 2016 were added to enhance our longitudinal dataset. Each observation was 1 year of data from one site. For the purposes of hypothesis exploration, B. burgdorferi detection was defined as the detection of at least one tick with B. burgdorferi at a site, with no baseline threshold for infection prevalence or requirement for subsequent detection the next year.
The main effect was categorized into “concurrent” to describe sites with concurrent initial detection of tick and pathogen, “greater than one” to describe sites with a delay between tick and pathogen detection, and “future.” Sites that had yet to have B. burgdorferi detected had an additional observation added and categorized as “future” to represent the ongoing temporal process of invasion. The site identification number was included as a random effect to account for multiple observations per site (i.e., each year of data). Exact logistic regression was chosen as some outcome and predictor combinations had <5 observations. A significance level of α ≤ 0.05 was used to assess the relationships. Statistical analyses were conducted using STATA version 16.1 (StataCorp).
Results
I. scapularis detection
Over the 3-year study period, 782 ticks of 7 different tick species were collected by drag sampling over 186 site visits (68 site visits in 2017, 61 in 2018, and 57 in 2019, with each site visited between 1 and 4 times per year). I. scapularis was the most common species, comprising 94.4% (n = 305) of samples in 2017, 96.0% (n = 313) in 2018, and 94.9% (n = 130) in 2019 (Table 1). I. scapularis was detected at 30 sites overall, 22 of which did not have previous I. scapularis presence.
Number of Ixodes scapularis and Prevalence of Borrelia burgdorferi in I. scapularis Collected by Tick Dragging at Sites in 2017, 2018, and 2019
CI, confidence interval.
I. scapularis was present at 12 of 38 sites in 2017 (31.6%), 23 of 45 sites in 2018 (51.5%), and 21 of 46 sites in 2019 (45.7%) (Table 1 and Fig. 1). Only a small number of other tick species were collected: 24 Dermacentor variabilis, 7 Haemaphysalis leporispalustris, 3 Ixodes cookei, 2 Ixodes dentatus, 1 Ixodes muris, and 1 Amblyomma americanum. Most sites with >1 tick species were composed of I. scapularis and D. variabilis. The detection of H. leporispalustris or I. cookei was also noted at four sites with I. scapularis and/or D. variabilis.
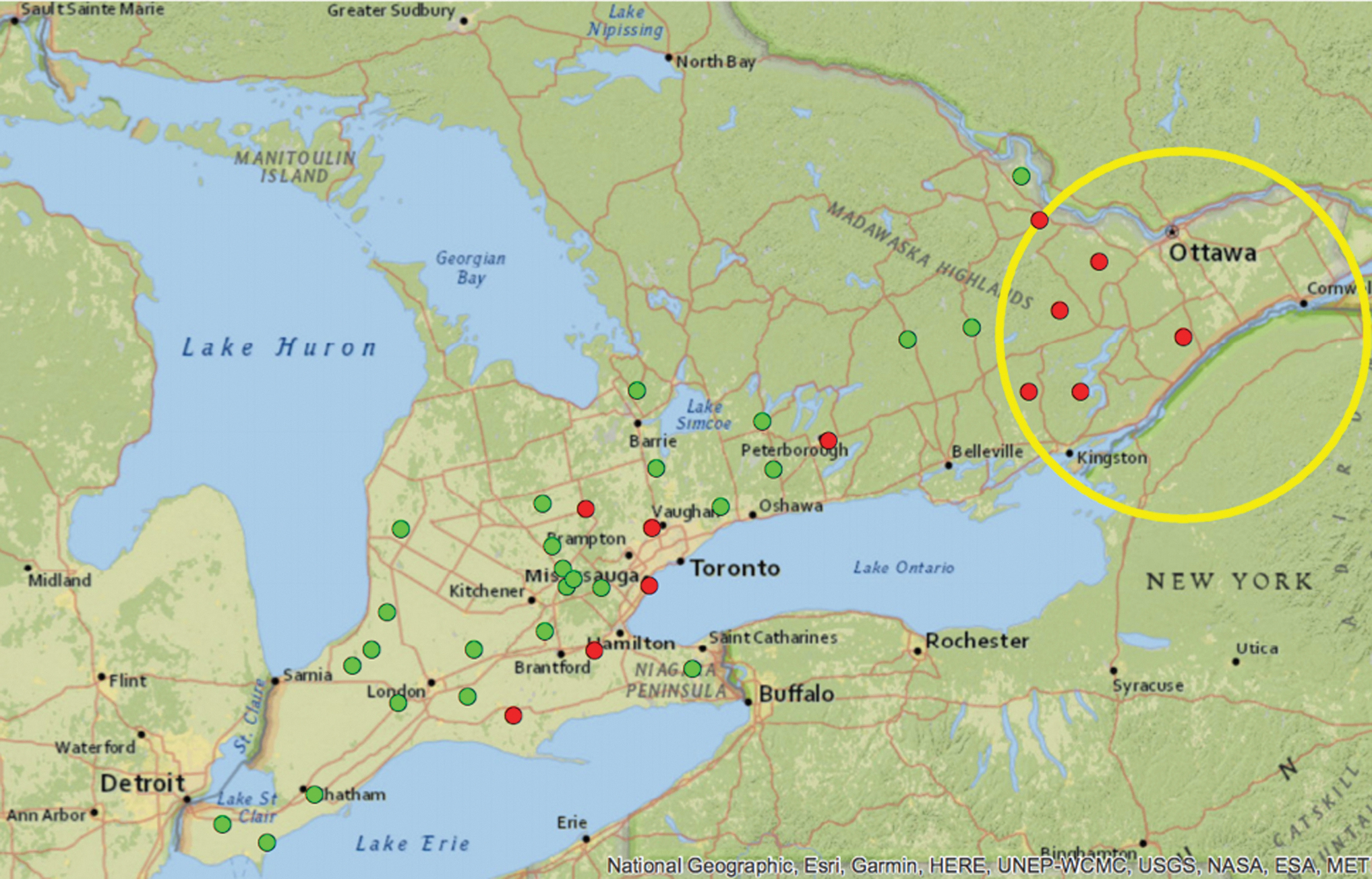
A spatial cluster of high prevalence of sites with Ixodes scapularis was detected in 2017 (yellow ring). I. scapularis-positive sites (red circles) and I. scapularis-negative sites (green circles) are illustrated on the map.
Pathogen detection
Six pathogens were detected across all sampled sites from 2017 to 2019. B. burgdorferi was the most commonly detected pathogen throughout the study period, found in 84 ticks at 9 sites overall (median: 2, range: 1–40). Eight sites tested positive for the first time (e.g., at least one tick from the site tested positive). Six sites were positive for B. burgdorferi in 2017, four sites in 2018, and four sites in 2019 (Table 1 and Fig. 2).

A spatial cluster showing high prevalence of sites with Borrelia burgdorferi-infected Ixodes scapularis ticks was detected in 2018 (yellow ring). B. burgdorferi-and I. scapularis-positive sites (red circles), B. burgdorferi-negative and I. scapularis-positive sites (orange circles), and B. burgdorferi-and I. scapularis-negative sites (green circles) are illustrated on the map.
Other pathogens were detected in I. scapularis, including 11 ticks with B. odocoilei, 4 with A. phagocytophilum, 4 with B. miyamotoi, 1 with B. microti, and 1 with B. kurtenbachii. Coinfection was detected in four ticks in 2019. Coinfections were as follows: one male I. scapularis tick with B. burgdorferi and B. odocoilei, one female I. scapularis tick with B. burgdorferi and A. phagocytophilum, one I. scapularis nymph with B. burgdorferi and B. odocoilei, and one I. muris tick coinfected with B. microti and B. kurtenbachii.
Statistical analysis
A spatial cluster of sites with I. scapularis was identified in eastern Ontario in 2017, which included six I. scapularis-positive sites (Fig. 1). A spatial cluster of sites with B. burgdorferi was detected in eastern Ontario in 2018, which was composed of three B. burgdorferi-positive sites (Fig. 2). No significant spatial clusters were identified for high prevalence of sites with I. scapularis in 2018 and 2019 or B. burgdorferi-infected I. scapularis ticks in 2017 and 2019.
Out of the 49 sites visited from 2017 to 2019, 15 had at least 1 year of baseline data where no I. scapularis were detected and were therefore eligible for inclusion in the exploratory univariable analysis assessing the years of I. scapularis presence until B. burgdorferi detection (Table 2). At least 1 tick was positive for B. burgdorferi at 5 of the 15 sites (Table 3). B. burgdorferi was detected concurrently with initial tick detection at four sites and in the fourth year of tick detection at one site (Table 3).
Exploratory Mixed Univariable Exact Logistic Regression Assessing the Potential Associations with Borrelia burgdorferi Detection (Outcome) with the Number of Years from Tick Detection to B. Burgdorferi Detection (Main Effect)
An additional observation was added for sites where I. scapularis was detected but not B. burgdorferi to represent an ongoing temporal process.
N/A, insufficient data to retrieve confidence interval.
Infection Prevalence (%) of Borrelia burgdorferi at 15 Sites Where Nymphal and Adult Ixodes scapularis Were Detected (95% Confidence Interval)
n = Number of I. scapularis tested for B. burgdorferi presence. Sites not sampled are indicated as (—).
Of the four sites with concurrent tick detection and B. burgdorferi detection, two sites were in central Ontario, one in eastern Ontario, and one in southwestern Ontario. The site observed to have 4 years between initial I. scapularis detection and B. burgdorferi detection was in eastern Ontario (Fig. 3). No statistically significant relationships were detected between the outcome, B. burgdorferi detection, and the main effect, years of I. scapularis detection before B. burgdorferi detection.
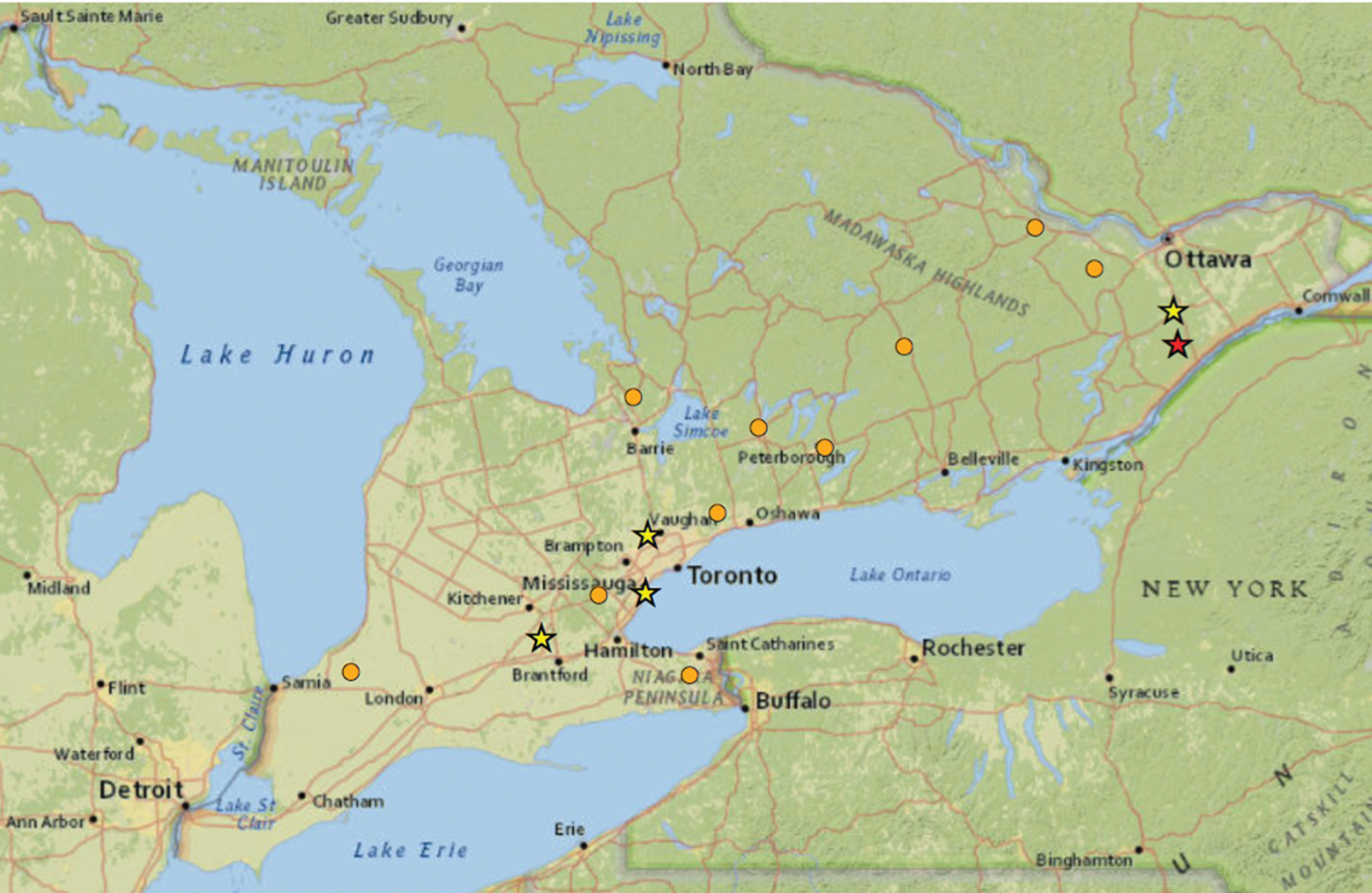
Borrelia burgdorferi was detected at five sites after I. scapularis detection from 2014 to 2019 (stars). Sites are classified into three categories: B. burgdorferi detection in the fourth year of I. scapularis tick detection (red star), B. burgdorferi detection concurrent with I. scapularis tick detection (yellow stars), and I. scapularis present with no B. burgdorferi (orange circles) in 2014–2019.
Discussion
Additional occurrences of I. scapularis were detected each year of the study across several areas of the province, which adds to our evidence base illustrating range expansion of this tick species in Ontario. Most newly detected sites were north of Lake Ontario and the St. Lawrence River where we detected a significant spatial cluster (also referred to as a “hot spot”) for I. scapularis. There was also notable expansion evident in the region north of the Greater Toronto Area, toward Lake Simcoe. Our findings build upon previous field sampling efforts that have documented ongoing range expansion and determined that eastern Ontario was a “hot spot” for I. scapularis detection (Clow et al. 2016, 2017a). In addition, our results closely mirror risk maps generated to predict I. scapularis distribution in 2020 (Ogden et al. 2008b, Leighton et al. 2012).
Several environmental factors need to be considered when evaluating these findings. Suitable habitats and hosts are critical in the expansion of I. scapularis and their associated pathogens (Ogden et al. 2005 2008b). As of 2011, the land north of Lake Ontario and the St. Lawrence River comprised 17% forested habitats, which provides the appropriate habitat and hosts for I. scapularis establishment (Wadkins 2011). During our study, I. scapularis was detected in most sampled sites in this area. In contrast, the area north of Lake Erie, where we detected minimal range expansion, is predominately agricultural land and thus has fewer areas with the necessary wooded habitat to support I. scapularis (Barker et al. 1992, Nelder et al. 2014).
However, we still detected I. scapularis in focal areas of suitable habitat in this region, and we know from previous research that small, suitable land fragments are important to consider in the ecology of I. scapularis given that nymphal ticks can be found at high densities in these areas (VanAcker et al. 2019).
B. burgdorferi was detected in at least one tick from sites in southwestern, central, and eastern Ontario, with eastern Ontario again a “hot spot.” Migratory birds have been deemed a primary method of tick introduction into Canada (Ogden et al. 2008a). The reservoir competency of many migratory bird species is questionable, and nymphal ticks that fed on migratory birds have shown to have low infection prevalence (∼10%) (Ogden et al. 2008a). With B. burgdorferi-infected larval blacklegged ticks rarely introduced, it takes time to establish a sustained transmission cycle between local small mammal or avian reservoir hosts and the establishing tick populations. At the sites where we detected B. burgdorferi for the first time, the infection prevalence was low, which could suggest adventitial introduction via migratory birds and limited local transmission.
Moreover, at two sites, B. burgdorferi was not detected in the following year of sampling, thus further suggesting adventitial introduction. That being said, given that tick dragging has low sensitivity and thus has a high false negative rate, the concurrent identification of numerous ticks and B. burgdorferi at several sites could also represent accelerated emergence of B. burgdorferi compared to the previously hypothesized 5-year lag (Ogden et al. 2013, 2014). As the abundance of established tick populations increases in Ontario, it is possible that both local and long-distance spread are occurring (Ogden et al. 2010, Clow et al. 2017a). In this context, different processes may be facilitating tick and pathogen invasion. We could therefore expect to see variability in the temporal relationship between tick and pathogen emergence.
Watts et al. (2018) modeled I. scapularis and B. burgdorferi invasion on the local scale in the Thousand Islands region. B. burgdorferi-infected mice carrying ticks were shown to facilitate pathogen dispersal on a local level, with dispersal influenced by host and landscape factors (Watts et al. 2018). Our findings remain inconclusive due to the limited timeframe and sampling approach, and therefore, we can only generate hypotheses and recommend future research. Ticks were only collected via drag sampling and no hosts were sampled for B. burgdorferi. Sampling of hosts is invaluable to illustrate local transmission, especially when populations are first establishing, and infection prevalence is low. Additional environmental and host sampling is recommended to build onto our baseline field sampling data, providing more rigorous data on the time between tick establishment and pathogen invasion and aiding in the understanding of the mechanisms driving B. burgdorferi emergence.
The detection of A. americanum, the lonestar tick, in southwestern Ontario in 2019 through tick dragging is worth noting. This tick species has not been identified through active surveillance methods conducted throughout Ontario since active surveillance began in 2008 (Nelder et al. 2019a). However, lonestar ticks are sporadically submitted through passive surveillance programs and thus this tick could be adventitious (Ogden et al. 2008a, Nelder et al. 2019a). Since A. americanum is a vector of several pathogens of public and animal health significance, our finding should be followed up with additional field sampling to assess if this tick is potentially from a recently established reproducing population (Nelder et al. 2019a).
Although the focus of our research was on B. burgdorferi, the additional pathogens detected in I. scapularis are also important to discuss. B. odocoilei is a pathogen of growing concern for animal health and was the most detected pathogen in our study after B. burgdorferi. This tick-borne parasite predominately affects members of the cervid family, and clinical symptoms include fever, lethargy, hemoglobinuria, and sudden death (Mathieu et al. 2018, Milnes et al. 2019). B. microti and A. phagocytophilum are rarely detected in I. scapularis populations in the province, and our findings continue to illustrate this low-infection prevalence (Werden et al. 2015).
That being said, the prevalence of this pathogen in blacklegged ticks in the eastern United States has been increasing and thus could become an increasing health risk (Nelder et al. 2016, 2019b). Specifically, eastern Ontario has been highlighted as an area of A. phagocytophilum emergence, and in our study, A. phagocytophilum was only detected at sites in eastern Ontario (Nelder et al. 2019b).
Our study has several limitations that should be acknowledged. First, we only collected data for 3 years and this timeframe is not sufficient to extensively explore the invasion process of I. scapularis and B. burgdorferi. Also, the number of visits and season of visit varied over the study period. In 2017 and 2018, over half of the sites were visited two or more times; but in 2019, each site was only visited once. In addition, fall sampling occurred only in 2017. This was predominately due to funding and available human resources. In general, when sites are visited more often, there are more opportunities to detect I. scapularis and thus B. burgdorferi.
Fall sampling may also lead to higher likelihood of B. burgdorferi detection as it is the peak activity time for adult I. scapularis (Lindsay et al. 1999a) and this life stage is more likely to be infected with B. burgdorferi. To avoid inaccurately presenting the data, we did not use tick abundance or infection prevalence as an outcome for any analyses. Tick dragging has low sensitivity (∼50%), particularly when tick populations are just establishing and thus we may have missed detecting I. scapularis at some sites if tick densities were low (Ogden et al. 2014). Furthermore, the sample size for the baseline field sampling data was small, which limited the power in our analyses. To build on our research, additional years of targeted and rigorous field sampling coupling environmental data with host data are required.
Conclusion
Our data have identified areas in Ontario known to have I. scapularis and B. burgdorferi, while highlighting regions of the province that could be at risk for tick and pathogen invasion. We have also detected B. burgdorferi presence that was subsequently explored using different hypotheses for B. burgdorferi invasion, and thus set the stage for future research. These findings can be used to inform the public about current risks and encourage the uptake of primary prevention measures to reduce I. scapularis exposure to humans and animals. In addition, they can help public health officials determine where to target surveillance efforts and inform health care professionals on the increasing risk of tick-borne pathogens.
Lyme disease is an ongoing and increasing threat to human and animal health in Canada, and a collaborative approach should be considered for future research to enable a holistic understanding of the human, animal, and environmental factors driving the ongoing expansion of the tick and its pathogens (Ogden et al. 2008b, Nelder et al. 2014).
Footnotes
Acknowledgments
We thank the dedicated summer student tick dragging team of Genevieve Lumsden, Rachel Finer, Tasmin Duncan, Courtney Winkels, Dimo Kecheliev, and Anthony Maraj.
Authors' Contributions
Study design was completed by C.M.J., K.M.C., and J.K.K. Laboratory analyses were completed by L.R.L. and A.D. Spatial and statistical analyses were completed by E.L.R. with input from C.M.J., J.K.K., C.R., L.R.L., and K.M.C. Article writing was led by E.L.R. with input from all authors. All authors approved the final version of the article.
Author Disclosure Statement
The authors disclose no conflicts of interest.
Funding Information
This research was generously supported by the PHAC. E.L.R. was supported by a graduate student stipend from the Department of Population Medicine, University of Guelph.