
Abstract
Background:
The spread of emerging tick-borne pathogens has steadily increased in Canada with the widespread establishment of tick vectors and vertebrate hosts. At present, Borrelia burgdorferi, the bacterium causing Lyme disease, is the most common tick-borne pathogen in Canada and primarily transmitted by Ixodes scapularis. A low prevalence of other emerging tick-borne pathogens, such as Anaplasma phagocytophilum, Babesia species, Borrelia miyamotoi, and Francisella tularensis have also been detected through surveillance efforts in Canada. Although Rickettsia rickettsii has been historically detected in Haemaphysalis leporispalustris in Canada, the current prevalence and geographic extent of this pathogen is unknown.
Material and Methods:
In this study, we assessed the presence and prevalence of several emerging tick-borne pathogens in ticks and hosts collected through tick dragging and small mammal trapping in Central Canada.
Results:
Nested PCR testing detected three pathogen species in ticks, with Babesia odocoilei and B. burgdorferi in I. scapularis in addition to R. rickettsii in H. leporispalustris. Three pathogen species were detected in small mammals by nested PCR including B. odocoilei in Blarina brevicauda, Babesia microti in Peromyscus leucopus, and a Hepatozoon species in P. leucopus and Peromyscus maniculatus. B. burgdorferi and Babesia species were the pathogens most often detected in our samples, suggesting they are widely distributed across Central Canada. We also detected B. odocoilei and R. rickettsii beyond their known geographic distribution.
Conclusions:
Our results provide evidence that emerging tick-borne pathogens may be present outside defined risk areas identified by current surveillance efforts in Canada. As a result, emerging tick-borne pathogens introduced by the dispersal of infected ticks by migratory birds or maintained by hosts and vectors through cryptic transmission cycles may go undetected. More comprehensive testing including all tick life stages and additional tick-borne pathogens will help detect the spread and potential risk of emerging or re-emerging tick-borne pathogens for human and wildlife populations throughout Canada.
Introduction
Emerging tick-borne pathogens have increased in prevalence in Canada with climate warming, habitat fragmentation, and changes in the abundances and distributions of tick and host populations (Bouchard et al, 2019; Leo et al, 2016; Ogden and Lindsay, 2016). Pathogen transmission cycles are sustained by contact between ticks and their hosts (Radolf et al, 2012). Generalist tick species feed on a wide variety of hosts, including Ixodes scapularis feeding on mammals, birds, and humans and Haemaphysalis leporispalustris feeding on lagomorphs and birds (Keirans et al, 1996; Kollars and Oliver, 2003; Lindquist and Wu, 2016). Although some host specificity may occur depending on local host availability (McCoy et al, 2013), specialist tick species feed on select hosts such as Ixodes banksi and beavers (Lindquist and Wu, 2016).
Birds and small mammals are competent reservoir hosts that successfully feed large numbers of ticks and have greater probabilities of infecting feeding ticks with pathogens (Bouchard et al, 2011; LoGiudice et al, 2003; Ogden et al, 2008; Scott et al, 2019; Zinck and Lloyd, 2022). As a result, emerging tick-borne pathogens may be detected in areas beyond currently defined risk areas in Canada owing to the dispersal of infected ticks by migratory birds and the distributional shifts of host populations (Fiset et al, 2015; Garcia-Elfring et al, 2017; Roy-Dufresne et al, 2013; Simon et al, 2014).
The predominant tick-borne pathogen found in sentinel surveillance efforts in Canada is Borrelia burgdorferi, the bacterium causing Lyme disease (Guillot et al, 2020). Lyme disease prevalence has grown rapidly to become the most common tick-borne disease in North America (Gasmi et al, 2017; Nelder et al, 2018). The first case of another Borrelia species in Canada, Borrelia miyamotoi, was reported in 2013 (Bouchard et al, 2019). These spirochetes are primarily transmitted by I. scapularis and Ixodes pacificus ticks as well as rodents, especially Peromyscus mice (Kulkarni et al, 2015).
Other emerging tick-borne pathogens targeted by current surveillance efforts in Canada are expected to increase in the future, as Ixodidae transmit 40% of documented emerging vector-borne zoonotic diseases globally (Swei et al, 2020). Many pathogens including Anaplasma phagocytophilum and Babesia microti are primarily transmitted by Ixodes ticks and small mammals in Canada (Bouchard et al, 2019; Kulkarni et al, 2015). More recently, Babesia odocoilei has been found in cervids and I. scapularis in Canada (Mathieu et al, 2018; Milnes et al, 2019; Pattullo et al, 2013). In contrast, Francisella tularensis, the bacterium causing tularemia, is transmitted by Dermacentor, Amblyomma, and Haemaphysalis ticks. Tularemia can also spread through contact with infected mammals, infective aerosols, or arthropod bites (Gabriele-Rivet et al, 2016; Petersen et al, 2009).
Rickettsia rickettsii, the bacterium causing Rocky Mountain spotted fever, is vectored by Dermacentor, Haemaphysalis, and Rhipicephalus ticks. In Canada, the historical extent of R. rickettsii has relied on surveys and human cases owing to its rare occurrence (Bouchard et al, 2019; Humphreys and Campbell, 1947; Kulkarni et al, 2015). This bacterium has been found previously in H. leporispalustris ticks and dogs in Ontario, Nova Scotia, and Western Canada (Gary et al, 2006; Leighton et al, 2001; Wood and Artsob, 2012). Although H. leporispalustris does not typically bite humans, this tick species is important for the maintenance of R. rickettsii strains of variable virulence in ecosystems (Freitas et al, 2009; Parker et al, 1951). The current prevalence and geographic extent of R. rickettsii strains in H. leporispalustris ticks in Canada are unknown.
Only two of these emerging tick-borne pathogens are listed as nationally notifiable diseases in Canada: Lyme disease and tularemia (Bouchard et al, 2019). However, current sentinel surveillance efforts in Canada primarily target A. phagocytophilum, Borrelia species, and Babesia species (Guillot et al, 2020; Wilson et al, 2022). As certain emerging tick-borne pathogens are not reportable to public health agencies, it is challenging to assess their degree of establishment and spread across Canada. In this study, we assess the presence and prevalence of several emerging tick-borne pathogens along the northward edge of their range in Ontario and Quebec, Canada. We report recent detections of B. odocoilei and R. rickettsii in southeastern Quebec outside of their known geographic extent based on ongoing surveillance efforts. Our results highlight the need for comprehensive pathogen testing to detect the presence and monitor the spread of emerging tick-borne pathogens throughout Canada.
Materials and Methods
Sixteen forested sites of varying risk for Lyme disease were sampled in July and August 2019 in Central Canada [Institut national de santé publique du Québec, 2018; Ontario Agency for Health Protection and Promotion (Public Health Ontario), 2018]. At each site, tick dragging and small mammal trapping were conducted across three 40 meters by 70 meters grids. Within each grid, a 1 m2 cotton flannel was dragged along four transects over low-lying vegetation and checked every 10 meters. Questing ticks were removed and placed into microvials with 95% ethanol and were later identified to the species using standard taxonomic keys (Egizi et al, 2019; Lindquist and Wu, 2016).
Small mammal trapping was conducted over 3 consecutive nights using 84 Sherman live traps (H.B. Sherman Traps, Inc.). Targeted species included mice (Peromyscus leucopus and Peromyscus maniculatus), shrews (Blarina brevicauda and Sorex cinereus), voles (Microtus pennsylvanicus and Myodes gapperi), and jumping mice (Napaeozapus insignis and Zapus hudsonius). Traps were baited with peanut butter and oatmeal, a water source (apple), and nesting material (cotton ball) in the late afternoon and checked the following morning. Nontargeted species and juveniles of targeted species were immediately released. Target species were killed in the field with isoflurane inhalation overdose followed by cervical dislocation. Owing to serious injuries, one red squirrel (Tamiasciurus hudsonicus) and two hairy-tailed moles (Parascalops breweri) were killed.
The liver of each specimen was dissected and placed into microvials with 95% ethanol. Feeding ticks were removed from the host and placed into microvials with 95% ethanol. All tick and small mammal specimens were accessioned in the collections of the Redpath Museum at McGill University (Montreal, Canada). All procedures were approved by McGill University (AUP No. 2019-8086), the Quebec Ministère des Forêts, de la Faune et des Parcs (SEG permit No. 2019-06-04-008-00-S-F), and the Ontario Ministry of Natural Resources and Forestry (WSCA No. 1093495).
Questing larval ticks were pooled together for testing by grid, with 2–10 larvae per pool. Feeding larval ticks found on the same individual host were pooled together, ranging from 1 to 10 larvae per pool. Questing and feeding nymphs and adults were tested individually. All DNA extractions and PCR were performed by Geneticks, Inc. As in Wills et al (2018), cleaned ticks were cut and homogenized using a microtube pestle in AquaGenomic solution. Samples were incubated in a heat block at 60°C for 45 min, vortexed briefly, and centrifuged for 4 min at 13,300 rpm. The supernatant was transferred to a microvial with 50 μL isopropanol, inverted, and centrifuged as before. After decanting the supernatant, the remaining DNA pellet was rinsed with 50 μL of 70% ethanol and left to air dry for 15 min at room temperature.
This pellet was resuspended with 50 μL of 1 mM Tris pH 8.0 and incubated in a heat block at 60°C for 1 h. DNA from mammal livers was extracted using the Thermo Scientific GeneJET Genomic Purification Kit (Thermo Fisher Scientific). The following modifications were made to the Mammalian Tissue Genomic DNA Purification protocol (Protocol A, 2016): 10 mg of liver was used, the extra centrifugation step 9 was included to remove residual solution, and no additional elution buffer was required after sitting for 5 min before centrifugation. DNA was used directly for PCR and then stored at −20°C.
Nested PCRs to identify Peromyscus specimens to species level were run using species-specific COIII primers following Tessier et al (2004). An initial denaturation time of 5 min was used. PCR products were run on 3% agarose gel, stained with Eco-Stain (Bio Basic, Markham, Canada), and visualized using a blue light transilluminator.
Nested PCRs were performed to target several pathogens in our tick and mammal specimens. The primers and conditions used are described in detail in Supplementary Table S1. All specimens were tested for A. phagocytophilum, Babesia species, and Borrelia species. B. burgdorferi and B. miyamotoi were tested once using the 5S–23S intergenic space (IGS) region. If a band was visible (i.e., positive PCR), two more replicates were conducted to identify false positives. An additional test using the flaB gene was performed to confirm the presence of B. burgdorferi (Supplementary Fig. S1). B. microti and B. odocoilei were targeted once with the 18S ribosomal RNA (rRNA) gene using the mic 494 and odo563 inner primers, respectively (Supplementary Fig. S2). If a band was visible, mic494 and/or odo563 primers were then tested twice more to distinguish false positives. An additional primer set targeting one Babesia species was used to confirm positive testing. The p44 gene was targeted to test for A. phagocytophilum.
Targeted testing of F. tularensis (fdx gene) and R. rickettsii (RRi6 hypothetical protein gene) was conducted for H. leporispalustris specimens, as these pathogens are typically found in this tick species, but not in Ixodes species. Amplified DNA was visualized on a 1.8% agarose gel stained with Eco-Stain (Bio Basic) using a blue light transilluminator. If no band was visualized, the sample was considered negative for that pathogen.
Amplified products were purified before sequencing using a cotton cushion following Sun et al (2012). The fragments were spun for 7 min at 5000 rpm, which were then reamplified using the corresponding inner primers to concentrate the amplicons for sequencing. Sanger DNA sequencing was performed at Bio Basic DNA Sequencing with the forward inner PCR primers.
All sequences were assessed for ambiguous base calls, end-reading errors, and quality scores using the 4Peaks software. Sequences with quality scores <20 were not included in our analyses. A MEGABLAST search using the nucleotide BLAST database (
Results
A total of 644 ticks were collected, with 506 larvae, 136 nymphs, and 2 adults (Table 1). For I. scapularis, we found 382 questing ticks including 255 larvae (29 pools), 126 nymphs, and 1 adult male in addition to 65 feeding ticks comprising 57 larvae (17 pools) and 8 nymphs. For H. leporispalustris, we only found 195 questing ticks at 4 sites in Quebec, which consisted of 194 larvae (22 pools) and 1 nymph. One Ixodes auritulus nymph feeding on one M. gapperi at Rose Hill and one Ixodes marxi adult female feeding on one T. hudsonicus at Saint-Majorique were found.
Total Number and Number of Infected Pools of Ticks at Our Study Sites in Ontario and Quebec, Canada
Larvae were tested together in pools, which were separated by grid if questing and by host if feeding. Nymphs and adults were tested individually. Questing pools are indicated by a “Q” and ticks found feeding on small mammals are indicated by an “F.” Numbers in parentheses represent the number of infected and total pools of ticks. These two infected small mammal individuals harbored different pathogen species than the feeding larvae, with one P. leucopus with a Hepatozoon spp. from New New Age Farm and one P. leucopus with Babesia microti from Henryville.
Infected small mammal host.
For questing I. scapularis, 29 larval pools and an adult male tested negative for all pathogens. However, 27 of 126 questing nymphs (21.42%) tested positive for a pathogen (Fig. 1 and Table 1). Five nymphs were infected with B. odocoilei (3.97%), including one nymph at New New Age Farm, Henryville, and Saint-Valentin, and two nymphs at Saint-Majorique. Similarly, 22 nymphs were infected with B. burgdorferi (17.46%), with 1, 3, and 18 nymphs at New New Age Farm, Henryville, and Saint-Valentin, respectively. For H. leporispalustris, 5 of 22 questing larval pools were infected with R. rickettsii (22.72%), with 4 pools at Coleraine and 1 pool at Frontenac. One uninfected H. leporispalustris nymph was also found at Coleraine.

Pathogens detected in ticks at our study sites in Ontario and Quebec, Canada. Circle size is representative of the abundance of local ticks, where larger circles denote greater tick abundances. No ticks were detected at sites with orange crosses. Circle coloration demonstrates the proportional results of pathogen testing for tick pools, which consisted of larval pools, individual nymphs, and individual adults. The proportion of the circle in light blue represents the tick pools that were negative for pathogen testing. Proportions of tick pools that were positive and harboring Babesia odocoilei (turquoise), Borrelia burgdorferi (light purple), or Rickettsia rickettsii (dark purple) are noted on the map. Study sites include (1) 3 Ridges Farm, (2) New New Age Farm, (3) North Tract—York Regional Forest, (4) Brown Hill Tract—York Regional Forest, (5) Upjohn Nature Reserve, (6) Dyer Memorial Nature Reserve, (7) Rose Hill Nature Reserve, (8) Kirkview Farm, (9) Saint-Polycarpe, (10) Saint-Valentin, (11) Henryville, (12) Lefebvre, (13) Parc du Sanctuaire Saint-Majorique, (14) Serpentine-de-Coleraine Ecological Reserve, (15) Frontenac National Park, (16) Saint-Sylvestre. The land cover map was extracted from provincial rasters of the 2019 annual crop inventories by Agriculture and Agri-Food Canada—Agroclimate (2019).
Feeding ticks were found on 22 small mammals (Table 1). For feeding I. scapularis ticks, 2 of 17 larval pools (11.76%) and 1 of 8 nymphs (12.50%) were infected (Fig. 1 and Table 1). One feeding larva from Henryville was infected with B. odocoilei, whereas its P. leucopus host was infected with B. microti. A larva feeding on an uninfected P. maniculatus from Saint-Majorique and a nymph feeding on an uninfected P. leucopus at Kirkview Farm were infected with B. burgdorferi. The feeding I. auritulus and I. marxi tested negative for all pathogens.
We did not detect similar infection rates in small mammals (Fig. 2 and Table 2). Only 4 of 105 small mammal hosts tested positive for a pathogen (3.8%). One B. brevicauda from Saint-Sylvestre tested positive for B. odocoilei and one P. leucopus from Henryville tested positive for B. microti. Two Peromyscus individuals tested positive for a Hepatozoon species, with one P. leucopus from New New Age Farm and one P. maniculatus from Henryville. We found a feeding larva on both infected P. leucopus, with one uninfected larva from New New Age Farm and one larva infected with B. odocoilei from Henryville.
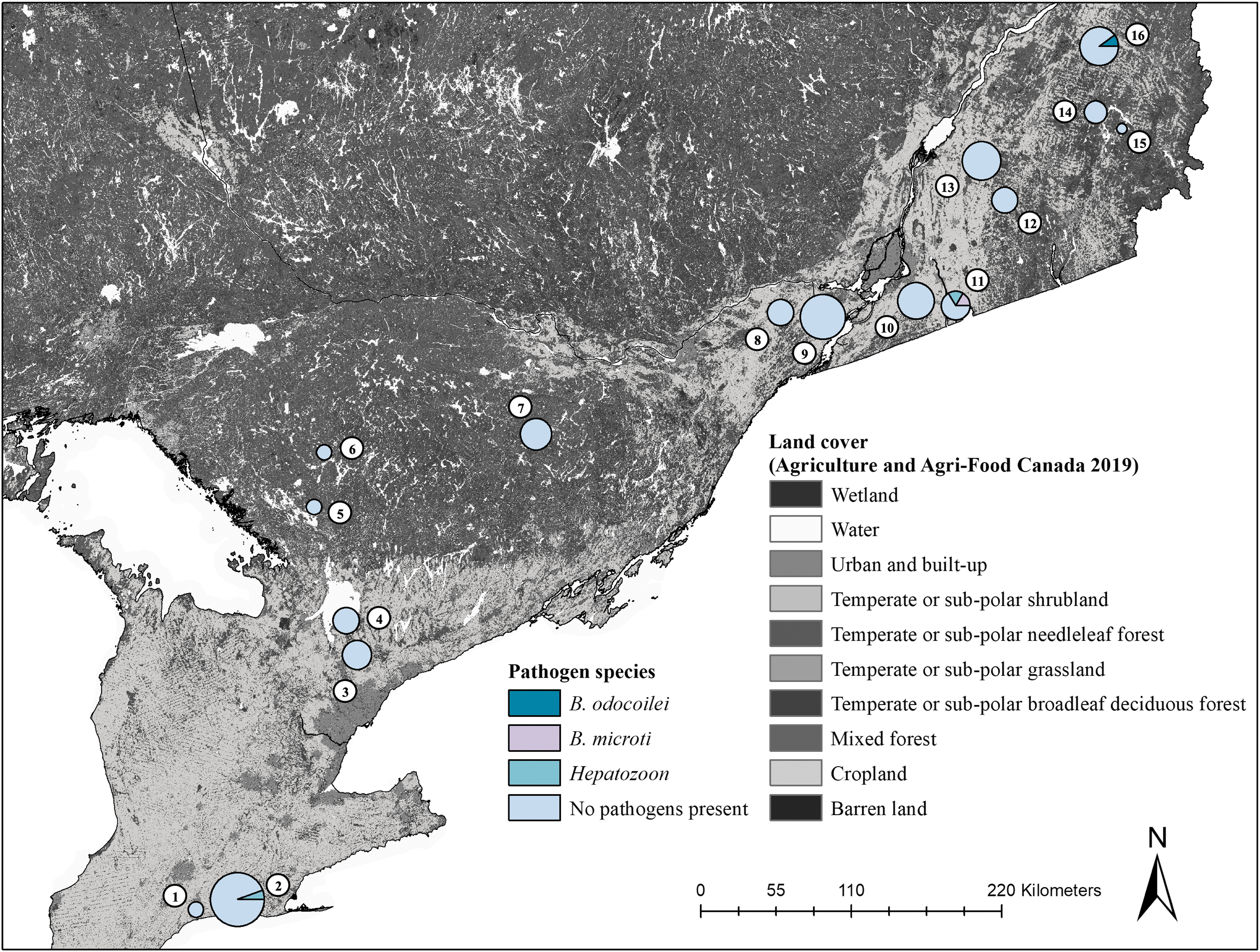
Pathogens detected in small mammals in Ontario and Quebec, Canada. Circle size is representative of the abundance of collected small mammals, where larger circles denote greater abundances of collected small mammal. Circle coloration demonstrates the proportional results of pathogen testing for individual small mammals. The proportion of the circle in light blue represents the collected small mammals that were negative for pathogen testing. Individual small mammals that were positive and harboring Babesia odocoilei (turquoise), Babesia microti (pink), and a Hepatozoon species (teal) are noted on the map. Study sites include (1) 3 Ridges Farm, (2) New New Age Farm, (3) North Tract—York Regional Forest, (4) Brown Hill Tract—York Regional Forest, (5) Upjohn Nature Reserve, (6) Dyer Memorial Nature Reserve, (7) Rose Hill Nature Reserve, (8) Kirkview Farm, (9) Saint-Polycarpe, (10) Saint-Valentin, (11) Henryville, (12) Lefebvre, (13) Parc du Sanctuaire Saint-Majorique, (14) Serpentine-de-Coleraine Ecological Reserve, (15) Frontenac National Park, (16) Saint-Sylvestre. The land cover map was extracted from provincial rasters of the 2019 annual crop inventories by Agriculture and Agri-Food Canada—Agroclimate (2019).
Total Number and Number of Infected Small Mammal Individuals at Our Study Sites in Ontario and Quebec, Canada
Numbers in parentheses represent the number of infected and total small mammal individuals. Only four small mammal specimens were infected across our sites. Two Peromyscus individuals were infected with a Hepatozoon spp., with one P. leucopus from New New Age Farm and one P. maniculatus from Henryville. One P. leucopus individual was also infected with B. microti from Henryville. One B. brevicauda individual from Saint-Sylvestre was infected with Babesia odocoilei.
Discussion
We detected B. odocoilei and R. rickettsii in ticks and small mammals beyond their distributional limits based on current surveillance efforts in southeastern Quebec. No recent detections have been reported for R. rickettsii in Haemaphysalis or Dermacentor ticks in Canada (Dergousoff et al, 2009; Teng et al, 2011; Wood and Artsob, 2012; Wood et al, 2016; Yunik et al, 2015). Historically, R. rickettsii has been identified in H. leporispalustris ticks in Alberta, British Columbia, Ontario, and Nova Scotia, but has not been detected in Quebec (Humphreys and Campbell, 1947; Wood and Artsob, 2012). In Canada, B. odocoilei was first detected through cervid infections (Mathieu et al, 2018; Pattullo et al, 2013). More recently, our study as well as others have detected B. odocoilei in I. scapularis ticks in Central Canada (Milnes et al, 2019; Robinson et al, 2022; Scott and Pesapane, 2021; Scott et al, 2021; Scott et al, 2020). We also report for the first time B. odocoilei being detected in a shrew in Canada.
These pathogens use two modes of transmission in ticks including transovarial transmission from female to larvae and transstadial transmission between immature and adult stages—the latter relying on a bloodmeal taken from a competent host (Freitas et al, 2009; Moore et al, 2018; Roth et al, 2017; Zembsch et al, 2021). The dispersal of ticks infected with Babesia species and R. rickettsii by migratory songbirds has allowed the spread of these pathogens in Canada (Scott et al, 2021; Scott et al, 2020; Scott et al, 2019). Local competent hosts can maintain these pathogens in the environment, but feeding ticks may not become infected owing to low levels of pathogen circulation and variable strain virulence (Freitas et al, 2009; McDade and Newhouse, 1986).
Transovarial transmission can help spread these pathogens without requiring an infected host, although not all offspring may become infected owing to partial transovarial transmission, leading to low larval infection rates (Freitas et al, 2009; Zembsch et al, 2021). Therefore, it is unknown how quickly these emerging or re-emerging pathogens will spread in Canada owing to biotic barriers affecting pathogen transmission.
We also detected a Hepatozoon species in two white-footed mice. This pathogen is vectored by many arthropods including mosquitoes, ticks, mites, fleas, and flies, which are its definitive hosts (Smith, 1996). In Canada, competent intermediate hosts of this parasite include snakes, frogs, and small mammals (Boulianne et al, 2007; Léveillé et al, 2021; Léveillé et al, 2020). This parasite is transmitted to a mouse or another intermediate host through the accidental or intentional ingestion of an arthropod with infective oocytes. Arthropods consist of a large portion of the diet of Peromyscus mice, thereby providing greater opportunities for parasite transmission (Wolff et al, 1985). The parasite will then develop in its intermediate host, where it will be ready to infect future feeding arthropods (Smith, 1996).
The most prevalent pathogen among our samples was B. burgdorferi. Current surveillance efforts in Canada detect a high incidence of B. burgdorferi in ticks and vertebrate hosts, with an increase in positive I. scapularis ticks from 5.9% to 23% in the past decade in Central Canada (Gasmi et al, 2017; Gasmi et al, 2016; Guillot et al, 2020; Kulkarni et al, 2019; Milnes et al, 2019; Nelder et al, 2014; Slatculescu et al, 2020). Lower prevalence of A. phagocytophilum, B. miyamotoi, and B. microti have also been found in I. scapularis ticks through surveillance efforts in Canada (Guillot et al, 2020; O'Brien et al, 2016; Wilson et al, 2022). We only found B. microti in one P. leucopus individual in an endemic region of Quebec.
Surprisingly, no small mammals tested positive for B. burgdorferi. We tested liver tissues, which have been used for B. burgdorferi detection in wild rodents (Zinck and Lloyd, 2022). Higher B. burgdorferi detection rates might have been found if tail, tongue, ear samples (Zawada et al, 2020), or lung tissues (André et al, 2017) were used. A study analyzing liver microbiomes in P. leucopus found a low number of Borrelia sequences with next-generation sequencing, which were undetectable by classic endpoint PCR (André et al, 2017). However, we used longer B. burgdorferi fragments here (∼340 or 605 bp) than in André et al (2017) (142 bp). Therefore, B. burgdorferi may not be detected in some samples because of low concentrations or longer amplicons.
The primary tick-borne pathogens of interest for sentinel surveillance efforts in Canada are A. phagocytophilum, Borrelia species, and Babesia species (Guillot et al, 2020). Typically, these pathogens are tested in nymphs and adults. Larvae are rarely tested for pathogens, as transovarial transmission would be required. However, tick-borne pathogens are expected to increase in the coming decades, as tick and host populations continue to expand into new areas in Canada. Therefore, future surveillance efforts in Canada should focus on two key aspects for more comprehensive emerging tick-borne pathogen testing. First, pathogen testing should be performed on all tick life stages to detect possible transovarial transmission. Second, testing of nontargeted pathogens such as Rickettsia species can help detect pathogens that may be introduced by the dispersal of infected ticks by migratory birds or maintained through cryptic transmission cycles (Hamer et al, 2011). As a result, surveillance efforts would better detect the spread and potential risk of emerging or re-emerging tick-borne pathogens for human and wildlife populations throughout Canada.
Conclusions
Our study identified two emerging tick-borne pathogens, B. odocoilei and R. rickettsii, outside of their known range in Quebec. We also detected cases of B. burgdorferi, B. microti, and Hepatozoon spp. in tick and small mammal specimens in Central Canada. We demonstrate that all tick life stages should be tested comprehensively for pathogens, especially with the increased presence and spread of emerging or re-emerging tick-borne pathogens in Canada.
Footnotes
Acknowledgments
The authors express their appreciation to the landowners and stakeholders who allowed access to their property for field sampling. The authors gratefully thank all their assistants for their invaluable help in the field: Daniella Cross, Connia Ren, Dania Shaban, Ellen Bidulka, Jihane Benbahtane, Christina Provost, and Sébastien Cyr. The authors also thank Robbin Lindsay and Antonia Dibernardo for sharing the protocols used by the National Microbiology Lab for DNA extraction and PCR testing of tick samples.
Authors' Contributions
K.E.C., J.T.K., and V.M. conceived the study. K.E.C. led the methodology, data collection, formal analysis, visualizations, and wrote the article. J.T.K. and V.M. jointly supervised the research, helped design analyses, and contributed to writing.
Author Disclosure Statement
The authors declare no conflict of interest.
Funding Information
This study was funded by Hydro-Québec and Natural Sciences and Engineering Research Council fellowships to K.E.C. J.T.K. is supported by funds from a Natural Sciences and Engineering Research Council Discovery Grant—RGPIN-2017-147544, Natural Sciences and Engineering Research Council Discovery Accelerator Supplement, and the University Research Chair in Macroecology and Conservation at the University of Ottawa. V.M. is supported by a Natural Sciences and Engineering Research Council Discovery Grant—RGPIN-2017-03839.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.