
Abstract
Background:
Leishmaniases are a group of vector-borne zoonotic diseases of public health relevance within the tropical and subtropical regions of the world. The state of Yucatan is a vulnerable and receptive area to localized cutaneous leishmaniasis (LCL) due to its proximity to the high-transmission endemic states of Campeche and Quintana Roo. Autochthonous cases of LCL caused by Leishmania (Leishmania) mexicana have been documented in the state, showing a geographical expansion of the disease.
Materials and Methods:
Using CO2-supplemented Centers for Disease Control and Prevention light traps and Shannon traps, we captured anthropophilic sandflies in the surroundings of a locality with recent records of autochthonous cases of LCL. Sandflies carrying Leishmania DNA were evidenced using PCR.
Results:
A total of 140 Phlebotominae (Diptera: Psychodidae) females of four species were captured: Lutzomyia (Tricholateralis) cruciata (Coquillett), Psathyromyia (Psathyromyia) shannoni (Dyar), Lutzomyia (Lutzomyia) longipalpis (Lutz and Neiva), and Dampfomyia (Coromyia) deleoni (Fairchild and Hertig). Molecular results showed that 6.1% (95% confidence interval [CI] = 2.3–12.9%) of Lu. cruciata and 43.8% (95% CI = 19.8–70.1%) of Pa. shannoni showed evidence of carrying L. (L.) mexicana DNA.
Conclusion:
We provide evidence of anthropophilic sandflies carrying L. mexicana DNA in a municipality with recorded autochthonous cases of LCL caused by this parasite species in the state of Yucatan, suggesting the emergence of new focus of LCL in Mexico.
Introduction
The leishmaniases are a group of diseases caused by intracellular protozoan parasites of the genus Leishmania transmitted to humans and other mammals through the bite of hematophagous dipterans of the family Psychodidae, subfamily Phlebotominae, in the American continent (Galati, 2003).
The natural transmission cycle of Leishmania begins when a susceptible sandfly feeds on an infected mammalian reservoir and acquires the parasite, which will multiply in its digestive system. The vector carrying the parasite will transmit it to a new susceptible host during the blood feeding. In this context, humans are incidental hosts of different leishmaniases (PAHO, 2019).
The clinical manifestations of the disease include cutaneous leishmaniasis (CL), mucocutaneous leishmaniasis, visceral leishmaniasis, and finally, many people have the infection with an asymptomatic condition (PAHO, 2022b; PAHO, 2019).
Seventy-five percent of cases of leishmaniasis in the Americas belong to the CL form, although it occurs more frequently in 18 endemic countries, including Mexico (PAHO, 2022a), where 99% of the cases correspond to the localized cutaneous clinical form (LCL), caused in 97% of cases by Leishmania (Leishmania) mexicana Biagi and by Leishmania (Viannia) braziliensis Vianna in the remaining 3% (CENAPRECE, 2015; Hernández-Rivera et al, 2015; InDRE, 2019).
LCL cases are reported predominantly in the south-southeast region of Mexico, including Tabasco state, Chiapas state, and the Yucatan Peninsula which includes Campeche, Quintana Roo, and Yucatan states. In 2019, 62% of the cases of localized cutaneous leishmaniasis (LCL) in Mexico occurred in Campeche and Quintana Roo states, while Yucatan state remains a vulnerable and receptive area due to its neighboring endemic states and vector sandfly species inhabiting the area (according to PAHO, 2019).
Historical record in the state shows that few LCL cases have been reported in the southern part of the state, just next to the geographic limits of Campeche and Quintana Roo. During the period of 1984 to 2014, only 26 cases of LCL were recorded by Mexican health authorities while an immunoreactive epidemiological report from Becanchén locality published in 1996 showed a prevalence of 17% of people with evidence of asymptomatic infection (Albertos-Alpuche et al, 1996; Secretaría de Salud México, 2021a).
However, since 2015 the epidemiological records in Yucatan state have shown an increase in the number of cases, and from 2015 to 2019 there were 119 LCL cases, of which 69.8% were imported cases and 30.2% autochthonous cases from 13 different localities, evidencing the geographic expansion of the disease and suggesting the emergence of a new transmission focus of LCL in Yucatan state (Canché-Pool et al, 2022; Secretaría de Salud México, 2021a).
Historically, the transmission cycle of Leishmania in Campeche and Quintana Roo states involves several rodent hosts and Bichromomyia olmeca olmeca (Vargas and Díaz-Nájera) as the primary vector (Biagi et al, 1965) and Lutzomyia (Tricholateralis) cruciata (Coquillett) and Psathyromyia (Psathyromyia) shannoni (Dyar) as possible important primary or secondary vectors due to their anthropophilic nature, abundance in the endemic areas, and molecular evidence of carrying Leishmania (Pech-May et al, 2010; Sánchez-García et al, 2010). Nonetheless, recent evidence suggests that these species have a major role in the parasite's transmission (Pech-May et al, 2016).
In the state of Yucatan, 15 species of sandflies have been documented: Brumptomyia hamata (Fairchild and Hertig), Brumptomyia mesai Sherlock, Dampfomyia (Coromyia) beltrani (Vargas and Díaz-Nájera), Dampfomyia (Coromyia) deleoni (Fairchild and Hertig), Pintomyia (Pifanomyia) serrana (Damasceno and Arouck), Psathyromyia (Forattiniella) mexicana (Fairchild and Hertig), Psathyromyia cratifer (Fairchild and Hertig), Psathyromyia undulata (Fairchild and Hertig), Micropygomyia (Coquillettimyia) chiapanensis (Dampf), Micropygomyia cayennensis maciasi (Fairchild and Hertig), Micropygomyia (Sauromyia) trinidadensis (Newstead), and the remarkable important vector species Lutzomyia (Lutzomyia) longipalpis (Lutz and Neiva), Lu. (Trl.) mexicana, Bi. olmeca olmeca, and Pa. (Pa.) shannoni; however, no sandfly specimens have yet been recorded carrying Leishmania for localities where autochthonous LCL cases occur (Canché-Pool et al, 2022; de Oca-Aguilar et al, 2013; Ibáñez-Bernal and Durán-Luz, 2022; Loría-Cervera et al, 2019; Navarrete-Carballo et al, 2022; Rebollar-Téllez and Manrique-Saide, 2001; Rebollar-Téllez et al, 2006).
The objective of this study is to document for the first time the molecular evidence of anthropophilic Phlebotominae naturally carrying the parasite L. (L.) mexicana from a locality with recent records of autochthonous LCL cases in the state of Yucatan as complementary evidence of focus emergence.
Materials and Methods
Study area
Based on the recent epidemiological context in the state of Yucatan, one of the localities with a recent report of autochthonous LCL (Canché-Pool et al, 2022) was selected for this survey: the sylvatic area of Pustunich, municipality of Ticul, Yucatan, Mexico (20°22′4.08″ N, 89°30′46.08″ W; altitude of 25 meters a.s.l.). The area has secondary vegetation from a medium deciduous and semideciduous tropical forest with the following characteristic species: Acacia cornigera (L.) Willd., Acacia pennatula (Schltdl. and Cham.), Annona reticulata (L.), Bursera simaruba (L.) Sarg., Bucida buceras L., Cedrela odorata L., Cochlospermum vitifolium (Willd.) Spreng., Gliricidia maculata (Humb., Bonpl., and Kunth) Steud., Enterolobium cyclocarpum (Jacq.) Griseb., and Caesalpinia gaumeri Greenm. (Flores-Guido et al, 2010). The climate is tropical subhumid with summer rains; precipitation fluctuates between 1000 and 1300 mm per year, and the average annual temperature is 25°C (Orellana et al, 2010).
Entomological survey
In February 2020, the month of highest occurrence for Phlebotominae in the Yucatan Peninsula (Andrade-Narvaez et al, 2003), we set a transect of four Centers for Disease Control and Prevention (CDC)-type traps with a supplementary source of CO2 (dry ice for enhancing the capture of anthropophilic species) separated by 100 meters. Traps were active for one night for 4 h from the beginning of twilight under favorable meteorological conditions for collection (i.e., no rain or winter storm wind).
In addition, two Shannon traps separated by 1 km were active for at least 3 h from twilight. Two expert trained personnel protected with adequate clothing, without exposed skin areas, went into each trap to capture sandflies using manual aspirators immediately upon their landing on the walls of the Shannon trap (NORMA Oficial Mexicana NOM-032-SSA2-2014, 2015; Rodríguez-Rojas and Rebollar-Téllez, 2017). Four automated Thermo hygrometers (Process Parameter Ltd, El-USB-2®) were used to collect information regarding the temperature and relative humidity during the fieldwork. Ethics approval was not required for this study; the survey was part of the routine practices promoted by SSY and the Mexican Ministry of Health programs (Secretaría de Salud México, 2015). Through these programs, trained personnel continuously conduct active entomological surveillance in the state of Yucatan based on national and international guidance (InDRE, 2022; NORMA Oficial Mexicana NOM-032-SSA2-2014, 2015; PAHO, 2019; Secretaría de Salud México, 2021b).
Specimens were kept in dry collecting containers and stored in regular field coolers for transportation to a laboratory facility. Once in the laboratory, containers were stored at 4°C for a maximum of 24 h and then dead specimens were transferred to a new clean container with 75% ethanol where sandflies were separated morphologically from other insects, under a Nikon® SMZ645 stereomicroscope using cold light and a chill table to uphold the temperature of the specimens, and transferred individually to 1.7 mL sterile Eppendorf® tubes with 75% ethanol and stored again at 4°C until their taxonomic determination.
Taxonomic identification
Sandflies were morphologically classified by sex and morphospecies under the stereoscope using disposable slides, cold light, and a chill table to uphold the temperature of the specimens. The morphological identification of species was done through three to four female and male specimens of each morphospecies prepared for permanent slides in Canada balsam following the protocol of Ibáñez-Bernal (1999). Species determination was done with a microscope Motic® BA300 for observation of the antennal configuration, male terminalia, cibarium structure, and spermathecae through taxonomic keys (Ibáñez-Bernal, 2005; Ibáñez-Bernal, 2002; Young and Duncan, 1994) following the classification of Galati (2003); the genus abbreviations used were those proposed by Marcondes (2007, 2019). The males were observed under the microscope in a temporary slide preparation and then stored again.
The remaining female specimens were dissected, separating the head and the last three segments of the abdomen for specific determination, and the thorax and remaining abdomen were processed for molecular procedures. All sandflies were kept and handled individually to avoid contamination.
DNA extraction
A GenElute® Mammalian Genomic DNA Miniprep Kit (St. Louis, MO) protocol was used for DNA extraction. Individual sandflies were frozen at −70°C for 30 min and then macerated using a disposable pestle in a 1.5 mL tube, both sterile. To avoid cross-contamination between samples, one pestle and 1.5 mL tube per sample were used. Then, 180 μL of lysis solution and 20 μL of proteinase K solution were added and vortexed. The tube was incubated at 55°C for 3 h and vortexed occasionally after digestion was complete.
Then, 200 μL of lysis solution was added, thoroughly vortexed (15 s), and incubated at 70°C for 10 min.
A binding column was assembled with a 2 mL collection tube. Then, 500 μL of column preparation solution was added to the binding column and centrifuged at 12,000 g for 1 min and then flow-through liquid was discarded.
Ethanol (200 μL; 95–100%) was added to the lysate mixture and mixed by vortex thoroughly (5–10 s). Then, it was transferred into the treated binding column and centrifuged at ≥6500 g for 1 min. The tube containing the flow-through liquid was discarded, and the binding column was placed in a new 2 mL tube. Five hundred microliters of the wash solution was added to the binding column and centrifuged for 1 min at ≥6500 g and then the flow-through liquid was discarded. Another 500 μL of the wash solution was added to the binding column and centrifuged for 3 min at maximum speed (12,000–16,000 g) to dry the binding column. The binding column was centrifuged for one additional minute to remove residual ethanol. Finally, the collection tube containing the flow-through liquid was discarded, and the binding column was placed in a new 2 mL collection tube.
For DNA elution, 40 μL of elution solution was added directly to the binding column, incubated for 5 min, and centrifuged for 1 min at ≥6500 g. DNA was stored at −20°C and then quantified with a NanoDrop® 2000 spectrophotometer (Thermo Scientific) and used for PCR amplification. The average DNA concentration of sandfly samples was 14.06 ng/μL (ranging from 4.2 to 26.6 ng/μL) in a total volume of 40 μL.
For the positive control, DNA of L. mexicana (MNYC/BZ/62/M379) was used. DNA was extracted from 1 × 106 parasites using a QIAGEN DNeasy Blood and Tissue Kit (Germany) according to the manufacturer's guidelines, quantified, and used for PCR amplification.
Molecular identification of L. (L.) mexicana in sandflies was done by conventional end-point PCR. We used the primers IR1 and LM17 to amplify a 790 bp specific genetic sequence of this parasite species. The IR1 forward sequence (5′-GCT GTA GGT GAA CCT GCA GCA GCT GGA TCA TT-3′) corresponds to the 32 final nucleotides of the conserved sequence from the 3′ region of the small subunit of 18S ribosomal gene (Cupolillo et al, 1995). The LM17 reverse sequence (5′-CCC CTC TCC TCC TCC CC-3′) corresponds to a specific region of the internal transcribed spacer of the ribosomal RNA gene (Berzunza-Cruz et al, 2009).
The PCR was completed with 50 μL of the following reaction mix: 25 μL of 2 × Thermo Scientific® DreamTaq® Green PCR Master Mix (Dream Taq DNA polymerase, 2 × Dream Taq Green Buffer, dATP, dCTP, dGTP, and dTTP, 0.4 mM each, and 4 mM MgCl2) (Thermo Scientific), 100 ng of primers IR1 and LM17, 10 μL of DNA (50–100 ng), and nuclease-free water to 50 μL. The amplification was performed in a 2720 Thermal Cycler (Applied Biosystems®, Singapore). Amplification conditions included one cycle at 94°C for 5 min, 30 cycles of 1 min at 94°C, 1 min at 65°C, and 1 min at 72°C, and one extension cycle at 72°C for 7 min (Berzunza-Cruz et al, 2009). For the negative control, nuclease-free sterile water instead of DNA was used in the reaction mixture. For the positive control, DNA of L. mexicana (MNYC/BZ/62/M379) was used.
The PCR products were analyzed by electrophoresis in 1.2% agarose gels in 1 × Tris-acetate-EDTA (TAE) buffer at 1000 V. The gels were stained with 1 μL of SYBR® safe (Invitrogen, Thermo Scientific) for every 10 mL of 1 × TAE buffer and visualized in a high-performance UV transilluminator Gel DocTM XR+ System® (Bio-Rad Laboratories, Inc.).
Data analysis
The prevalence of Leishmania carriers and their 95% confidence interval (CI) were estimated using the Clopper–Pearson method with Quantitative Parasitology Web software (Reiczigel et al, 2019).
Results
A total of 140 female and 238 male specimens belonging to species Da. deleoni (1 female), Lu. longipalpis (18 female, 224 male), Pa. shannoni (23 female, 13 male), and Lu. cruciata (98 female, 1 male) were captured. The captures occurred between 18:00 and 21:00 h completing a total of 24 trap-hour collecting effort; the average temperature and relative humidity during sandfly captures were 25.9°C and 70.3%, respectively.
Of all specimens collected, 37 female and 23 male were from the CDC+CO2 traps, and the remaining 103 female and 215 male were from the Shannon trap. Collection of Da. deleoni (1 female), Pa. shannoni (16 female, 8 male), Lu. longipalpis (206 male), and Lu. cruciata (86 female, 1 male) was in the Shannon traps. Captures in the CDC+CO2 traps were Lu. longipalpis (18 female, 18 male), Lu. cruciata (12 female), and Pa. shannoni (7 female, 5 male).
The results of the molecular analysis for Leishmania showed that six female specimens belonging to Lu. cruciata and seven specimens of Pa. shannoni showed evidence of carrying L. (L.) mexicana (Figs. 1 and 2), which implies an infection prevalence of 6.1% (95% CI = 2.3–12.9%) and 43.8% (95% CI = 19.8–70.1%), respectively. Prevalence estimation for Pa. shannoni was based on 16 specimens, because four were used for taxonomic determination and three were in suboptimal conditions for molecular procedures.
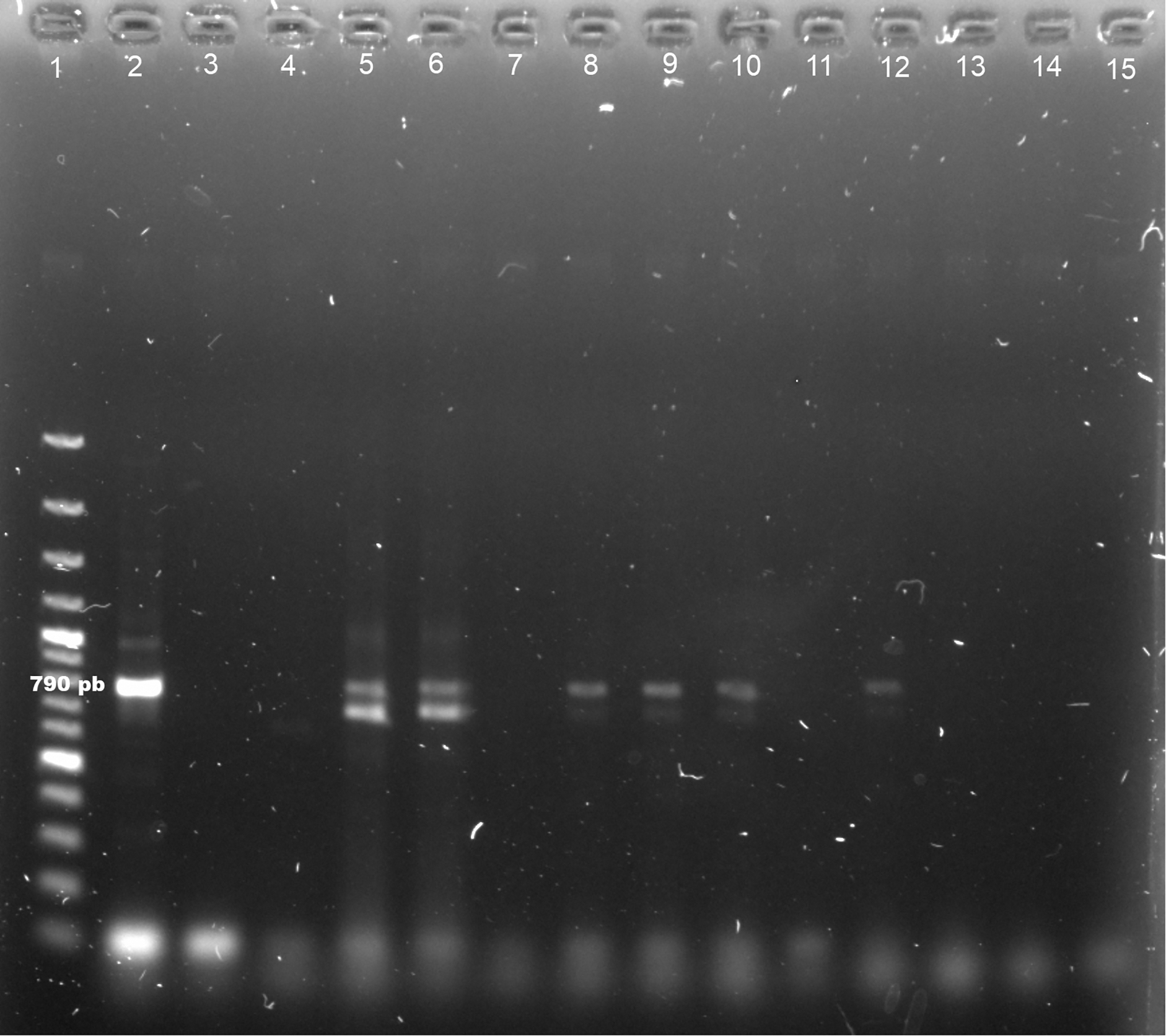
Detection of Leishmania (Leishmania) mexicana DNA in female specimens of Lutzomyia cruciata by PCR analysis. L. mexicana (IR1/LM17 primers, 790 pb). Line 1: 100 bp DNA ladder, Line 2: positive control L. mexicana parasite DNA (MNYC/BZ/62/M379), Line 3: negative control (no DNA), Lines 5, 6, 8–10, 12: Lu. cruciata positive for L. mexicana, Lines 4, 7, 11, 13–15: Lu. cruciata negative for L. mexicana.
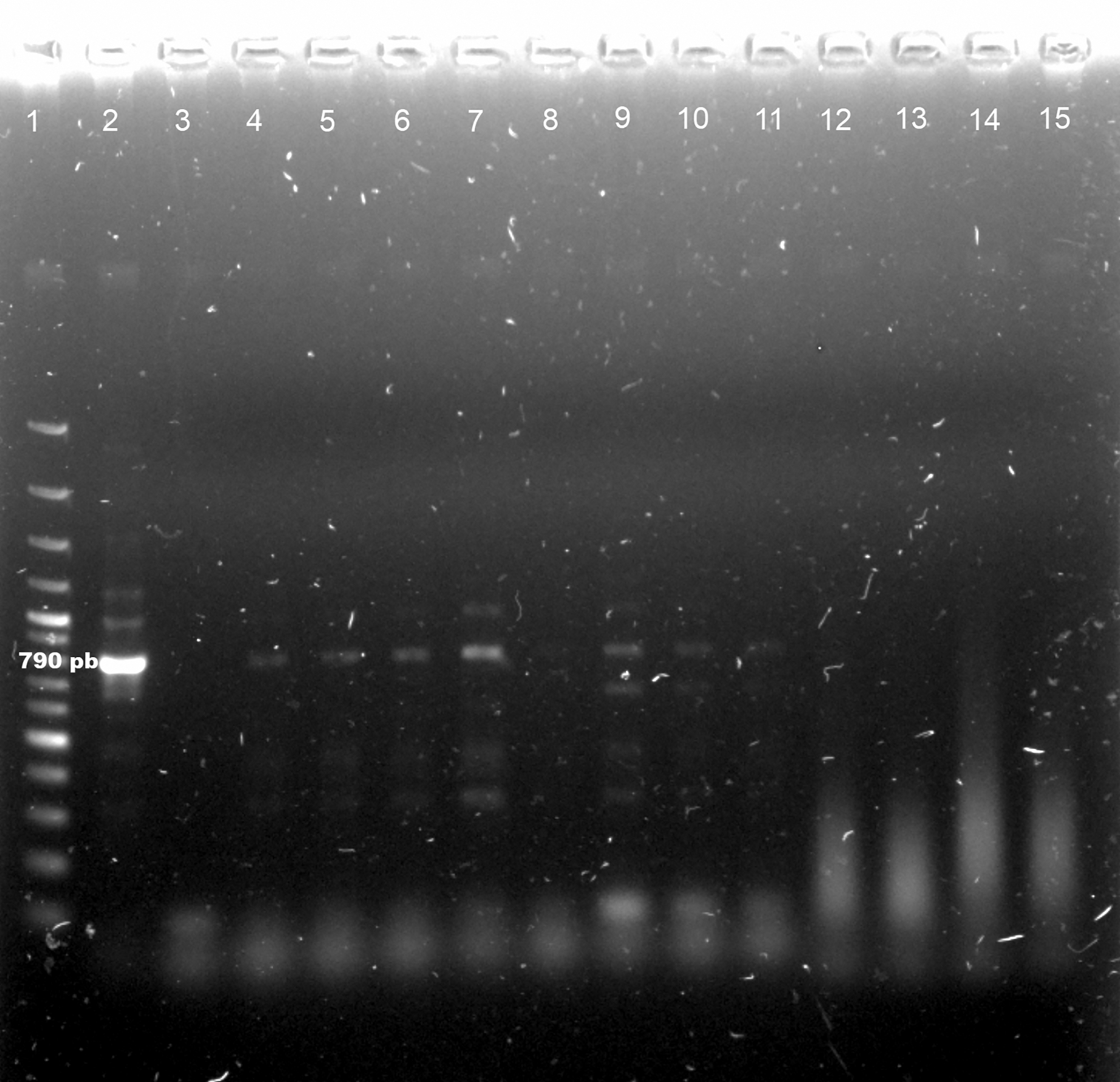
Detection of Leishmania (Leishmania) mexicana DNA in female specimens of Psathyromyia shannoni by PCR analysis. L. mexicana (IR1/LM17 primers, 790 pb). Line 1: 100 bp DNA ladder, Line 2: positive control L. mexicana parasite DNA (MNYC/BZ/62/M379), Line 3: negative control (no DNA), Lines 4–7, 9–11: Pa. shannoni positive for L. mexicana, Lines 8, 12–15: Pa. shannoni negative for L. mexicana.
Discussion
This study reports for the first time the presence of anthropophilic sandflies carrying L. mexicana in a locality with recent previous occurrence of autochthonous cases of LCL caused by the same parasite species in the state of Yucatan (Canché-Pool et al, 2022), a region that, according to PAHO, is vulnerable and receptive to the transmission of leishmaniasis due to its proximity to the endemic states of Campeche and Quintana Roo and because of the presence of important vector sandflies like Lu. longipalpis, Lu. cruciata, Bi. olmeca olmeca, and Pa. shannoni in several localities of the state (Ibáñez-Bernal and Durán-Luz, 2022; PAHO, 2019; Rebollar-Téllez and Manrique-Saide, 2001; Rebollar-Téllez et al, 2006).
The results showed the presence of four sandfly species at the study site. Although not all the species were captured using the Shannon trap (Lu. longipalpis was only collected with CDC+CO2 traps), there is evidence that females of all identified sandflies may exhibit anthropophilic behavior (Navarrete-Carballo et al, 2022; Young and Duncan, 1994).
Of the collected sandflies, Lu. cruciata and Pa. shannoni are the species of most significant interest in the transmission of L. mexicana as probable primary and secondary vectors in different regions of Mexico and Central America (Pech-May et al, 2016; Pech-May et al, 2010).
The species recognized as the primary vector of L. mexicana in Mexico is Bi. olmeca olmeca (Biagi et al, 1965). However, its geographic distribution is not uniform in the country, and some studies suggested that it may be absent or have shallow populations in regions with transmission foci (González et al, 2011; May-Uc et al, 2011; Navarrete-Carballo et al, 2022; Pech-May et al, 2010). In this study we did not collect Bi. olmeca olmeca; nonetheless, it is a species that is distributed in the region (Ibáñez-Bernal and Durán-Luz, 2022). Longitudinal studies must help to describe and understand the whole community of sandflies in the area and their population ecology, including the driving factors of their presence and local abundance.
In this context, leishmaniasis transmission cycles have a focal distribution in specific geographic areas (Hashiguchi et al, 2020). These sites are called natural foci of infection, where three critical elements converge within a favorable environment: competent and infectious vectors, competent vertebrate reservoirs, and susceptible hosts such as humans or domestic animals (Vélez et al, 2017).
These components, especially the vectors, are conditioned by abiotic environmental factors such as climate, humidity, altitude, temperature, and vegetation type. The boundaries of transmission foci correspond to the spatial distribution and relative density of vector species (Reisen, 2010). Therefore, knowledge of the behavior of vectors is essential to understanding the epidemiological risk of infection (Vélez et al, 2017).
Using ecological network models, González-Salazar and Stephens (2012) proposed a predictive map of leishmaniasis transmission risk based on geographic records of host distribution and their interaction with sandfly vectors and vegetation cover. Although the model was entirely spatial, the predictive scenario for the Yucatan state clearly showed, without a temporal context, the potential high risk of Leishmania transmission for this area of the Yucatan Peninsula.
More recent studies based on ecological niche models showed meaningful forecasts for expansion of the distribution of vectors such as Lu. cruciata and some host rodents such as Yucatan deer mouse (Peromyscus yucatanicus Allen and Chapman) and Gaumer's spiny pocket mouse (Heteromys gaumeri Allen and Chapman) (Moo-Llanes et al, 2019; Moo-Llanes et al, 2013), leading to potential risk of an emergent transmission focus in Yucatan state because these species, as well as Ototylomys phyllotis Merriam, tolerate changes of land use associated with human activities, including the use of rural and suburban peridomiciles and the empirical evidence of carrying L. mexicana (Reyes-Novelo et al, 2011; Sosa-Bibiano et al, 2022).
The territory of Yucatan state has been experiencing a noticeable increase of intensification of land use and human activities like stone mining (Estrada-Medina et al, 2019), urban expansion, agricultural land expansion (Ramírez-Carrillo, 2020), and pollution (Reyes-Novelo et al, 2022), bringing expected environmental consequences like forest fragmentation, species assemblage changes, and modification of ecosystem functioning. In this context, several studies have shown that land-use changes lead to vector-borne disease outbreaks as eco-epidemiological consequences like those described for leishmaniases and other parasitic diseases in the Americas (de Oca-Aguilar et al, 2022; Guo et al, 2019; Travi et al, 2002).
Recent studies in Yucatan state describe new insights of eco-epidemiology of L. mexicana hosts in companion animals (Arjona-Jiménez et al, 2012; Longoni et al, 2011; López-Céspedes et al, 2012), wild rodents (Sosa-Bibiano et al, 2022), and people (Canché-Pool et al, 2022). In addition, potential vectors occur in different localities and ecotopes (de Oca-Aguilar et al, 2022; Navarrete-Carballo et al, 2022). Although there are several forecasts for the emergence of new foci of transmission of L. mexicana, there is still little evidence at the local scale about the effect of the increasing environmental changes in sites vulnerable and susceptible to endemic transmission, particularly those of anthropic origin (Reyes-Novelo et al, 2022).
The increase in the number of cases of local origin in recent years in Yucatan state suggests the occurrence of changes in the dynamics of the eco-epidemiological components involved in the transmission cycles of leishmaniasis, which could be favoring the emergence and persistence of zoonotic foci of transmission in this state (Canché-Pool et al, 2022).
Therefore, given the previous description of emergence and progressive increase of autochthonous LCL cases, considering the finding of anthropophilic species such as Lu. cruciata and Pa. shannoni carrying L. mexicana, and the existence of favorable eco-epidemiological factors like land-use changes, forest fragmentation, and rural and suburban geographic growth in the region for its persistence and expansion, the support and continuity of an active epidemiological surveillance program of leishmaniasis transmission in the state of Yucatan are essential to prevent major outbreaks.
Footnotes
Acknowledgments
The authors thank Yucatan Health Services personnel (SSY) from the vector control program, particularly Hugo Itzá for his active help and assistance in the fieldwork.
Authors' Contributions
Conception and design of the study: L.G.P.-B., E.B.C.-P., E.R.-N. Acquisition of data: L.G.P.-B., A.G.C.-A., E.B.C.-P., R.T.-M., J.R.T.-C., E.R.-N. Analysis and interpretation of data: E.R.-N., E.B.C.-P. Drafting the article: E.R.-N. Revising article critically for important intellectual content: H.A.R.-P., R.F.-M., O.R.C., E.R.-N., E.B.C.-P. All authors read and approved the final version of the article.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This research is supported by grant CIRB-2021-0004. E.R.-N. and E.B.C.-P. were supported by Consejo Nacional de Ciencia y Tecnología (47934 and 53525, respectively).