
Abstract
Background:
Leptospirosis represents a major public health threat in Cambodia. To help establish preventive strategies for leptospirosis, we isolated and identified Leptospira from soil collected in Phnom Penh and its neighboring province during the dry and rainy seasons.
Materials and Methods:
Soil samples were collected from 15 sites in the Phnom Penh metropolitan area and Kandal Province in 2019 and 2020. A total of 120 soil samples were collected and analyzed after culturing. Cultured isolates were identified at the species level using 16S rRNA and gyrB gene sequencing analysis.
Results:
The positive culture rate for Leptospira was significantly higher for samples collected during the rainy season (31 out of 60 samples, 51.7%) than during the dry season (19 out of 60 samples; p < 0.05, chi-squared test). In terms of sampling area, 10 (35.7%) and 13 (46.9%) soil samples from Phnom Penh and 9 (28.1%) and 18 (56.3%) soil samples from Kandal Province were culture-positive when collected during the dry and rainy seasons, respectively. Three isolates from the Kandal samples collected during the dry season were categorized as pathogenic clade type P1 according to 16S rRNA and gyrB sequence analyses. In addition, samples collected during the rainy season from Phnom Penh and Kandal Province (three each) were categorized as clade type P1. The samples with isolates in clade type P1 were collected from riverbanks during both the dry and rainy seasons and from a hospital garden during the rainy season.
Conclusions:
This is the first report on the isolation of clade P1 Leptospira species in environmental soil samples from Cambodia. Environmental soil appears to be a reservoir for Leptospira in the capital city and surrounding areas of Cambodia. This study contributes to the understanding of Leptospira species distribution in Asian countries.
Introduction
Leptospirosis is a worldwide re-emerging and neglected zoonotic disease caused by infection with aerobic gram-negative spirochetes of the genus Leptospira (Adler and de la Pena Moctezuma, 2010; Hartskeerl et al., 2011). It commonly occurs in tropical and subtropical countries, affecting more than a million people and causing 60,000 deaths annually (Costa et al., 2015).
The classification of Leptospira was updated from three (pathogenic, intermediate, and saprophytic) to two clades (pathogenic and saprophytic) after comparative genomic analyses in 2019 (Vincent et al., 2019). The pathogenic clade is further subdivided into pathogenic (P1) and intermediately pathogenic (P2). Similarly, the saprophytic clade is further divided into a saprophytic subclade (S1) and a new subclade (S2) that was separated from S1 by molecular phylogeny.
Reservoir animals for Leptospira include wild rodents, cattle, and canines, in which the pathogen is maintained in the kidneys. Humans and other animals acquire leptospirosis through exposure to soil or water contaminated with reservoir animal urine that contains pathogenic Leptospira (Adler and de la Pena Moctezuma, 2010). Thus, occupational contact with natural water, especially by farmers, outdoor aquatic sports instructors, and military personnel, is a potential transmission route for Leptospira (Atil et al., 2020). Moreover, some outbreaks of leptospirosis are associated with heavy floods caused by typhoons (Saito et al., 2014; Su et al., 2011).
Given the epidemiological significance of leptospirosis, identification of Leptospira in environmental samples is important. Leptospira can be isolated from environmental samples, such as soil, using a novel selective medium known as STAFF (Chakraborty et al., 2011), which comprises a cocktail of antimicrobial agents, including sulfamethoxazole, trimethoprim, amphotericin B, fosfomycin, and 5-fluorouracil. There are several reports on the appearance of Leptospira in water and soil samples. For example, Slack et al. (2009) identified L. kmetyi in environmental samples in Malaysia, and 75% of soil and water samples in an elephant camp in Thailand were Leptospira culture-positive (Chaiwattanarungruengpaisan et al., 2020).
Using STAFF, 40% of soil and water samples from Metro Manila and the surrounding area of the Philippines and 46% of samples from Fukuoka, Japan, were culture-positive for Leptospira (Saito et al., 2013). Furthermore, Masuzawa et al. (2018) reported that 37.9% of soil samples from Japan were Leptospira culture-positive compared with 57.7% of soil samples from New Caledonia (Thibeaux et al., 2017).
In Cambodia, 8.3% and 3% of humans with typical symptoms of leptospirosis in Takeo Province and the Phnom Penh metropolitan area, respectively, were positive for Leptospira antibodies in serological tests (Hem et al., 2016; Laras et al., 2002). In addition, Kudo et al. (2018) reported the presence of Leptospira in 12.3% of wild rodents tested in Cambodia.
To our knowledge, the isolation of Leptospira from soil samples in an urban area of Cambodia has not been reported. Therefore, we aimed to isolate and identify Leptospira from soil samples collected in Phnom Penh and its neighboring province during the dry and rainy seasons. Further characterization of Leptospira present in soil, regardless of the season, will better inform the formulation of preventive and control strategies against leptospirosis.
Materials and Methods
Study site
Phnom Penh is the capital city of and most populous area in Cambodia (National Institute of Statistics, 2019). It neighbors Kandal Province, which contains commuter towns. The Bassac River flows through Phnom Penh to Kandal Province (Fig. 1) and is utilized for fishing, business, and leisure.
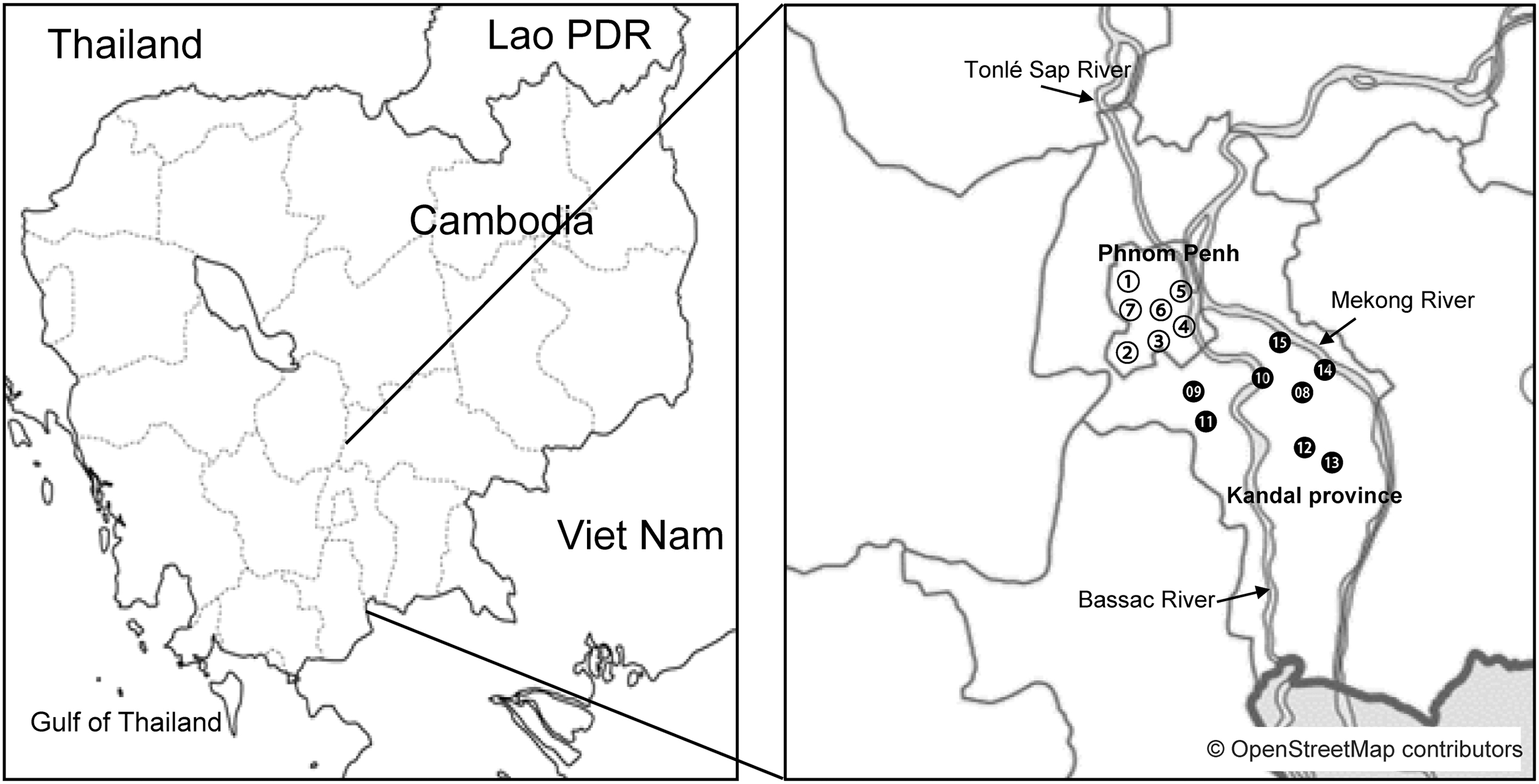
Sampling sites in Phnom Penh (○) and Kandal Province (●). The land at the numbered sites was used as follows: hospital (1, 3, 7, 9, 10, 12), residential area (2), temple (11), park (4), riverbank (5, 13, 15), market (6, 8), and restaurant (14).
Collection of soil samples
Soil samples (∼10 g) were collected from the soil surface using a spatula in June 2019 (rainy season) and February 2020 (dry season) from seven sites in Phnom Penh and eight points in Kandal Province (Fig. 1). These sites included hospital gardens, markets, restaurants, and riverbanks. At each site, 4 soil samples were taken, totaling 60 samples per season or 120 samples over 2 seasons. To avoid drying, samples were kept in sealed plastic bags at room temperature in the dark. Culturing of soil samples was initiated within 12 h of sampling in Phnom Penh.
Culture and isolation method
An aliquot soil sample (5 g) was placed in a test tube containing 9 mL of 20 mM HEPES buffer (pH 7.4), mixed well, and allowed to settle for 1 h. Subsequently, 2 mL of supernatant was transferred to a new test tube containing 2.5 mL of 2 × Korthof's medium and 0.5 mL of 10 × STAFF (400 μg/mL sulfamethoxazole, 200 μg/mL trimethoprim, 50 μg/mL amphotericin B, 4 mg/mL fosfomycin, and 1 mg/mL 5-fluorouracil), and incubated at 30°C for 1 month (Chakraborty et al., 2011). The cultures were monitored daily by dark field microscopy.
Approximately 1 month later, when Leptospira was detected and the sample was found to be contaminated with other microorganisms after microscopic observation, the sample was filtered through a 0.2-μm pore-size membrane filter to remove contaminants. An aliquot of the filtered sample (0.5 mL) was then transferred to a new test tube containing Korthof's medium without antibiotics. For Leptospira isolation, 100 μL of the culture was inoculated onto Korthof's agar for colony isolation. When a single colony was observed by visual inspection, a pinprick-sized colony was transferred into Korthof's medium without antibiotics and incubated until Leptospira growth became confluent.
DNA extraction and analyses
Cultures with confluent growth were separated by centrifugation (10,000 × g, 10 min) at 4°C. DNA was extracted from the pelleted Leptospira cells using a QIAamp DNA Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer's instructions.
The extracted DNA was subjected to 16S rRNA gene target sequencing to identify species of Leptospira isolates at the clade level. The forward primer was 8UA (5′-AGAGTTTGATCMTGGCTCAG-3′) and the reverse primer was 1485R (5′-TACGGYTACCTTGTTACGACTT-3′). The target gene was ∼1430 base pairs long (Masuzawa et al., 2018).
Furthermore, gyrB gene target sequencing was performed using the same DNA samples as those used in the 16S rRNA gene analysis. The forward primer was UP1TL (5′-CAYGCNGGNAARTTYGA-3′) and the reverse primer was UP2rTL (5′-TCNACRTCNGCRTCNGTCAT-3′). The target gene was ∼1400 base pairs long (Villanueva et al., 2014).
The purified PCR 16S rRNA and gyrB amplicons were subjected to DNA sequence analysis using the Eurofins Genomics sequencing service (Tokyo, Japan). The 16S rRNA and gyrB sequences were assembled to constitute a contig sequence using Genetyx Ver. Ten (Hitachi, Tokyo, Japan). Phylogenetic analysis was performed using the Clustal W algorithm created by MegAlign (DNASTAR, Madison, WI). A phylogenetic tree was constructed using the neighbor-joining method with 1000 bootstrap resampling. The obtained DNA sequence data were deposited and assigned to the DNA Data Bank of Japan (DDBJ).
This study was reviewed and approved by the Ethics Committee of the Ministry of Health, Royal Government of Cambodia.
Results
Isolation of Leptospira spp. from soil samples
Of the 120 samples collected during the dry and rainy seasons (60 samples each season), 19 (31.7%) and 31 (51.7%) samples, respectively, were Leptospira culture-positive. The culture positivity rate was significantly higher (p < 0.05, chi-squared test) for the rainy season samples compared with the dry season samples. Regarding the source of the samples, 10 (35.7%) and 9 (28.1%) soil samples collected during the dry season from Phnom Penh and Kandal Province, respectively, were culture-positive, compared with 13 (46.4%) and 18 (56.3%) samples collected during the rainy season from Phnom Penh and Kandal Province, respectively (Table 1).
Number of Isolated Leptospira spp. from Soil Samples According to Gene Type, Clade Type, and Sampling Area
p < 0.05, chi-squared test, total samples in the rainy season compared with total samples in the dry season.
p < 0.05, chi-squared test, samples from Phnom Penh in the rainy season compared with samples from Phnom Penh in the dry season.
Identification of Leptospira species
The results of the phylogenetic tree analysis based on Basic Local Alignment Search Tool (BLAST) analysis of the 16S rRNA and gyrB sequences from isolated Leptospira are shown in Figs. 2 and 3. During the dry season, three samples from Kandal Province were categorized as clade type P1 (Gen Bank accession number for the 16S rRNA gene sequence: LC595137; sample numbers D57-1, D59-2, and D60-1), and three samples from Phnom Penh were categorized as clade type P2 (LC595136—D17-1, D20-1; LC595135—D32-1). Furthermore, 13 samples from both areas during the dry season were categorized as clade type S1 (LC595130—D5-1, D8-1, D9-1, D37-1, D41-1, D52-1; LC595131—D7-1, D24-1, D30-1, D43-1; LC595132—D6-1, D42-2; LC595133—D8-2), and 1 sample from Kandal Province was categorized as clade type S2 (LC595134—D51-1).

Phylogenetic tree based on the alignment of 16S rRNA sequences of isolated Leptospira obtained from soil samples. We used a Clustal W algorithm followed by the neighbor-joining method with 1000 bootstrap resampling. Numbers in parentheses are accession numbers of the DDBJ. DDBJ, DNA Data Bank of Japan.
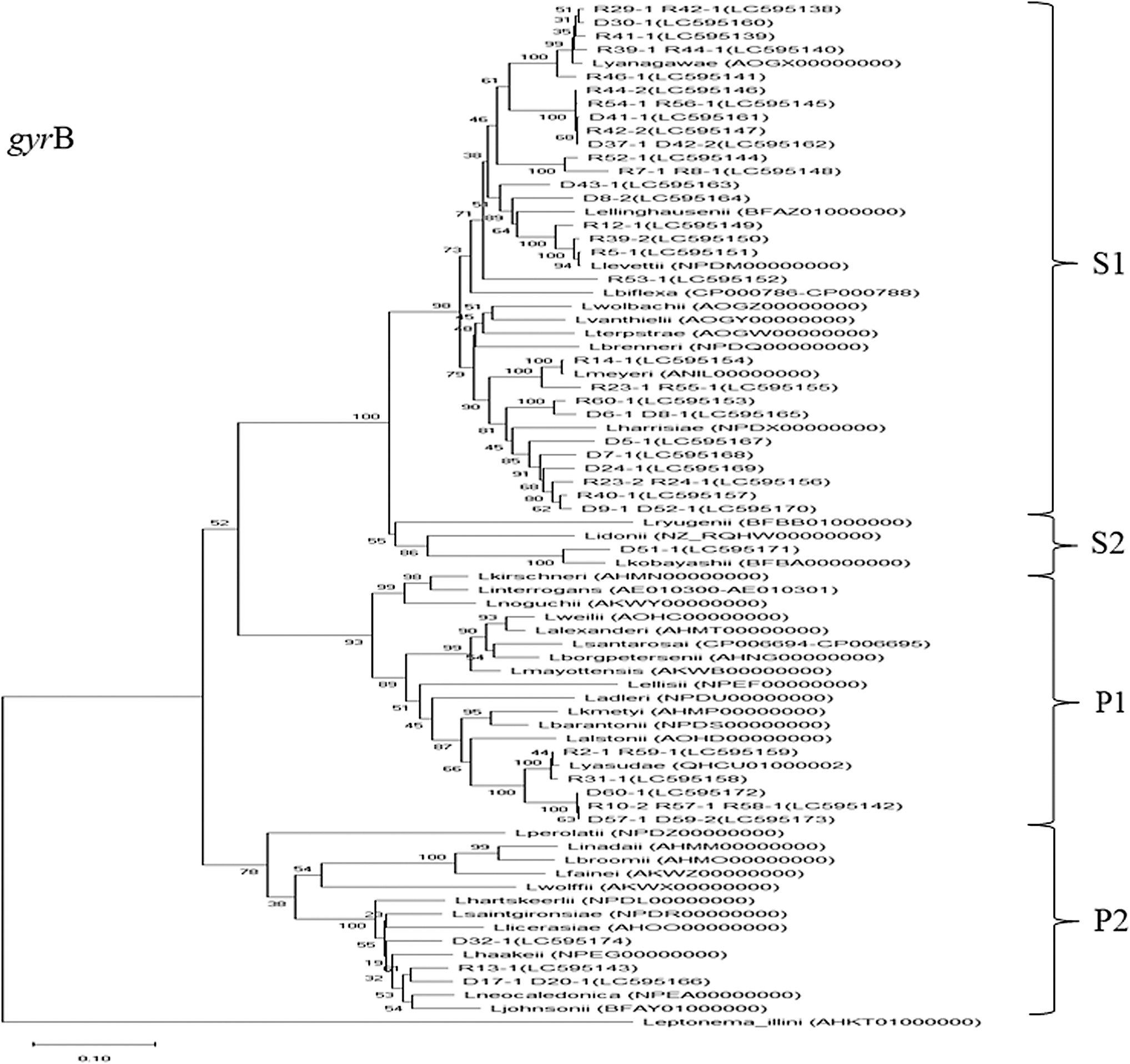
Phylogenetic tree based on the alignment of gyrB sequences of isolated Leptospira obtained from soil samples. We used a Clustal W algorithm followed by the neighbor-joining method with 1000 bootstrap resampling. Numbers in parentheses are accession numbers of the DDBJ.
During the rainy season, a total of six samples from Phnom Penh and Kandal Province were categorized as clade type P1 (LC595123—R10-2, R57-1; LC595128—R2-1, R31-1, R59-1; LC595129—R58-1), and one sample from Phnom Penh was categorized as clade type P2 (LC595124—R13-1). Furthermore, 24 samples from both areas during the rainy season were categorized as clade type S1 (LC595120—R8-1, R23-1, R39-1, R42-2, R44-1, R54-1, R60-1; LC595121—R56-1, LC595122—R12-1, R29-1, R42-1, R53-1; LC595125—R24-1, R39-2, R41-1, R52-1; LC595126—R5-1, R7-1, R23-2, R40-1, R44-2, R55-1; LC595127—R14-1, R46-1). However, there were no samples categorized as clade type S2 during the rainy season.
Samples categorized in clade type P1 were collected from riverbanks during the dry (D57-1, D59-2, and D60-1) and rainy seasons (R57-1, R58-1, and R59-1), and from the hospital gardens during the rainy season (R2-1, R10-2, and R31-1).
Discussion
This study revealed the presence of pathogenic Leptospira species clade type P1 in environmental soil samples obtained from land surfaces during the dry and rainy seasons in Phnom Penh and Kandal Province of Cambodia. All isolates belonging to clade type P1 were closely categorized as Leptospira yasudae (Figs. 2 and 3). L. yasudae was first described by Casanovas-Massana et al. (2020), who also reported that L. yasudae is the heterotypic synonym of Leptospira dzianensis (Casanovas-Massana et al., 2021). Casanovas-Massana et al. (2020) reported that L. yasudae did not induce acute disease or chronic carriage in a hamster model. Pathogenic strains are divided into two groups: virulent pathogens and low-virulence pathogens, according to their core and accessory genome phylogeny (Thibeaux et al., 2018). To determine the virulence of P1 isolates in this study, additional research is required, such as animal experiments and examination of core and accessory genome phylogeny.
During the rainy season, the culture positivity rate was 46.4% in the capital Phnom Penh and 56.3% in Kandal Province, for a combined positivity rate of 51.7%. However, during the dry season, the positivity rate was 35.7% in Phnom Penh and 28.1% in Kandal Province, for a combined rate of 31.7%. The higher positivity rate of Leptospira species in environmental soil samples during the rainy season than during the dry season was similar to that previously reported from other countries, such as New Caledonia and China (Cucchi et al., 2019; Thibeaux et al., 2017).
The average rainfall in Cambodia from 1991 to 2020 was relatively high from May to October and low from November to April (Asian Development Bank, 2021). In this study, the examined samples were obtained from the land surface without deeper drilling. Soil humidity should be higher during the rainy season as there is much higher rainfall than in the dry season. Several environmental factors, such as pH, temperature, and moisture, influence the survival of Leptospira, especially soil humidity (Andre-Fontaine et al., 2015). In this study, the increase in surface soil moisture may be related to the appearance of Leptospira spp. Saito et al. (2013) reported that a moisture content ≥20% and a pH value of 6.2–7.2 were suitable soil conditions for maintaining Leptospira, positively influencing their distribution and survival.
No statistically significant difference in the positivity rate of Leptospira was observed between the two study areas. However, the intended purpose of the land might influence the detection of pathogenic Leptospira. Clade type P1 Leptospira was isolated from samples obtained from the gardens of three different hospitals in Phnom Penh during the rainy season, and from riverbank samples in Kandal Province during the rainy and dry seasons. Similarly, Thibeaux et al. (2017) reported positive cultures of Leptospira from soil samples obtained during a leptospirosis outbreak in New Caledonia, but no positive cultures were obtained from the control area in which there were no cases of leptospirosis.
In Cambodia, L. interrogans and L. noguchii, which have significant virulence in humans, have been isolated from wild rodents (Kudo et al., 2018). L. interrogans is a slow-growing pathogenic species, which can be overgrown by fast-growing saprophytic species (Chaiwattanarungruengpaisan et al., 2018; Narkkul et al., 2020; Scialfa et al., 2018). To identify pathogenic Leptospira, such as L. interrogans and L. noguchii, further studies using a nonculture method, such as DNA metabarcoding, are needed (Sato et al., 2019). In this study, the majority of the isolates were categorized in the S1 clade; 77.4% were isolated during the rainy season and 63.2% during the dry season. One isolate in the S2 clade was detected during the dry season. Vincent et al. (2019) reported S2 isolates in Japan, Algeria, and mainland France; however, to our knowledge, this study is the first to detect a member of the S2 clade in the area of the Indochina peninsula.
The present investigation had some limitations. First, sequence analysis was performed using PCR targeting of 16S rRNA and gyrB genes; however, its discriminatory power was insufficient to achieve identification at the species level. Although the sequence analysis results did not show discordance between the 16S rRNA and gyrB genes, the number of sequence reads available for analysis was limited. To properly identify strains, further analysis of each isolate is required. Vincent et al. (2019) recommended the construction of a robust phylogenetic classification using the ppk gene of the core genome of Leptospira.
In addition, since we did not conduct animal experiments or employ other methods of virulence observation, the pathogenicity of clade type P1 isolates was not determined in this study. Moreover, sample numbers and target areas were limited. To fully understand the epidemiological mechanism of leptospirosis in humans and animals, the selection of target areas and sampling must consider the prevalence of leptospirosis according to environmental factors, such as typhoons and river flooding in Cambodia.
Conclusions
To the best of our knowledge, this is the first report on the isolation of clade P1 Leptospira species from environmental samples in both rainy and dry seasons in Cambodia. Our findings highlight environmental soil as a reservoir for Leptospira in the Cambodian capital city and surrounding areas. The positivity rate of Leptospira in soil samples was higher in the rainy season than in the dry season. These findings contribute to the understanding of Leptospira species distribution in Asian countries.
Footnotes
Acknowledgments
The authors would like to thank Ms. Hinako Izawa, Mr. Yudai Asano, and Ms. Shoko Fujimaki of the Department of Medical Technology and Sciences, International University of Health and Welfare, for their help with the laboratory work, as well as for their support to the team.
Authors' Contributions
Investigation (lead), software (lead), data curation (lead), and writing—original draft preparation (lead) by Y.A. Conceptualization (lead), investigation (supporting), writing—reviewing (lead), and supervision (lead) by Y.K. Conceptualization (equal) and supervision (equal) by V.K. Resources (lead) and investigation (supporting) by V.K. Resources (supporting) and investigation (supporting) by E.R. Methodology (lead), software (equal), and supervision (equal) by T.M.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This research was supported by education and research grants from the International University of Health and Welfare in 2018 and 2019, and the international research fund of the Heiwa Nakajima Foundation in 2018.