
Abstract
Background:
Chagas disease is one of the world's most neglected tropical diseases, infecting over six million people across the Americas. The hemoparasite Trypanosoma cruzi is the etiological agent for the disease, circulating in domestic, peridomestic, and sylvatic transmission cycles that are maintained by triatomine vectors and a diversity of wild and synanthropic hosts. Public health and wildlife management interventions targeting the interruption of T. cruzi transmission rely on an understanding of the dynamics driving the ecology of this zoonotic pathogen. One wildlife host that purportedly plays a role in the transmission of Chagas disease within the southern United States is the striped skunk (Mephitis mephitis), although infection prevalence in this species is poorly understood.
Materials and Methods:
To this end, we conducted a PCR-based surveillance of T. cruzi in 235 wild skunks, representing 4 species, across 76 counties and 10 ecoregions in Texas, United States, along with an evaluation of risk factors associated with the infection.
Results:
We recovered an overall T. cruzi prevalence of 17.9% for all mephitid taxa aggregated, ranging between 6.7% for plains spotted skunks (Spilogale putorius interrupta) and 42.9% for western spotted skunks (Spilogale gracilis). We report the first cases of T. cruzi infection in plains spotted and American hog-nosed skunks (Conepatus leuconotus), of important note for conservation medicine since populations of both species are declining within Texas. Although not statistically significant, we also detected trends for juveniles to exhibit greater infection risk than adults and for differential sex biases in T. cruzi prevalence between taxa, which align with variations in species-specific seasonal activity patterns. No geographic or taxonomic risk factors were identified.
Conclusion:
Our study contributed key data for population viability analyses and epidemiologic models in addition to providing a baseline for future T. cruzi surveillance among skunks and other wildlife species.
Introduction
The zoonotic hemoflagellate parasite Trypanosoma cruzi is the etiologic agent of Chagas disease (American trypanosomiasis), which is estimated to infect six to seven million people across the Americas (WHO, 2020). Although T. cruzi has historically inflicted the greatest social and economic costs on poor, rural populations throughout Latin America, a growing number of autochthonous cases of Chagas disease have been reported in the United States in recent years (Beatty and Klotz, 2020; Bern, 2015; Garcia et al, 2017; Garcia et al, 2015; Irish et al, 2022). In light of the underreporting of locally acquired infections and the globalized spread of infection via human migrations, the disease burden in the continental United States alone is estimated to be as high as 300,000 cases (Bern, 2015; Irish et al, 2022; Montgomery et al, 2016; Schmunis and Yadon, 2010). To mitigate the risk of Chagas disease in North America, effective vector control and disease surveillance strategies must be developed using a holistic understanding of the complex adaptive system that underlays T. cruzi's transmission cycle (Jansen et al, 2015; Levin, 2005; Moo-Millan et al, 2019; Silveira and Vinhaes, 1999). The domestic, peridomestic, and sylvatic transmission cycles of T. cruzi are maintained in the southern United States by 11 triatomine vectors and a diversity of wild, domestic, and exotic mammalian hosts, many of which exhibit acute, latent, and chronic Chagas disease symptoms analogous to those evidenced in human patients (Bern, 2015; Bern et al, 2011; Gunter et al, 2017; Hodo et al, 2018; Kjos et al, 2008; Malik et al, 2015; Ryan et al, 1985). Striped skunks (Mephitis mephitis) are among the wildlife taxa reported to play a role in Chagas disease ecology within North America (Brown et al, 2010; Charles et al, 2013; Galaviz-Silva et al, 2017; Hodo et al, 2018; Kramm et al, 2019; Kramm et al, 2019; Matamoros, 2016; McKeever et al, 1958; Ryan et al, 1985).
Owing to their omnivorous diet, denning habits, foraging activities, mating behaviors, and other adaptive behavioral traits, striped skunks and other New World mephitids (Mammalia: Mephitidae) are exposed to multiple routes of potential T. cruzi infection, although ingestion of infectious triatomines has been widely supported as the primary means of exposure (Ceballos et al, 2006; Charles et al, 2013; Davis et al, 1980; Gulas-Wroblewski, 2021; Jansen et al, 2015; Ribeiro et al, 1987; Roellig et al, 2009). Moreover, striped skunks are reported to exhibit adverse clinical manifestations of Chagas disease such that infection rates among imperiled North American mephitid species are of marked conservation medicine concern (Davis et al, 1980; Gulas-Wroblewski, 2021; Helgen et al, 2016; Ryan et al, 1985).
Only McKeever et al (1958) have attempted to conduct T. cruzi surveillance among a large sample size of skunks across an extensive geographic area in Georgia and Florida. Skunk surveys for T. cruzi in Texas have remained limited in both size and scope (Table 1). The risk for T. cruzi transmission is especially high in Texas, which encompasses impoverished and vulnerable communities with a history of neglected tropical diseases (Hanford et al, 2007; Hotez et al, 2012a; Hotez et al, 2012b; Sarkar et al, 2010). Texas also overlaps the endemic range of 5 skunk species, which are distributed across a wide diversity of the state's 11 ecoregions and a variety of anthropogenically influenced habitats (Gould et al, 1960; Schmidly and Bradley, 2016). Therefore, we aimed to investigate skunk involvement in Chagas disease ecology by: (1) evaluating the prevalence of circulating T. cruzi infections in Texas mephitids through PCR-based surveillance and (2) exploring factors contributing to their disease risk (i.e., sex, taxonomy, age, and location). Risk factors that we identify can inform ecological niche models to increase the precision and accuracy of predictions related to vector–host interactions and the transmission dynamics of Chagas disease across geographic and temporal scales, which can ultimately be used in public health strategy development (Gurgel-Gonçalves et al, 2012; Peterson, 2006; Peterson et al, 2002). Moreover, an understanding of skunk disease ecology based on this field survey of T. cruzi infections can assist in wildlife conservation and management planning, such as creating baseline data for population viability analyses incorporating parasite-induced morbidity and mortality estimates (Wilber et al, 2020).
Previous Reports of Trypanosoma cruzi Prevalence in North American Striped Skunks (Mephitis mephitis)
Counties are noted for locations within Texas, USA.
IFAT, indirect immunofluorescent antibody.
Materials and Methods
Ethical approval
This study was classified as exempt by the Institutional Animal Care and Use Committee (IACUC) at Texas A&M University because our methodology did not constitute “use of animals” as defined by IACUC.
Sample collection
Whole-blood (WB) specimens were opportunistically collected from living and deceased mephitids within Texas and stored on Type I Nobuto blood sampling filter paper (ADVANTEC, Tokyo, Japan) before DNA extraction. Vehicle-killed carcasses were sampled between March 2017 and January 2019. Additional samples consisted of scavenged WB derived from excess WB collected for independent purposes (e.g., genetic analyses, diagnostic procedures) by: (1) biologists researching skunk species in Texas with animal use protocols approved by the IACUC of their home institutions, (2) wildlife rehabilitators licensed by Texas Parks and Wildlife Department (TPWD), and (3) personnel at Angelo State University for Angelo State Natural History Collections' skunk biobanking project. Information related to the following factors were recorded for each individual sampled: county of origination, collection date, species, sex, and age [nonsexually mature juveniles <1 year of age; sexually mature adults 1 year of age or older as determined by morphological indicators per Crabb (1944) and Verts (1967)].
DNA extraction and PCR
Compared with serologic and culturing methods, PCR is the optimal means for detecting circulating T. cruzi during the acute phase of infection, even when immune response is low or absent (Braz, 2008; Ferreira and Borges, 2002; Gürtler et al, 1993; Jiménez-Coello et al, 2015; Jiménez-Coello et al, 2010; Jiménez-Coello et al, 2008; Kramm et al, 2019; Picka et al, 2007). PCR can also identify T. cruzi with high specificity in the more chronic phases of infection if parasites were liberated and are present in peripheral blood and/or if reinfection has occurred (Braz, 2008; Jiménez-Coello et al, 2008).
DNA was extracted from WB samples stored on Nobuto strips using the DNeasy Blood & Tissue Kit (Qiagen, MD) with the optimized DNA extraction methodology outlined in Gulas-Wroblewski et al (2021). Extracted DNA was added to a 20 μL reaction with TaqMan Fast Advanced Master Mix (Thermo Fisher Scientific, MA), then analyzed on a ViiA 7 Real-Time (RT) PCR System (Thermo Fisher Scientific). To detect T. cruzi DNA, Cruzi 1/2/3 assay as given in Piron et al (2007) was used, whereas the Actin f/r/p assay described by Piorkowski et al (2014) was used to detect β-actin. An extraction negative control, no template control (5 μL of laboratory grade H2O), and a positive control (5 μL of T. cruzi/mouse DNA sample) was included in each RT-PCR run. DNA samples were tested in triplicate for T. cruzi DNA detection and singly for β-actin detection.
Samples with cycle threshold (Ct) values <40 were interpreted as positive in the Cruzi 1/2/3 assay, and those with Ct values <38 were identified as positive in the Act.f/r/p assay. If all samples for an individual were negative based on these measures, but at least one sample was within 1 Ct unit of the cut-off point, the WB specimen was re-extracted, retested, and reanalyzed per the protocols detailed previously. Individuals were defined as positive for T. cruzi if at least one of their three DNA samples was positive in the Cruzi 1/2/3 assay. A skunk was identified as negative for T. cruzi if all three DNA samples were negative in the Cruzi 1/2/3 assay but the sample was positive in the Act.f/r/p assay.
Statistical analysis
County information for each sampled skunk was used to delineate the ecoregion and level of Chagas disease hotspot in which the individual was located. Because the ecoregions of Texas cross county lines, counties sampled for skunks were assigned to 10 ecoregions following the TPWD's deer management ecoregion assessments (TPWD, 2011). Each county was also appointed a value between 0 and 3 to represent the degree to which positive cases of Chagas disease were previously recorded in the jurisdiction. A value of one was added to a county's “Chagas disease hotspot” sum for the presence of each of the following: T. cruzi–positive triatomine vectors (2013–2018), T. cruzi–positive canine sentinels (2013–2015), and autochthonous human Chagas disease cases (2013–2018) as reported by the Texas Department of State Health Services (TDSHS, 2019).
Pearson's chi-squared tests (α = 0.05) were used to evaluate T. cruzi prevalence within each species and with all mephitid species pooled for deviations between species, sexes, ages, ecoregions, and Chagas disease hotspots. When 20% or more of the cells in the chi-squared tests had expected counts <5, likelihood ratio values (α = 0.05) were assessed. Odds ratios were also calculated for sex and age based on all skunk species and within each mephitid taxon. All statistical analyses were performed on STATA 16.1 software (StataCorp, TX).
Results
Prevalence of T. cruzi in Texas skunks
WB samples were collected and successfully extracted for DNA from a total of 235 individual skunks sampled between March 2004 and June 2019, representing 42 Conepatus leuconotus (American hog-nosed skunk), 171 M. mephitis (striped skunk), 7 Spilogale gracilis (western spotted skunk), and 15 Spilogale putorius interrupta (plains spotted skunk). No hooded skunk (Mephitis macroura) specimens were available for analysis. Overall, 42 mephitids (17.9%) tested positive via PCR for T. cruzi infection. Of these positive individuals, 30 were striped skunks, 8 were American hog-nosed skunks, 3 were western spotted skunks, and 1 was an eastern spotted skunk. Within skunk species, the resulting prevalence of Chagas disease ranged from 6.7% (S. putorius interrupta) to 42.9% (S. gracilis) (Fig. 1). The variation in T. cruzi infection between species was not statistically significant, a pattern that was consistent when both species of Spilogale were pooled for analysis and across paired species comparisons (Supplementary Table S1).

Prevalence values for circulating Trypanosoma cruzi infections in Texas skunk species.
Age variation in T. cruzi infections
All sampled skunks were definitively classed according to age (juveniles <1 year old; adults ≥1 year old) with 226 adults and 9 juveniles identified in total. Six striped skunks, two American hog-nosed skunks, and one plains spotted skunk were juveniles, of which 33.3%, 50%, and 0%, respectively, tested positive for T. cruzi infection. In comparison, adult striped skunks, American hog-nosed skunks, and plains spotted skunks exhibited Chagas disease prevalence of 17%, 17.5%, and 7.1%, respectively. With all mephitid species aggregated for analysis, juvenile skunks were ∼2.4 times more likely to test positive for T. cruzi than were adults, although without statistically significant support (Table 2). Similarly, chi-squared tests and likelihood ratio values failed to recover any significant relationship between age and T. cruzi infection in skunks overall or across species (Supplementary Table S1).
Prevalence and Odds Ratio Values for Variables Associated with Trypanosoma cruzi Infection in Pooled and Individual Species of Texas Skunk
n = total number of individuals tested.
Sex variation in T. cruzi infections
Extensive degradation and/or damage of road-killed and otherwise deceased skunks precluded the determination of sex for 115 sampled individuals (49% of the total tested). However, 41 females and 79 males were incontrovertibly identified. No western spotted skunk females were sampled, and only one plains spotted skunk female was sampled. Overall, the prevalence of T. cruzi infections in female skunks (24.4%) was slightly higher than in male skunks (21.5%), a variation that is not statistically significant (Table 2 and Supplementary Table S1). When evaluated at the species level, female striped skunks exhibited higher T. cruzi prevalence (28.1%) than males (17.6%), whereas prevalence was greater in male American hog-nosed skunks (28%) than in females (12.5%) (Table 2 and Supplementary Table S2).
Geographic variation in T. cruzi infections
Only one individual, an adult male American hog-nosed skunk that tested negative for T. cruzi, was unable to be identified to the county level. At least one skunk was collected from 76 counties, which cover all 10 of Gould's ecoregions of Texas (Fig. 2 and Table 3). T. cruzi infections were identified in skunks collected in 24 counties (31.6% of the total counties surveyed) ranging from 7.7% (Val Verde county) to 100% (Coryell, Guadalupe, and Webb counties) prevalence with an average of 44% and a median of 37% prevalence (Fig. 1 and Supplementary Table S2). When counties were clustered by ecoregion, T. cruzi–positive skunks were recorded in 6 of the 10 ecoregions with prevalence ranging from 7.1% (Rolling Plains) to 27.3% (Blackland Prairies) with a median of 20% (Table 3). We noted a weak statistical association between T. cruzi prevalence for striped skunks and location within a “Chagas disease hotspot” (likelihood ratio value of 6.75, p = 0.08). Otherwise, we did not recover any statistically significant relationship between skunk infections and county, ecoregion, or “Chagas disease hotspot” (Supplementary Table S1).
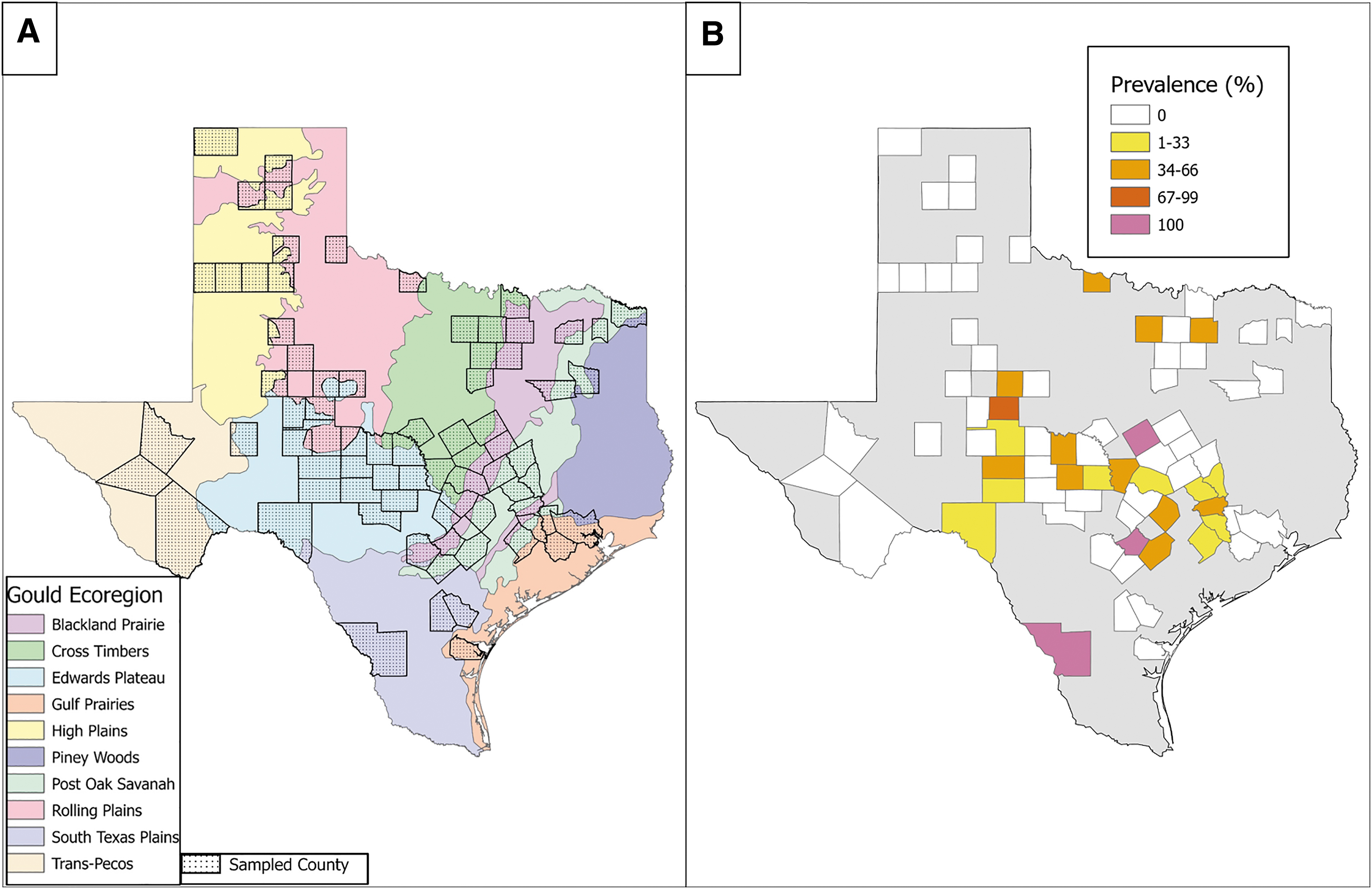
Geographic distribution of sampling locations and Trypanosoma cruzi prevalence values for skunks across Texas.
Trypanosoma cruzi Prevalence Values for Each Gould Ecoregion of Texas
Skunk species were aggregated within each ecoregion. n = total number of individuals tested.
Discussion
Our sampling of 235 individual skunks from 4 species across Texas is the most extensive T. cruzi surveillance of mephitid taxa across a wide geographic region to date. All earlier records of T. cruzi infections in North American skunks have been reported from striped skunks, with prevalence ranging between 0 and 100% and a median incidence of infection of 32%, 38%, and 100% based on PCR, culture, and serology, respectively (Table 1). Our value of 18% PCR positivity based on 171 striped skunks is substantially lower than the pooled average (48%) for the total 112 striped skunks tested before our investigation, demonstrating the imprecision of low sampling sizes for Chagas disease surveillance in this species (Table 1).
Previous evaluations of T. cruzi in members of the genus Spilogale recovered negative results for two Mexican southern spotted skunks (Spilogale angustifrons) and seven eastern spotted skunks (McKeever et al, 1958; Zavala-Velázquez et al, 1996). As such, ours are the first records of T. cruzi infections in the genus Spilogale as well as for the taxa S. gracilis and S. putorius interrupta. We also report the first incidence of T. cruzi for American hog-nosed skunks, consonant with the prevalence of infections in South American Molina's hog-nosed skunks (Conepatus chinga) and Central American striped hog-nosed skunks (Conepatus semistriatus) (Cardinal et al, 2008; Ceballos et al, 2006; de Luca D'Oro et al, 1993; Montamat et al, 1991; Pietrokovsky et al, 1991; Wisnivesky-Colli et al, 1992; Zeledón et al, 1975). The discovery of T. cruzi in plains spotted and American hog-nosed skunks is of conservation concern because both taxa have been exhibiting population declines across their endemic ranges within the United States (Meaney et al, 2006; Perry et al, 2021). Any adverse effects of Chagas disease analogous to those described for striped skunks may threaten the already imperiled plains spotted and American hog-nosed skunk populations with parasite-induced morbidity and mortality (Davis et al, 1980; Ryan et al, 1985).
Our detection of circulating T. cruzi infections in four species representing all three New World mephitid genera provided a unique opportunity to evaluate risk factors associated with Chagas disease in skunks. However, sampling bias hindered our capacity to assess variations between age classes with statistical confidence (Table 2 and Supplementary Table S1). Although not statistically significant, the trend of higher T. cruzi risk for juveniles of all skunk species pooled, striped skunks, and American hog-nosed skunks aligns with previous findings for a smaller sample of striped skunks in El Paso county, Texas as well as for domestic dogs (Canis lupus familiaris) and suburban common opossums (Didelphis marsupialis) in Mexico and laboratory rats (Rattus norvegicus) (Arce-Fonseca et al, 2017; Galaviz-Silva et al, 2017; Matamoros, 2016; Pérez et al, 2011) (Table 2). Increased disease risk in juveniles may be a function of detrimental immunoendocrine response, which promotes elevated parasitemias in younger animals (Pérez et al, 2011). Chagas disease in neonates can also be exacerbated by vertical transmission of parasites as evidenced in bats, laboratory rodents, domestic dogs, and humans (Alkmim-Oliveira et al, 2013; Andrade, 1982; Añez et al, 2009; Howard et al, 2014; Moreno et al, 2003; Rodríguez-Morales et al, 2011; Sánchez Negrette et al, 2005). Because other studies have reported more severe disease manifestations and elevated mortality in T. cruzi–positive younger animals when compared with adults, a trend toward higher infection risk in juvenile skunks holds significance for the population ecology of these species, especially concerning for taxa of conservation interest (Kjos et al, 2008; Moreno et al, 2003; Rodríguez-Morales et al, 2011).
In contrast to the age-related prevalence patterns we observed, no difference was found between juvenile and adult T. cruzi infection incidence for striped skunks in Uvalde County, Texas and in Neuva León, Mexico (Charles et al, 2013; Galaviz-Silva et al, 2017). Age bias in T. cruzi infections was also absent in populations of domestic dogs in Panama and raccoons and rodent species in south Texas (Charles et al, 2013; Pineda et al, 2011). Therefore, our inability to find a statistically significant correlation between age and T. cruzi infection risk among Texas skunks may represent the absence of any relationship between these conditions.
Similarly, our surveillance did not find any statistically significant difference in T. cruzi circulating infections between male and female skunk taxa overall, with the prevalence for both sexes varying by only 2.9% when all species were aggregated (Table 2 and Supplementary Table S1). Previous studies found no sex bias in T. cruzi prevalence among striped skunks in Bexar and Uvalde counties, Texas and in Mexico (Charles et al, 2013; Galaviz-Silva et al, 2017; Soria, 2018). The independence of host sex and T. cruzi infection dynamics has been supported by other studies in relation to parasite loads in laboratory mice, susceptibility in laboratory rats, and prevalence in populations of white-eared opossums (Didelphis albiventris) in Argentina, Virginia opossums (Didelphis virginiana) in south Texas, and domestic dogs in Panama (Pérez et al, 2011; Pineda et al, 2011; Soares et al, 2012; Wisnivesky-Colli et al, 1992; Zecca et al, 2020).
Within species, sex biases in T. cruzi risk can stem from differential parasite exposure, susceptibility, virulence, or severity of clinical symptoms related to variations in female and male immunoendocrine functions, behaviors, diets, habitat use, and/or morphology (Brown et al, 2010; do Prado et al, 1999; Galaviz-Silva et al, 2017; Lourenço et al, 2008; Majeau et al, 2020; Pinto et al, 2010; Rabinovich et al, 2001; Ruiz-Piña and Cruz-Reyes, 2002; Santos et al, 2007; Schuster and Schaub, 2001; Tartalini et al, 2011; Yabsley and Noblet, 2002; Zuk and McKean, 1996). Small sample size precluded our investigation of sex bias in T. cruzi infections among spotted skunk species; however, discernible, although not statistically significant, variations in parasite prevalence between male and female American hog-nosed and striped skunks were detected (Table 2 and Supplementary Table S1). Compared with the opposite sex, male American hog-nosed skunks and female striped skunks exhibited higher T. cruzi prevalence, an interspecific deviation that likely reflects differences in sexually divergent seasonal activity patterns among these skunk taxa.
The highest total daily movement (in terms of both activity level and distance covered) corresponds with the period(s) of greatest energetic requirements in skunks and varies between sexes (Zhang et al, 2019). For female skunks, total activity levels are greatest during the stages of lactation and young at heel, whereas males experience these during the mate-searching and intrasexual competitions of the breeding season (Ellsworth, 2016; Larivière and Messier, 1997; Zhang et al, 2019). These time periods correspond to a higher risk of exposure to T. cruzi vectors owing to both elevated activity across a greater area and increased consumption of prey, including triatomines, to meet increased energetic demand.
In Texas, the peak breeding season for male striped skunks ranges from late February through the end of March, whereas females are lactating the dependent young primarily between May through August and into early fall in the case of late-season litters (Patton, 1974; Schmidly and Bradley, 2016) (Fig. 3). Male American hog-nosed skunks participate in two main breeding seasons, exhibiting peak activity in January through March, August, and October through December (Brashear et al, 2015; Dragoo and Honeycutt, 1999; Ellsworth, 2016). The majority of female American hog-nosed skunks in Texas are lactating the young from April through late June (Bailey, 1905; Ellsworth, 2016; Patton, 1974; Schmidly and Bradley, 2016; Taylor and Davis, 1947) (Fig. 3). When these seasonal behavioral patterns are aligned with T. cruzi–prevalence values for Texas triatomines (Curtis-Robles et al, 2018), the periods of greatest potential vector exposure for striped skunk females and American hog-nosed skunk males coincide with peak parasite prevalence within triatomines. In contrast, compared with the peak activity periods for the opposite sex, peak activity periods for male striped skunks and female American hog-nosed skunks overlap seasons of lower triatomine T. cruzi positivity (Fig. 3). The effects of reproductive ecology and behavior on the prevalence of T. cruzi infection within and between skunk species deserve more attention in future research, particularly among western and eastern spotted skunk species. These spotted skunk taxa share similar timing of lactation and young rearing by females but exhibit a significantly offset breeding season owing to the presence of delayed implantation in western spotted skunks (Greensides, 1973; Kaplan and Mead, 1994; Kinlaw, 1995; Mead, 1968).
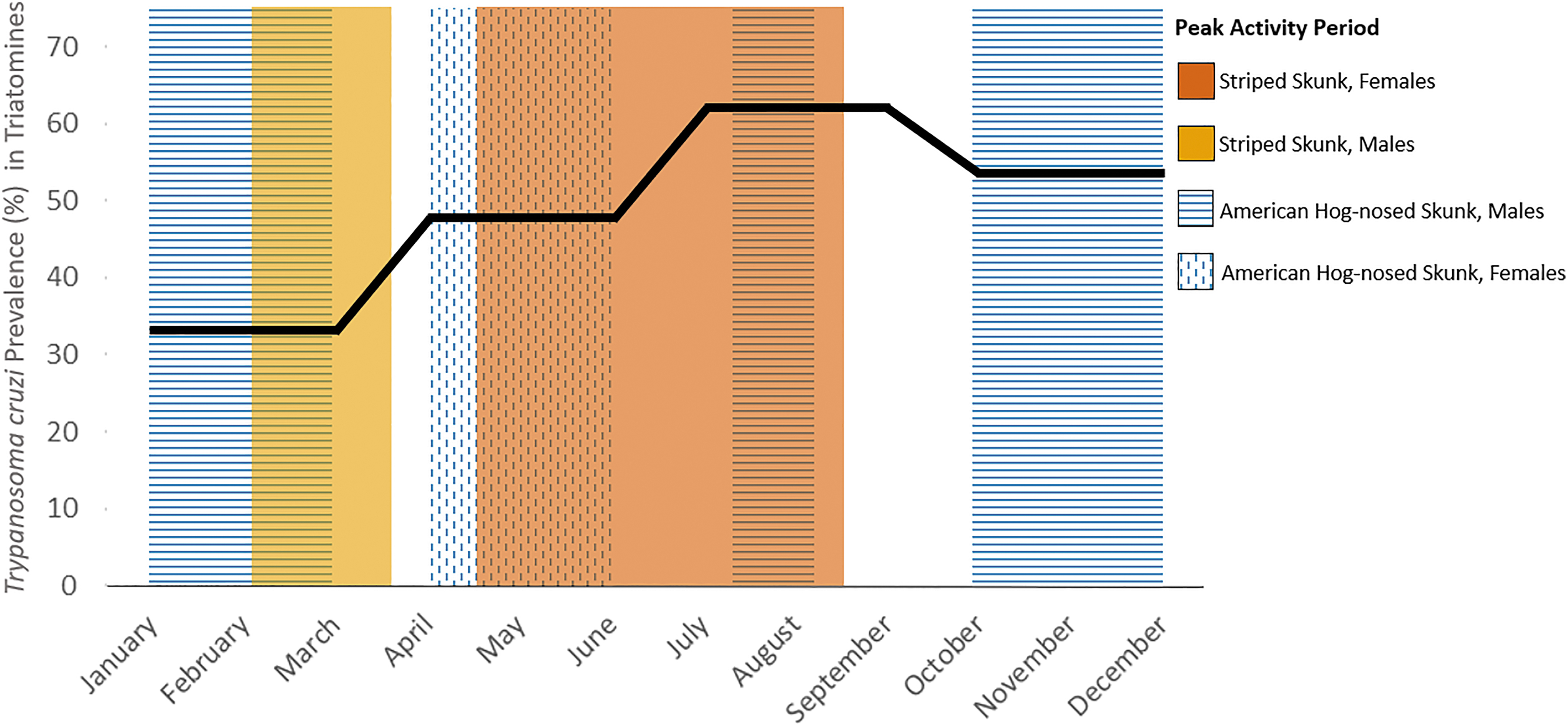
Temporal intersection of Trypanosoma cruzi prevalence of triatomine vectors (bold black line) and sexually divergent activity patterns of American hog-nosed (Conepatus leuconotus) and striped skunks (Mephitis mephitis) in Texas. Prevalence values of triatomine vectors per Curtis-Robles et al (2018). For American hog-nosed skunks, note the overlap of the males' peak activity periods in August and October to December with higher T. cruzi prevalence in triatomines compared with T. cruzi prevalence in triatomines during females' peak activity period from April to June. For striped skunks, note the overlap of the females' peak activity periods in the summer through early fall with higher T. cruzi prevalence in triatomines compared with T. cruzi prevalence in triatomines during males' peak activity period in the spring.
Conclusions
We performed the first known wide-scale and taxonomically diverse survey for circulating T. cruzi infections in skunks across Texas, reporting the first positive cases for plains spotted and American hog-nosed skunks. Although sampling biases precluded our incontrovertible assessment of the risk factors associated with T. cruzi prevalence, several trends were discernible related to age and sex biases, which can inform population and host–parasite models supporting conservation, wildlife management, and public health strategies. Future research should extend the geographic scope and number of individuals sampled, particularly in the case of spotted skunks, to further investigate the risk factors for T. cruzi in skunks and the roles these mesocarnivores play within Chagas disease transmission cycles. This study highlights the value in surveying multiple taxa of closely related wildlife species across a wide expanse of habitats to investigate the complex adaptive systems that underpin the ecology of vector-borne zoonotic diseases.
Footnotes
Acknowledgments
The authors thank the following individuals and institutions for access to and donations of skunk WB samples: the Angelo State Natural History Collections; Angelo State University Skunk Day participants; Stephanie Diaz (Wild West Wildlife Rehabilitation Center); Stephanie Brewer-Martinez; Michelle Camara (Southern Wildlife Rehab, Inc.); Diane Cook; Dove Key Ranch Wildlife Rehabilitation, Inc.; Alexandra A. Gibson; Daniel L. Heare; Melinda D. Luper, DVM; Victoria Mann; J. Clint Perkins and all of the Texas Eastern Spotted Skunk research project members; and Anton F.-J. Wroblewski.
The authors also thank the William T. Shearer for Human Immunobiology for providing the facilities and resources needed to carry out this study. Partial funding for collection of some of the skunk samples used was provided to R.C.D. by the Texas Comptroller of Public Accounts. Skunk silhouettes were provided by Anton F.-J. Wroblewski. This study was integrated into B.G.W.'s PhD dissertation, which was approved by the Department of Wildlife and Fisheries Sciences, Texas A&M University and her committee, Donald Brightsmith, Gerard Kyle, Thomas Lacher, Jr., Roel Lopez, and Amanda Stronza. The authors are also grateful to two anonymous reviewers whose comments improved the quality of this work.
Authors' Contributions
Author contributions are as follows: writing—original draft preparation, B.E.G.-W.; writing—review and editing, B.E.G.-W., R.G., R.B.K., R.C.D., and K.O.M.; conceptualization, B.E.G.-W. and K.O.M.; methodology, investigation, and analysis, B.E.G.-W., R.G., and R.B.K.; resources and project administration, K.O.M.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This research was supported in part by National Institute of Allergy and Infectious Diseases award number R03AI123650.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.