
Abstract
Background:
It is well established that infection patterns in nature can be driven by host, vector, and symbiont communities. One of the first stages in understanding how these complex systems have influenced the incidence of vector-borne diseases is to recognize what are the major vertebrate (i.e., hosts) and invertebrate (i.e., vectors) host species that propagate those microbes. Such identification opens the possibility to identify such essential species to develop targeted preventive efforts.
Methods:
The goal of this study, which relies on a compilation of a global database based on published literature, is to identify relevant host species in the global transmission of mosquito-borne flaviviruses, such as West Nile virus, St. Louis virus, Dengue virus, and Zika virus, which pose a concern to animal and public health.
Results:
The analysis of the resulting database involving 1174 vertebrate host species and 46 reported vector species allowed us to establish association networks between these species. Three host species (Mus musculus, Sapajus flavius, Sapajus libidinosus, etc.) have a much larger centrality values, suggesting that they play a key role in flavivirus community interactions.
Conclusion:
The methods used and the species detected as relevant in the network provide new knowledge and consistency that could aid health officials in rethinking prevention and control strategies with a focus on viral communities and their interactions. Other infectious diseases that harm animal and human health could benefit from such network techniques.
Introduction
Pathogens that are potentially multihost, such as Zika virus (ZIKV), Dengue virus (DENV), and West Nile virus (WNV), represent a public health threat because of the burden they impose on affected communities (Gubler, 2002). Because the virus can use a variety of hosts as refuge, it is necessary to fully characterize their life cycles, vector, and host preferences, as well as their spatial and temporal dynamics to design novel preventive and control techniques (Taylor et al., 2001). To do so, it is possible to identify relevant hosts and vectors that propagate infections inside ecosystems through a network approach based on graph theory.
In multihost transmission, the fact that the pathogen can be transmitted (or not) through different host species may affect the infectious disease transmission, as explained by the dilution effect theory where a high hosts competence richness in a species assemblage tends to decrease pathogen prevalence (Dushoff, 1996; Haydon et al., 2002; Hess, 1996; Holt et al., 2003; Roche et al., 2013; Roddam, 2001; Sober and Steel, 2017; Woolhouse et al., 2001). In other words, the diversity of competencies and distributions of hosts and pathogens regulate the disease risk and may influence interactions in communities. For this reason, it is crucial to identify hosts and vectors of multihost parasites for effective control (Taylor et al., 2001).
While there is a wide range of hosts, there is also a large array of micro- and macroparasites that are frequently overlooked, but have the same kind of ecological interactions as any living being (Mihaljevic, 2012). Current approaches typically attempt to consider interactions between a couple of parasites, but very rarely with the entire parasite diversity or parasite communities (Abbate et al., 2018; Seabloom et al., 2015), and thus all the interactions that can be found between all of them (Dallas and Presley, 2014; Johnson et al., 2015; Nieto-Rabiela et al., 2018; Rynkiewicz et al., 2015).
To understand how diverse interactions among viral communities can influence the prevalence of an infection, we must first understand which host can acquire several parasites at the same time. With this knowledge, we may characterize the interaction network and identify significant hosts and vectors that could be targeted to control infectious diseases, allowing the network to be fractured or disassembled later and therefore lower disease prevalence or impact (Holme, 2017; Luke and Harris, 2007).
In this study, we compile a global database based on a literature review to identify which vertebrate and invertebrate species are infected by a given virus, using here four viruses that affect human and animal health (Farfan-Ale et al., 2010; Fernández-Salas et al., 2007; Ibáñez-Bernal et al., 1997). Then, we utilize graph theory to model this complex system and identify the connections between host species, vectors, and viruses to find nodes with a high degree that may represent potentially interesting targets.
Methods
The Flaviviridae family is well-known to be mostly mosquito-borne viruses (Bolling et al., 2012). As multipathogen/multihost dynamics, they frequently involve a variety of vector and host species (Hoyos et al., 2021). The WNV and the Saint Louis Encephalitis Virus (SLEV) have the most reported interaction in the literature (Bolling et al., 2015; Bolling et al., 2012; Salas-Benito and De Nova-Ocampo, 2015; Weaver and Barrett, 2004). The likelihood of contact would be substantially smaller in the case of a short-term infection. In this regard, they represent an excellent example of a multihost, multivector, multipathogen system that we can use in our research.
Database
We searched for the name of the virus (“SLEV,” “WNV,” “DENV,” and “Zika”) and the words “Mammal,” “Bird,” “Reptilian,” “Amphibian,” or “Vector” and the word “detection” for example “DENV AND (mammal OR bird OR reptile OR amphibian OR vector) AND (detection)” in the Web of Science database, PubMed database and WorldWide science between the years 1990–2022. The virus, host, and vector, as well as the location and reference from which the virus was identified, were all examined to create a database with all these information.
Networks
We used the igraph package implemented in R software version 3.5.3 (R Core Team, 2017) to create three different networks (host-vector, virus-host, virus-vector), and their visualization has been done with the package “visNetwork” (Almende and Thieurmel, 2016) in the software R to detect the relevant vertebrate hosts and vectors in the multiple transmissions of flavivirus.
Host-vector network
First, we created a host-vector network to visualize all the interactions between host species. A bipartite and weighted network with the two types of nodes, vertebrate and invertebrate host (host and vector, respectively), has been constructed. When vertebrate hosts and vectors shared at least one of the four flaviviruses chosen, they were connected in the network. It is worth mentioning that this does not rule out the possibility that this interaction could be due to shared interactions with a third species not observed, but we assume the vertebrate and invertebrate species have the capability to facilitate the viral interactions inside them and disperse the virus. We ran a sensitivity test by creating a network with stronger connections (assuming a link when at least two or three viruses are shared, see Supplementary Appendix SA1), and the conclusions.
Virus-host and virus-vector network
Then, two other networks were constructed, one for hosts and one for vectors. Two types of nodes are involved in these networks: the virus and its hosts, which are connected with an edge if an infection has been reported. As a high-degree value suggests that they can host many viruses, they can present coinfections and could be essential in viral interactions (mutations, cross-antigenicity, exclusion) inside them.
Network measurements
We measured different centrality values using this approach, including degree (the number of links for each node) and betweenness (the number of times a node acts as an intermediary to connect each possible pair of nodes) (Martínez-López et al., 2009; Newman, 2014, chap. 7; Nieto-Rabiela et al., 2019). These were calculated using the algorithms included in the “IGRAPH” (Csárdi and Nepusz, 2006) and “NETWORK” packages (Butts, 2008). In the multiple transmissions of flavivirus, network-level measures were effective for summarizing the “big picture” of the network and identifying the relevant vertebrate hosts and viruses.
Results
Database
WNV, SLEV, DENV, and ZIKV (Farfan-Ale et al., 2010; Fernández-Salas et al., 2007; Guerbois et al., 2016; Ibáñez-Bernal et al., 1997) were the viruses that we selected.
We found 8697 articles and the PRISMA graphic available in Fig. 1 explain the selection papers. There were 216 articles in English that fit the requirements since 1991 until November 2022. Among these articles, hosts were isolated in laboratory experiments and were thus omitted from further analysis. There were 1318 reports of vertebrate hosts that correspond to 1174 species. Finally, the final networks involve 1174 vertebrate host species, including 982 birds, 176 mammals, 7 domestic animals, 7 reptiles, and 2 amphibians (Fig. 2). There are 46 species on vectors (40 mosquitoes and 6 mites).
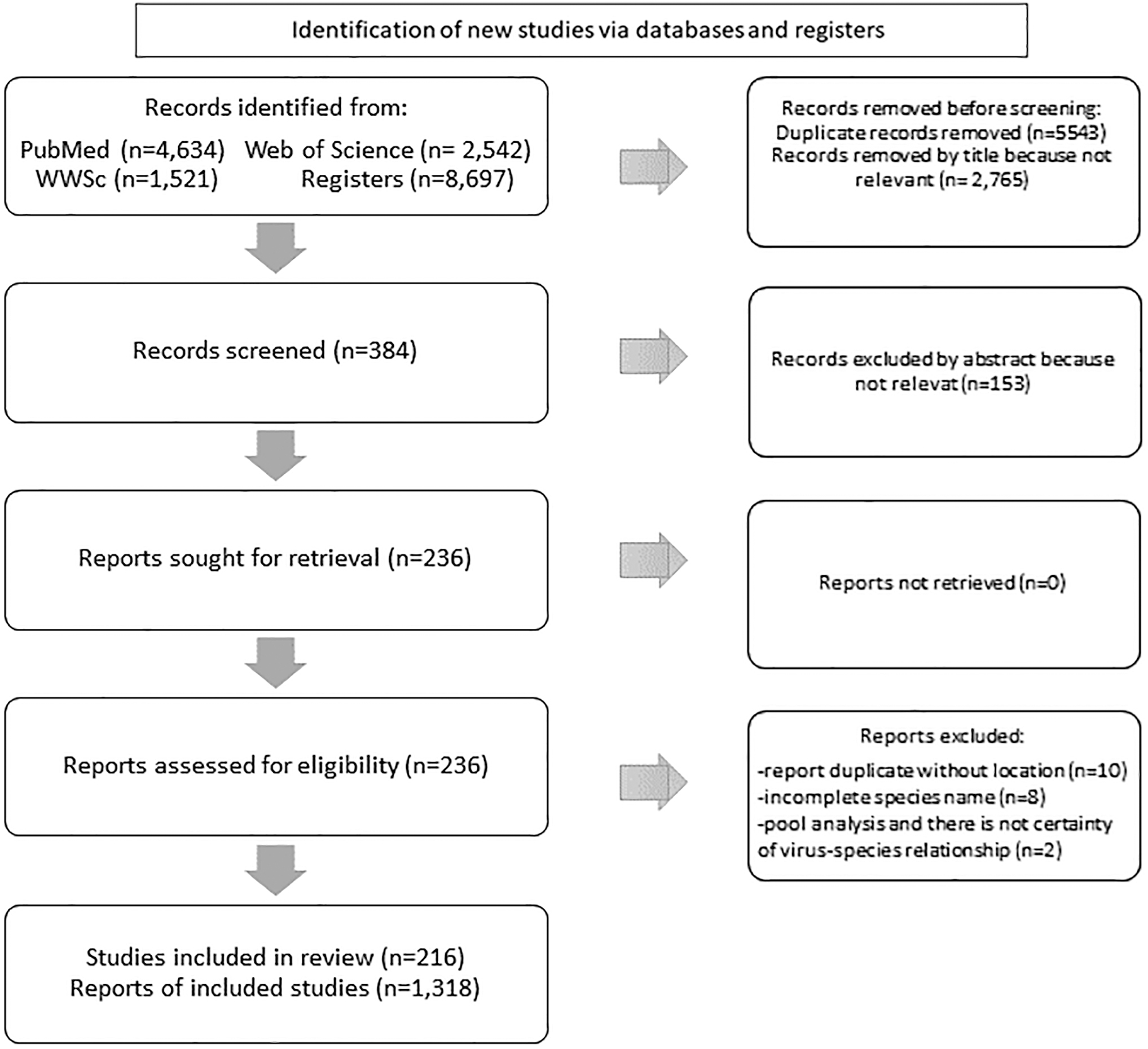
PRISMA-style chart.
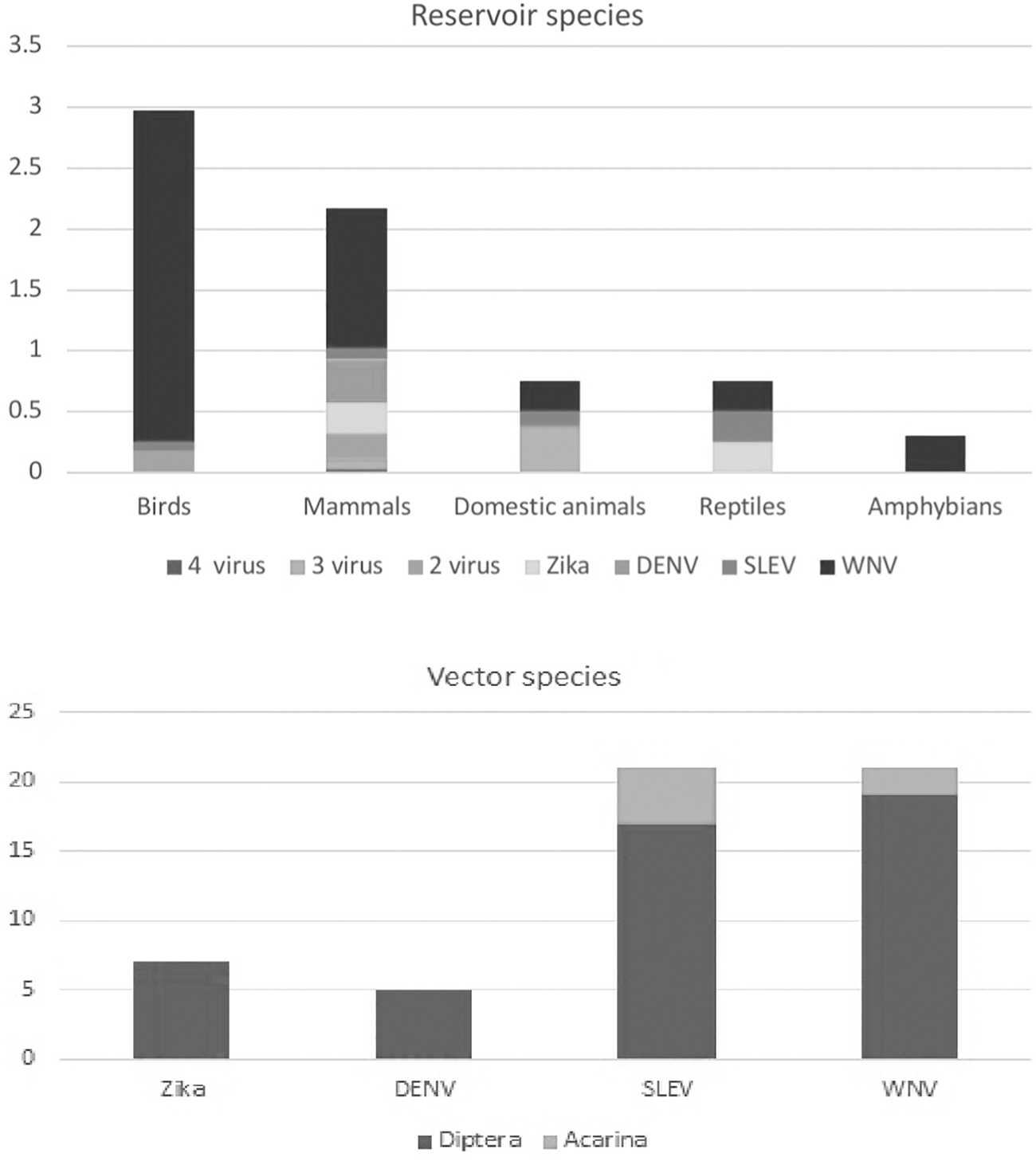
Viral occurrence frequency in hosts and vectors. The x axis shows the type of host, the y axis shows how many species have been recorded in this type of host. In log 10, the graphic depicts the frequency of hosts in each class in each viral susceptibility group.
WNV has been found in 21 mosquito vector species and 1090 vertebrate host species, with the following distribution: 966 different bird species, 119 different mammal species, 3 reptile species, and 2 amphibian species. SLEV has been found in 21 mosquito species, 3 mite species, and 110 vertebrate hosts, including 85 bird species (2 of which are domestic), 23 mammalian species (2 of which are domestic), and 2 reptile species. Seven mosquito species have been identified infected by Zika, as well as 41 mammalian species, 2 reptile species, and 3 bird species, for a total of 46 vertebrate host species. Only mammals have been reported to have DENV, and we found 41 vertebrate host species and 5 mosquito vector species.
Vertebrate host-vector network
In the Vertebrate host-vector network, we found 1222 nodes with 25,415 edges after analyzing the 1174 vertebrate hosts and 46 vectors. This is a low-density network with a diameter of 4 and a density value of 0.033 (maximum value is 1). The average degree by species was 42.46 and the average distance between them was 744.18. Table 1 shows the top five vertebrate host and vector values; the rest of the data may be found in the Supplementary Material. The diameter suggests the diversification between the viral communities (greater number of viral combinations between hosts), while the density suggests that the viral communities of the hosts are more diverse (greater number of viruses within the same host).
Top Five of Degree and Betweenness in the Vertebrate Host-Vector Network
Virus-vertebrate host network
The network with four viruses (ZIKV, WNV, SLEV, DENV) for a total of 1178 nodes and 1287 links, we analyzed 1174 vertebrate hosts and 4 viruses. With a diameter of 4, the density is 0.001. The average degree was 2.15 and the average distance was 775. Table 2 shows the top five vertebrate host and virus values; the rest of the information is available in the Supplementary Material.
Vertebrate Hosts and Viruses Top Five of Degree and Betweenness in the Virus-Vertebrate Host Network
Relevant vertebrate hosts to have viral co-occurrence
We found 86 species with the capability to host two different viruses (Supplementary Appendix SA1). Mus musculus, Sapajus flavius, and Sapajus libidinosus are the species with the capability to host the four different viruses and the last two with distribution in Brazil. The species with the capability to host three viruses were Alouatta carava, Rattus rattus, Myotis lucifugus, Artibeus jamaicensis, Equus caballus, and Sigmodon hispidus. A. jamaicensis can host WNV, SLEV, and DENV; and S. hispidus and E. caballus can host WNV, SLEV, and Zika. Results are consistent with previous research (Sotomayor-Bonilla et al., 2021).
Virus-vector network
We looked at 50 nodes spread across 46 vectors and 4 viruses (Zika, WNV, SLEV, and DENV). With a low density of 0.044 and a diameter of 6, the network includes 54 linkages. The average degree was 2.16 and the average distance between them (betweenness) was 56.9. Aedes aegypti and Aedes albopictus have higher values than Zika, implying that they play an important role in world flavivirus transmission. Table 3 shows the top five viruses and vectors in terms of degree and betweenness values, with the rest of the data in the Supplementary Material.
Vectors and Viruses Top Five of Degree and Betweenness in the Vertebrate Host Simple Network
Relevant vectors to have viral co-occurrence
Ae. aegypti and Ae. albopictus can host Zika, SLEV, and DENV, and Culex nigripalpus, Culex quinquefasciatus, Culex stigmatosoma, and Culex tarsalis can transmit SLEV and WNV.
Discussion
In this study, we discovered that only 4 virus species from the same family are linked with more than 1000 host species and 30 vector species. We have been able to identify different species presenting unusual characteristics in terms of the network metrics, suggesting that they can play a larger role than other species in viral transmission. Our main important result shows that C. nigripalpus is ranked first in the central values of the vertebrate host-vector network, and then third in the centrality values of the virus-vector network. This is because C. nigripalpus has a large number of vertebrate hosts, the vector is more important among the three components because it connects them all. Therefore, our results show that the control focus should be on C. nigripalpus in American continent, but in a global level C. quinquefasciatus.
In our network, the centrality values were higher in rodents and bats than in the other host species, except in one bird species (Bubulcus ibis). They are all considered invasive or generalist species with a wide range of distribution (Fowler and Cubas, 2001, chap. 9). They are potentially significant species in transmission because they may host a variety of viruses, have a high rate of interaction with host species, and can transfer viruses (Becker et al., 2018; Egoscue, 1976; Erazo et al., 2019). Regarding vectors, mites are not relevant for viral transmission, according to their low centrality values, and some investigations imply that mites are not always able to complete the cycle for some infections (Whitaker et al., 2009). As there is not much information about them, this judgment may be limited. On the opposite, Culex spp. are competent vectors, as shown in Fig. 3, because they have a greater impact on virus dissemination and are linked to viruses like WNV and SLEV, which have many vertebrate hosts recognized (Crockett et al., 2012; Newman et al., 2011).
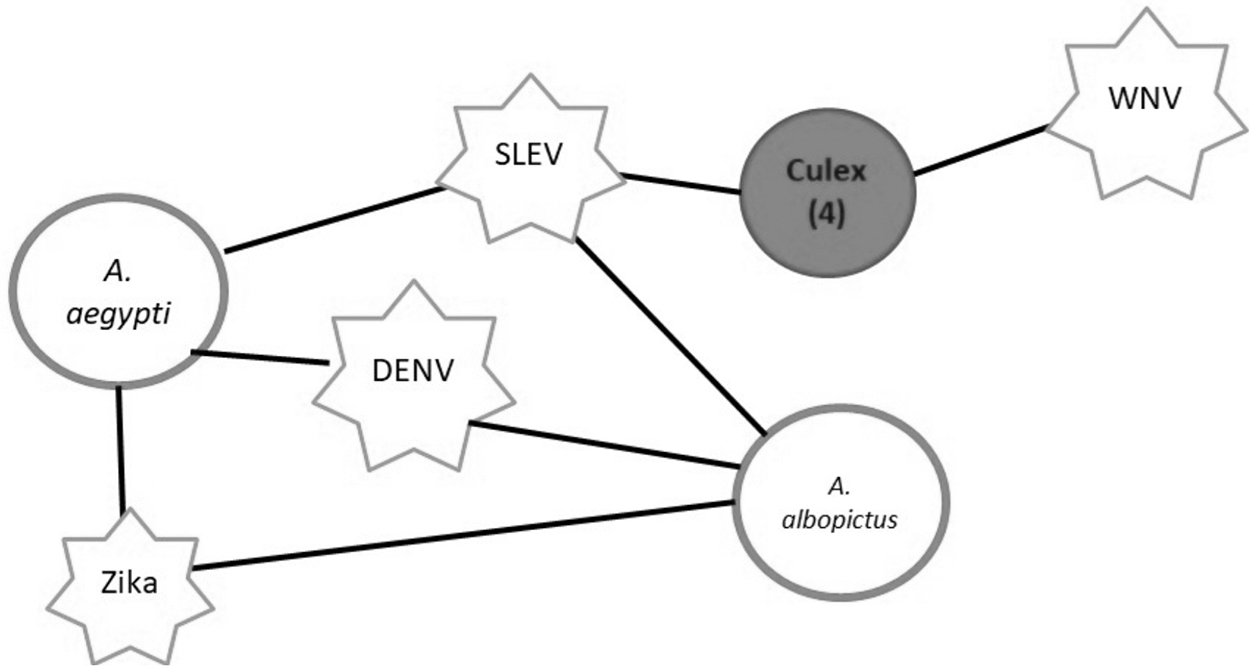
Relevant vectors Network. Viruses are represented by star, vectors by circle, and associations by connections. C. nigripalpus, C. quinquefasciatus, C. stigmatosoma, and C. tarsalis are represented by the gray circle.
Our study suffers of several caveats that need to be discussed. Due to the amount of the available data, the study was conducted at a worldwide scale rather than at a local one. It is therefore important to be careful with a potential local interpretation of the results. But to fully understand the system, modeling potential networks are required, rather than relying solely on published data that provide results for the local reality (Cooper et al., 2012). In this case, we assume a link between two host species when a virus infects two host species, despite the fact that they could have acquired the virus through interaction with a third host species.
We performed a sensitivity analysis with a network where we assume a link only if two viruses were shared by two hosts and our conclusions remain consistent (diameter of 4 and a density value of 0.020). We also want to reinforce that in the network, we are not saying that a link is a direct interaction, much less a physical one, but rather a vector/host in which a greater number of the flaviviruses that we select in our research transit. Therefore, these bases are a point of interest to understand, study, and monitor the existent of real transit through them, either with the help of more species between them. If there is communication, mutation or viral changes in one of these hosts/vectors could reach the other in a matter of a few years without us realizing it.
The next step is elucidating the information of the relevant nodes suggested by the betweenness of the network. The interesting point is to locate which host species can serve as a bridge between viruses. The importance of these bridges can be revealed by the number of connections associated to each of these host species (Pastor-Satorras et al., 2014). We mean that if a host has the capacity to host the four viruses, it is more likely that viral mutations occur in those hosts or that these hosts can help us in epidemiological control by propagating attenuated viruses, with crossed antigenicity or excluding the problem virus of the virus. However, this requires gathering more data of positive and negative hosts at a local scale to characterize more precisely these networks and offer a surveillance/control opportunity.
Meta-analyses give us the advantage of processing a larger amount of data with a low budget and finding and rediscovering patterns on a large scale. But it always carries several biases since the researcher does not have control over the method of collecting information in the field as it would be in their own investigation. However, we can apply inclusion and exclusion criteria trying to modify these biases. In our investigation, we work with the information available, and it should be updated. We did not find information on true negatives that would clarify “ditches” between species. But research biases are great opportunity niches in future research. There is also a temporal bias since the samplings have been conducted at different times. To minimize the temporal bias, we assume the capacity of the host species to host the virus, but we do not take current circulation into account.
Another bias is the oversampling and the undersampling biases in different regions, so we do not take their location into account. We cannot detect global patterns using our network, because the American continent (America) contains more species that share viruses than the other continents by oversampling. In contrast, we found only two flaviviruses of the four reported in Europe, and both viruses share a nonvertebrate host. To fully comprehend the system, extrapolation and modeling potential networks are required, rather than relying solely on published with geographical information data that provide results for the spatial-temporal reality (Cooper et al., 2012). Our objective is to locate the appropriate species in multiple transmissions focusing on viral communities inside the host, we prefer to apply prospective thinking in this scenario.
Conclusion
Flaviviruses are a large group of viruses that have a significant impact on human and animal health. Understanding the interactions between various vertebrate and invertebrate hosts and viruses can provide us with the information we need to make better decisions. So that these findings may help us develop control measures to control some flaviviruses, as well as other viruses, bacteria, parasites, or organisms in more complicated epidemiological situations.
Footnotes
Acknowledgments
We are grateful to CONACyT and l'Ambassadede France au Mexique. We also thank CONACyT PRONAII 5 project—SOCIO-ECOSISTEMAS Y VIRUS EMERGENTES—(No. 303002, “DIVERSIDAD BIOLÓGICA, SOCIO—ECOSISTEMAS Y ENFERMEDADES VIRALES EMERGENTES EN MÉXICO”). Special thanks to Anuwat Wiratsudakul to encourage me.
Authors' Contributions
F.N.-R.: conceptualization, data curation, formal analysis, investigation, methodology, and writing—original draft. F.E.: methodology. B.R.: supervision and validation writing—reviewing and editing. G.S.: supervision and writing—reviewing and editing.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
Consejo Nacional de Ciencia y Tecnología (CONACyT) Proyectos Nacionales de Investigación e Incidencia (RONAII) 5 project— SOCIO-ECOSISTEMAS Y VIRUS EMERGENTES—(No. 303002, “DIVERSIDAD BIOLÓGICA, SOCIO— ECOSISTEMAS Y ENFERMEDADES VIRALES EMERGENTES EN MÉXICO”).
Supplementary Material
Supplementary Material
Supplementary Appendix SA1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.