
Abstract
Background:
The dengue, Zika, and Chikungunya arboviruses have spread in America in the past year, thus becoming global health issues. These viruses are maintained in nature in two transmission cycles: an urban cycle, transmitted from hematophagous mosquitoes to humans, and a wild cycle, recorded only in Africa and Asia, involving mosquitoes and nonhuman primates as natural hosts. The evidence shows that these arboviruses infect other wild mammals in America, such as rodents, marsupials, and bats. This study aimed to determine the potential natural infection of arboviruses in bats captured in contrasting sites (tropical forests, urban areas, and caves) in Oaxaca, Mexico.
Materials and Methods:
Liver samples were collected from some bats and tested for RNA from dengue, Zika, and Chikungunya with the quantitative real-time PCR assay. We analyzed 162 samples that encompassed 23 bat species.
Results:
No natural infection with any of the three arboviruses was detected in any sample tested.
Conclusion:
The existence of a wild cycle of the three arboviruses in the American continent is not ruled out. However, owing to the low or zero prevalence recorded in other studies and the present study, bats are likely involved in the arbovirus transmission cycle as accidental hosts.
Introduction
Arboviruses (arthropod-borne viruses) are a group of viruses transmitted by arthropods and hematophagous insects, such as mosquitoes and ticks (WHO, 1967). This group comprises ∼500 viruses of the families Bunyaviridae, Flaviviridae, Rhabdoviridae, Reoviridae, and Togaviridae (Tsai, 1999). About 150 arboviruses cause disease in humans characterized by arthritis and rash (e.g., Mayaro virus and Chikungunya virus [CHIKV]), encephalitis (St. Louis and Nile viruses), fevers and myalgia (dengue), or hemorrhagic fevers (dengue and yellow fever) (Arredondo-García et al., 2016; Navarrete-Espinosa and Gómez-Dantés, 2006). The dengue virus (DENV), Zika virus (ZIKV), and CHIKV have spread across American countries in the past years, thus becoming a global public health issue (Patterson et al., 2016).
The DENV and ZIKV Flaviviruses belong to the family Flaviviridae, whereas CHIKV is an Alphavirus of the family Togaviridae (International Committee on Taxonomy of Viruses, 2023). DENV, which causes dengue fever and dengue hemorrhagic fever, is endemic to many countries in America and one of the most important re-emerging vector-borne diseases in North America (WHO, 2022). As surveillance and prevention of these viruses involve monitoring their vectors, poor control of the latter, together with accelerated population growth, migrations, national and international travel, and changes in ecosystems have fostered their expansion in tropical and temperate regions (Gaunt et al., 2001; Vasilakis et al., 2011). In 2022, >2.7 million cases of disease from the three arboviruses have been reported in America, with 89.96% corresponding to dengue, 8.96% to Chikungunya, and 1.08% to Zika (OPS, 2022).
The three arboviruses are maintained in nature through two transmission cycles facilitated mainly by mosquitoes of the genus Aedes (Aedes aegypti and Aedes albopictus). The urban cycle is well-documented in the Neotropics and involves humans as natural hosts (Althouse et al., 2016; Weaver and Vasilakis, 2009; Weaver et al., 2016). A second cycle—the wild cycle—recorded only in Africa and Asia involves nonhuman primates as natural hosts. However, there is serological and molecular evidence that these arboviruses infect other wild mammals in America, such as rodents, marsupials, and bats. Hence, there is increasing interest in the search for potential wild reservoirs (de Thoisy et al., 2009; Perea-Martínez et al., 2013).
Bats are characterized by wide distribution and high diversity, with >1400 recognized species worldwide (Simmons and Cirranello, 2022). Besides, the order Chiroptera harbors a higher viral richness than other groups of mammals owing to their unique and sophisticated immune system, the ability to fly, gregarious habits, and longevity relative to their weight and size (Calisher et al., 2006; Hayman, 2016; Olival et al., 2017).
In America, bats' natural and experimental arbovirus infections have been better documented for DENV than ZIKV and CHIKV. Bat susceptibility to ZIKV has been investigated primarily in Africa, the continent where the virus originated (Dick et al., 1952; Shepherd and Williams, 1964; Simpson and O'Sullivan, 1968). In America, experimental studies of ZIKV infection have been conducted in Myotis lucifugus and Artibeus jamaicensis (Malmlov et al., 2019; Reagan et al., 1955), and Torres-Castro et al. (2021) reported the first cases of natural ZIKV infection in A. jamaicensis in Yucatan, Mexico. In addition, CHIKV has been inoculated in Eptesicus fuscus in the United States, and antibodies against it have been detected in A. jamaicensis and Artibeus lituratus of Grenada (Bosco-Lauth et al., 2016; Stone et al., 2018) but there are no reports of natural CHIKV infections in wild bats.
However, viral RNA and antibodies against DENV have been identified in 19 bat species in America (Hernández-Aguilar et al., 2021a). In Mexico, positive cases have been reported in A. jamaicensis, A. lituratus, Carollia sowelli, Desmodus rotundus, Glossophaga mutica, Myotis nigricans, Natalus mexicanus, Pteronotus mesoamericanus, and Sturnira parvidens in the states of Veracruz, Colima, Jalisco (Aguilar-Setien et al., 2008), Yucatan (Machain-Williams et al., 2013), Morelos (Perea-Martínez et al., 2013), Campeche, Chiapas (Sotomayor-Bonilla et al., 2014), and Hidalgo (Abundes-Gallegos et al., 2018). As of 2022, there is no evidence of any reports of infections with any of the three arboviruses in bats inhabiting Oaxaca.
Therefore, the objective of this study was to determine, through quantitative real-time PCR (qRT-PCR), the potential natural infection with DENV, ZIKV, and CHIKV in bats captured in sites with different forest cover and land use (medium semi-evergreen tropical forest, low deciduous tropical forest, urban areas, and caves) in Oaxaca, Mexico.
Materials and Methods
Study area
This study was carried out within the range of the nine bat species that host the DENV in Mexico (Hernández-Aguilar et al., 2021b). Two municipalities of the state of Oaxaca, Mexico, differing in population size and the percentage of area covered by urban areas, agriculture, and tropical forests were chosen. The first locality was San Miguel Lachiguiri (hereafter Lachiguiri), belonging to the municipality of Santiago Lachiguiri, which harbors ∼500 inhabitants. It is located 550 meters above sea level, with a predominantly warm subhumid climate: minimum and maximum annual temperatures of 14°C and 24°C, respectively, and mean annual precipitation of 1500 mm (National Institute of Statistics, Geography, and Informatics, 2020). The second municipality, Santa Maria Huatulco (hereafter Huatulco), was home to 50,862 inhabitants as of 2020. It is located 220 meters above sea level, and the prevailing climate is warm and subhumid with rains in summer.
The mean annual temperature is 28°C, and the mean annual precipitation is 1.090 mm (National Institute of Statistics, Geography, and Informatics, 2020), and it receives ∼730,000 tourists annually (93% are nationals and 6.9% are foreigners; State Information Office for Sustainable Rural Development, 2016).
The sampling design included variables that contribute to explaining the variation in the detection of DENV in bats in America (Hernández-Aguilar et al., 2021a): mean annual temperature, mean annual precipitation, human population size, and collection site (urban areas, forest cover, and caves). In each municipality, a circular landscape with a radius of 5.5 km (9503.34 hectares) was selected from an urban center (Fig. 1a, b). Bats were captured in 47 sites (24 in Huatulco and 23 in Lachiguiri) spread in areas covered by tropical forest (medium semi-evergreen forest and low deciduous forest), urban settlements, and caves. In Huatulco, 11 sites are located in the urban area, 11 in tropical forests, and 2 in caves; in Lachiguiri, 9 sites are located in the urban area, 12 in medium semi-evergreen forests, and 2 in caves (Fig. 1a, b).
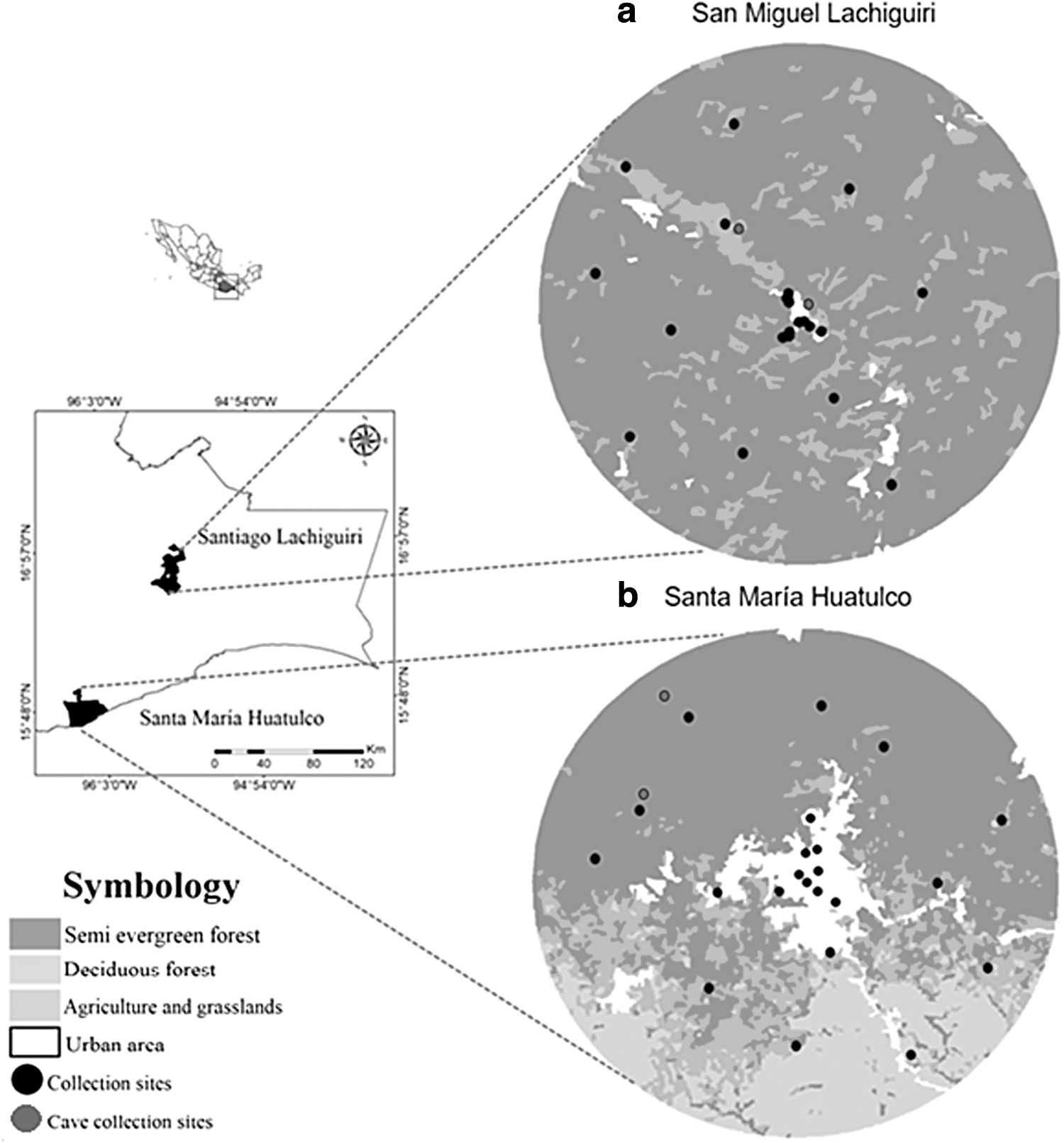
Geographic location of the Santiago Lachiguiri and Santa Maria Huatulco municipalities and collection sites in
Fieldwork
Bat collection
The sampling design included temporality as a variable. To this end, 18 samples were carried out between January, February, November, and December 2020 and from January 2021 to February 2022. Three dry seasons (October–April) and one rainy season (May–September) were included. Each monthly sampling lasted six nights. Bats were captured in urban sites and forests using three 2 × 6 meters and two 2 × 12 meters mist nets; in caves, a Bat Harp Trap—Austbat (Bat Conservation and Management, Inc., Carlisle, PA) was placed at the entrance of each cave from 18:00 to 23:30 hours. Captured bats were individually placed in 15 × 25 cm cotton bags.
The following measurements were recorded from each individual: total length, tail length, hind foot length, ear length, and forearm length, in millimeters, and weight, in grams. In addition, we recorded the sex and reproductive condition (female: pregnant, nursing, inactive; male: scrotal testes, inactive) (Kunz, 1996). Bat species were identified with the keys of Medellín et al. (2008), and Álvarez-Castañeda et al. (2017). Of all the captured bats, 96.05% were released at the capture site (n = 3938) without taking any sample for virus diagnosis. Only a small percentage (3.95%, n = 162) was killed to take liver samples for laboratory processing for virus diagnosis. Proper bat management followed the guidelines recommended by Sikes and Animal Care and Use Committee of the American Society of Mammologists (2016) for handling wild animals.
Bats to be killed were first sedated with 2% isoflurane inhalations for 2 min and then killed with an intracardiac injection of 0.2 mL sodium pentobarbital. Liver samples were collected and stored in 2.0 mL cryogenic vials (Corning Inc., Corning, NY) containing 1 mL of RNAlater ™ (Sigma-Aldrich Chemical) stabilizing solution. Each vial was labeled with the data of the collected specimen and kept refrigerated for subsequent molecular analyses. All liver specimens and replicates were collected under the scientific collection license FAUT-0143 granted to Dr. Consuelo Lorenzo and deposited in the Collection of Mammals of El Colegio de la Frontera Sur, San Cristobal de las Casas, Chiapas, Mexico. The collection catalog numbers of each specimen are given in Supplementary Table S1.
Molecular analysis
The tissue was homogenized (TissueRuptor; Qiagen) with 1 mL of viral transport medium. Viral RNA was extracted from the supernatant with the QIAamp® Viral RNA Mini Kit (Qiagen) following the manufacturer's instructions. The RNA template used for the PCR was kept at −80°C. Arboviruses were identified using the qRT-PCR assay using the protocol of Johnson et al. (2005) for DENV, Lanciotti et al. (2007) for CHIKV, and Lanciotti et al. (2008) for ZIKV. The reaction mixture included 12.5 μL of 2 × PCR Master mix, 5.45 μL of water, 7.5 μL of primer mix, 0.5 μL of Superscript III RT/Platinum® Taq Mix, 5 μL of template, and 0.045 μL of the probe for each arbovirus serotype (Table 1).
Sequences of the Oligonucleotides and Probes Used in the Quantitative Real-Time PCR Assay for the Detection of Arbovirus RNA in Bats in Oaxaca, Mexico
DENV-1 (ID. MT929160. InDRE), DENV-2 (ID. MT899085. InDRE), DENV-3 (ID. MW308179. InDRE), DENV-4 (ID. OM417339. InDRE), CHIKV (ID. KP795107.1 InDRE), and ZIKV (ID. KU686218.1 InDRE) donated by the Institute for Epidemiological Diagnosis and Reference, Mexico (InDRE) were used as positive controls and Molecular Biology Grade Water was used as a negative control (Qiagen). The qRT-PCR protocol was performed on an ABI FAST 7500 (Applied Biosystems) thermal cycler under the following conditions: reverse transcription (RT) at 50°C for 30 min, RT inactivation at 95°C for 10 min, then fluorescence detection including 45 cycles at 95°C for 15 s, and finally annealing at 60°C for 60 s. All the analyses were carried out in the Virology and Molecular Biology Laboratory of the Laboratorio Estatal de Salud Pública (State Public Health Laboratory) of the state of Oaxaca (LESPO), in the city of Oaxaca, Mexico.
Results
Overall, 4100 individuals from 23 bat species were captured: 2605 bats in Huatulco and 1495 in Lachiguiri (Table 2). The most abundant species were Mormoops megalophylla (33.9% of total captures, n = 1391), Pteronotus fulvus (24.2%, n = 995), and P. mesoamericanus (20.4%, n = 840). In general, 15 species were recorded in caves, 19 in forests, and 11 in urban areas. Of these, 43.5% were frugivorous, 26.1% insectivorous, 17.4% nectarivorous, 8.7% omnivorous, and one hematophagous species (4.3%) (Table 2).
Diversity, Abundance, and Trophic Guild of Bats Captured in Santa Maria Huatulco and San Miguel Lachiguiri, Oaxaca, Mexico During the Sampling Period
C, cave; Fr, frugivore; He, hematophage; In, insectivore; LDTF, low deciduous tropical forest; MSETF, medium semi-evergreen tropical forest; Ne, nectarivore; Om, omnivore; SMH, Santa Maria Huatulco; SML, San Miguel Lachiguiri; U, urban area.
A total of 162 liver samples distributed in the 23 captured species were analyzed, 106 samples corresponded to males and 56 to females (Table 3). The particular data for each individual are given in Supplementary Table S1. Of the 162 samples analyzed, 70.3% corresponded to phyllostomid bats, 24.1% to species of the Mormoopidae family, 2.5% to Vespertilionidae, and 3.1% to Natalidae. A greater number of samples was analyzed for the most abundant species during the captures, such as A. lituratus (n = 28), A. jamaicensis (n = 27), G. mutica (n = 18), Carollia perspicillata (n = 16), and P. mesoamericanus (n = 11). For 10 (43.4%) of the 23 less abundant species considered in the analyses, a single tissue sample was analyzed (Table 3).
Bat Species, Sex, and Number of Samples Tested for Viral RNA of Dengue, Zika, and Chikungunya in Santa Maria Huatulco and San Miguel Lachiguiri, Oaxaca, Mexico
ST, samples tested.
The number of samples analyzed from bats captured in forests, urban areas, and caves was 53, 54, and 55, respectively. All liver samples tested by qRT-PCR were negative for all three arboviruses. No sample amplified any dengue, Zika, or Chikungunya viral protein (Fig. 2).

qRT-PCR amplification plots for positive controls and unamplified samples for
Discussion and Conclusions
At present, the arboviruses ZIKV, CHIKV, and mainly DENV represent the most important diseases for public health in tropical and subtropical countries (Patterson et al., 2016). The global incidence of dengue increased in the past decade, and currently, about one-half of the world's population is at risk of infection (WHO, 2022).
The diseases caused by the three arboviruses occur in tropical and subtropical climates, mainly in urban and peri-urban areas (WHO, 2022). In this study, we covered sites with different forest covers and land uses (forests, urban areas, peri-urban areas, sites adjacent to crops, pastures, and caves) to assess whether the collection site is a variable that may explain the incidence of arboviruses in the different bat species. However, no bats were infected with any of the three arboviruses studied at the capture time. Although it is true that the sample size of each bat species may have influenced the probability of finding a natural infection (only a single tissue sample was analyzed for 43.4% of the species), the number of samples analyzed for the more abundant species, such as A. lituratus (n = 28) and A. jamaicensis (n = 27), was higher than those analyzed in other studies that reported positive results.
For instance, Torres-Castro et al. (2021) analyzed samples from 22 individuals of A. jamaicensis and reported 2 samples positive for ZIKV but none positive for DENV. With a sample size (n = 29) similar to the one tested in this study, Sotomayor-Bonilla et al. (2014) reported one A. jamaicensis individual infected with DENV in Mexico. In contrast, other studies, such as Cabrera-Romo et al. (2016), analyzed a greater number of samples (n = 240) than our study and also found no molecular evidence of arboviruses.
The species, colony size, mean annual temperature, mean annual precipitation, human population size, and bat collection site (urban area, forest cover, and caves) contributed to explaining the variation in the detection of DENV in bats in America (Hernández-Aguilar et al., 2021a). Based on the above, the sampling design of this study included all the variables just mentioned to increase the probability of finding a bat infected with any of the three arboviruses. We included samples of 23 bat species captured in sites with different forest cover and land use (tropical forests, urban areas, caves) in 2 municipalities with different population sizes and covering 4 seasons over 2 years (3 dry seasons and 1 rainy season); however, we did not get positive test results of natural infection.
The prevalence of arboviruses in bats may be influenced by other factors, such as the organs being examined. This study tested the bat liver to detect an infection with DENV, ZIKV, and CHIKV. However, these arboviruses and other flaviviruses such as the West Nile virus have shown a higher viral load in human urine and saliva for a longer time than in serum (Gourinat et al., 2015; Malmlov et al., 2019). Therefore, screening tests in bat urine and saliva samples may be a better option in future studies. Although knowledge of natural arbovirus infections in bats has grown in the past decade (Hernández-Aguilar et al., 2021a, Malmlov et al., 2019), the impact of these viruses on bat populations is still unknown.
For example, if ZIKV were fatal in bats, molecular detection of the virus would be further complicated (Gourinat et al., 2015; Malmlov et al., 2019). In this respect, serology represents an alternative to identifying past infections with these arboviruses, as reported in some studies in America (Machain-Williams et al., 2013; Platt et al., 2000; Sotomayor-Bonilla et al., 2014).
The low prevalence of arboviruses indicates that bats may be exposed to these viruses in different geographic areas (Calderón et al., 2019). However, the absence of DENV, ZIKV, and CHIKV in some studies (Bittar et al., 2018; Cabrera-Romo et al., 2016; Cabrera-Romo et al., 2014; Sotomayor-Bonilla et al., 2018) and this study supports the hypothesis that bats may not be major hosts in the maintenance of arboviruses in their wild cycle, and that the role of this group in this cycle would be as accidental hosts (Kading and Schountz, 2016). In addition, future studies should evaluate the IgG antibodies against the DENV, ZIKV, and CHIKV arboviruses that help to have an idea of the prevalence of these arboviruses in Neotropical bats.
The existence of a wild cycle for the three arboviruses in America is not ruled out. Modeling studies support the above and state that the establishment of this cycle will depend on the size of the host and vector populations, the host birth rates, and the probability of infection with the virus (Althouse et al., 2016).
This study is a baseline for further studies. ZIKV showed testicular tropism in A. jamaicensis (Malmlov et al., 2019), which may alter male fertility and, thus, affect bat populations. Future experimental inoculation studies should aim at identifying the organs most affected by arboviruses. Studies of natural infections in bats have reported the absence of signs of arbovirus infections. This finding may be owing to the sophisticated immune system of bats, which allows them to tolerate infections that are most deadly in other species without showing signs of the disease caused by the virus (Weinberg and Yovel, 2022).
Bats have been mentioned as secondary food sources for mosquitoes (González-Salazar et al., 2017; Kading and Schountz, 2016), but the transmission routes of arboviruses to bats are still unknown. It has been suggested that bats could be infected by mosquito bites or by consuming infected mosquitoes (Abundes-Gallegos et al., 2018; Cabrera-Romo et al., 2014; Sotomayor-Bonilla et al., 2018; Vicente-Santos et al., 2017). Future studies will help to better understand the routes of infection in bats. In addition, a criterion to consider in future studies is to carry them out in areas with differences in the prevalence of dengue, Zika, and Chikungunya cases in humans (e.g., areas with thousands of cases, others with dozens of cases, and areas with no registered cases) and relate this with the infection in bats.
Finally, we recommend that natural infection, experimental inoculation, and serological studies be conducted in other wild animals, such as nonhuman primates, rodents, raccoons (Procyon sp.), and armadillos (Dasypodidae), to understand the role of each group in the wild cycle of arboviruses in America (Althouse et al., 2016; Malmlov et al., 2019).
Footnotes
Acknowledgments
The Colegio de la Frontera Sur and the Instituto Politécnico Nacional, campus CIIDIR-Oaxaca, provided logistical support for the drafting of the article. Fieldwork was covered with funds from the project 320315 supported by CONACyT in 2022 granted to C.L. and SIP 20210956 to A.S.-M. The Consejo Nacional de Ciencia y Tecnología (CONACyT) awarded a fellowship grant for the first author's postgraduate studies. The authors thank the field guides, the people who lent us their homes, farmland, and forest for bat sampling. The authors thank all the volunteers who assisted in the sampling and J. Bolaños Citalán and D. Castañeda for their support in logistics before field trips. The authors thank J. Pérez Torres, C. Mac Swiney González, and L. Ruiz Montoya for their comments to improve the article. The authors also thank the staff of the Laboratorio Estatal de Salud Pública of Oaxaca, who supported the molecular analysis of arboviruses. M.E. Sánchez-Salazar translated the article into English.
Authors' Contributions
All authors confirm their participation in the preparation of the article, and all approved the final version of the article. I.H.-A. carried out the field and laboratory work, data analysis, and drafted the article. C.L., A.S.-M., and E.J.N. wrote and reviewed preliminary versions of the article. L.R.R.-P. performed the laboratory analysis and interpretation of the data and reviewed preliminary versions of the article.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
The Consejo Nacional de Ciencia y Tecnología de Mexico awarded a scholarship for postgraduate doctoral studies to the first author. Fieldwork was covered with funds from the project 320315 supported by CONACyT in 2022 granted to C. Lorenzo and SIP 20210956 to Antonio Santos Moreno.
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.