
Abstract
Background:
Bartonella species and Borrelia burgdorferi sensu lato (Bbsl) are emerging zoonotic pathogens. The vectors and frequency of infections with both pathogen groups in the southern United States is understudied. This study describes an investigation of Bartonella and Bbsl in yellow flies collected at a residence in northeast Florida, USA, that led to subsequent discoveries of both organisms in lone star ticks (Amblyomma americanum) and a human patient.
Materials and Methods:
DNA samples from flies, ticks, and human patient blood specimens were tested via polymerase chain reaction assays for Bartonella or Bbsl species. DNA sequences were compared to reference strains for identification and characterization.
Results:
An exploratory investigation of arthropod-borne pathogens in yellow flies collected at a residence in northeast Florida revealed the presence of uncharacterized Bartonella species DNA sequences similar to ones previously detected in two lone star ticks from Virginia. Subsequent testing of several lone star ticks from the area detected similar sequences of Bartonella in three ticks. Testing of stored blood samples from a resident of the site, who had experienced chronic relapsing and remitting symptoms for over a decade, identified nearly identical Bartonella DNA sequences in multiple samples collected over a 10-year period. Two lone star ticks and several samples from the same patient and time period also tested positive for Bo. burgdorferi DNA, suggesting possible long-term coinfection of the patient with both organisms.
Conclusion:
This investigation identified highly similar Bartonella DNA sequences in yellow flies, lone star ticks, and a human patient in northeast Florida. Similarly, Bo. burgdorferi DNA was detected in two lone star ticks and multiple specimens from the patient. Positive PCR results from archived patient blood samples documented the presence of both organisms at multiple time points over more than a decade. More studies on human patients with chronic undefined illness and on the presence of Bartonella and Bbsl in hematophagous arthropods and animal hosts in the southeastern United States are needed.
Introduction
Lyme disease (LD) caused by Borrelia burgdorferi (sensu stricto) is the most commonly reported vector-borne disease in the United States. Bo. burgdorferi is a tick-borne pathogen believed to be transmitted exclusively by Ixodes spp. ticks (Bobe et al., 2021). Infection with Bo. burgdorferi can lead to multisystem manifestations affecting the skin, joints, nervous system, or the heart (Steere et al., 2016). It is believed that most cases recover following antibiotic treatment, but some go on to experience chronic health problems that can persist for years (Bobe et al., 2021).
While cases of LD are reported in most states in the southeastern United States each year, incidence rates are much lower there than in other regions such as New England and the Upper Midwest. This is attributed to multiple factors, including much lower Ixodes scapularis infection rates and different host-seeking behaviors in immature tick life stages (Ginsberg et al., 2021).
In the southern United States, the lone star tick (Amblyomma americanum) is by far the most abundant and aggressive human-biting tick species and is recognized as a vector of several pathogens (Childs and Paddock, 2003; Goddard and Varela-Stokes, 2009; Masters et al., 2008). However, the ability of A. americanum to acquire, maintain, and possibly transmit Bo. burgdorferi is still debated (Stromdahl et al., 2018; Stromdahl et al., 2015).
During the past 30 years, Bartonella species have been increasingly recognized as causes of human infections worldwide and responsible for diverse clinical manifestations (Breitschwerdt, 2017; Breitschwerdt et al., 2020; Cheslock and Embers, 2019). Bartonella spp. infect a wide range of mammalian host species and can be transmitted by many blood-feeding arthropods, including fleas, body lice, sand flies, bat flies, and mites; evidence is growing that ticks are also vectors of some Bartonella spp. (summarized by Breitschwerdt, 2017; Cheslock and Embers, 2019).
This report describes the discovery by our laboratory of a previously detected uncharacterized Bartonella sp. in several yellow flies (Diachlorus ferrugatus) collected from a homesite in southeastern Jacksonville in northeast Florida. Subsequent testing of lone star ticks collected by the resident at the same site revealed similar Bartonella DNA sequences in several ticks. Testing of blood samples from the resident at the homesite detected nearly indistinguishable Bartonella sequences in several samples collected over a 10-year period. Multiple samples from the same subject also tested positive for Bo. burgdorferi DNA.
Materials and Methods
Sample collection and testing
In September 2022, the University of North Florida (UNF) Public Health Research Laboratory began extracting DNA from yellow flies that had been collected in June 2020 from the southeast Jacksonville, Duval County, FL, residence of one (K.L.C.) of the authors, hereafter referred to as the case patient. The flies were collected with a commercial horse fly trap (Bite-Lite horse fly trap) and preserved individually in microtubes containing 100% ethanol (EtOH). The flies were identified phenotypically; notably, D. ferrugatus is the only known Diachlorus sp. in the study area.
DNA was extracted from individual flies using a commercial kit (Omega Biotek E.Z.N.A. Tissue DNA Kit, Norcross, GA), using the manufacturer's protocol for tissues, and eluted with 200 μL of elution buffer. Ticks tested for this study were collected by the patient, either attached to his body or clothing and acquired either from vegetation or from his pet cats over the previous 12 years. Collected ticks were stored in either 95% EtOH or 100% acetone before DNA extraction.
Ticks were also identified phenotypically based upon standard keys (Keirans and Durden, 1998; Keirans and Litwak, 1989). DNA from the ticks was extracted with different kits used at different times and included the MoBio UltraClean Tissue & Cells DNA Isolation Kit, GenElute (Sigma), Thermo GeneJET (Thermo Scientific), and a salting-out procedure using the MasterPure DNA Purification Kit (Lucigen). DNA from the ticks was eluted in Tris-EDTA buffer and stored at 0–4°C.
Human case data for the study were obtained per University of North Florida Institutional Review Board approval #468310-9. Blood samples from the patient were collected from January 2012 through October 2022 and a saliva swab sample was obtained in September 2022. Whole blood was obtained by finger or earlobe lancet sticks. Some whole blood samples were collected into small tubes containing the ethylenediaminetetraacetic acid anticoagulant and stored frozen at −20°C. Others were blotted onto Whatman 3M filter paper as dry blood spot (DBS) samples that were air-dried and stored individually in small, plastic zip-closure bags at 0–4°C. The saliva sample was collected using a BBL™ CultureSwab™.
Two small volumes of anticoagulated whole blood (12.5 or 25 μL) were used for DNA extraction and added directly to the lysis solution. The DBS samples and saliva swab were treated as tissue specimens. DNA extraction comparisons with dried blood included using 1, 2, or 3 individual 6-mm circular spots of blood-soaked filter paper. All patient specimens were extracted with reagents from the MasterPure Kit.
Two volumes of extracted DNA, 2.5 and 5 μL, were used for PCR testing of human samples. Negative control PCR samples containing nuclease-free water in the place of a template were included along with DNA extraction controls in each round of PCR testing at a rate of one control for every six experimental samples. To avoid the possibility of DNA amplicon contamination, no positive control samples were used in the present study.
Fly, tick, or human extracted DNA was tested by PCR for the presence of Bartonella species 16S-23S rRNA intergenic spacer (ITS), citrate synthase (gltA), and nuoG gene DNA and Bo. burgdorferi sensu lato (Bbsl) flaB gene DNA. The Bartonella ITS and nuoG gene PCR primers were previously described (Billeter et al., 2008; Colborn et al., 2010). Although described in a real-time PCR assay, the nuoG gene primers were used in a standard PCR.
The gltA gene nested PCR primers were derived from multiple reports (Birtles and Raoult, 1996; Billeter et al., 2012; Norman et al., 1995). Two Bbsl flaB gene PCR primers were combined in a single-reaction assay that amplifies a 284-bp product. The primers were as follows: 301F (5′-ACA-TAT-TCA-GAT-GCA-GAC-AGA-GG-3′) (modified from the study by Johnson et al., 1992) and 584R (5′-GCA-ACA-TTA-GCT-GCA-TAA-ATA-TTT-ACA-GC -3′) (this study).
Amplification parameters for all PCR tests began with a denaturation step at 94°C for 2.5 min. The Bartonella nested gltA PCR consisted of 30 amplification cycles per round at an annealing temperature of 48°C for 30 s in the primary reaction and annealing at 55°C in the nested reaction. The nuoG PCR consisted of 45 cycles with primer annealing at 56°C. For Bartonella ITS and Bbsl flaB PCRs, the first 10 cycles included denaturation at 94°C, 30 s, annealing for 2 min (66°C for Bartonella ITS and 60°C for the Bbsl flaB gene), and extension for 2 min at 72°C.
This stage was followed by 45 (for Bartonella ITS) or 40 (for Bbsl flaB) additional cycles at 92°C, 30 s, primer annealing for 30 s (64°C for Bartonella ITS and 56°C for Bbsl flaB), and extension for 30 s at 72°C. PCR products were electrophoresed in 2% agarose gels, including ethidium bromide, and visualized under UV light.
DNA sequence analysis
PCR products of the expected range for positive results were sequenced with both primers used for PCR. Sequences were aligned using Clustal (Larkin et al., 2007) and compared with sequences in the GenBank database using the Basic Local Alignment Search Tool (BLAST) (Altschul et al., 1990). Yellow fly-, lone star tick-, and human patient-associated Bartonella DNA sequences were assigned GenBank acc. nos. OQ180938–OQ180949. Tick- and patient-associated Bo. burgdorferi DNA sequences were assigned GenBank acc. nos. OQ158858–OQ158862.
A multiple sequence alignment of investigator-derived sequences combined with reference sequences was conducted with Clustal and utilized for phylogenetic analysis. Following a model test, a maximum likelihood analysis was conducted using the Kimura 2-parameter model with gamma distribution in MEGA11 (Kimura, 1989; Stecher et al., 2020; Tamura et al., 2021) with 500 bootstrap replicates (Felsenstein, 1985).
Results
None of the negative DNA extract control samples or PCR reagent negative controls tested positive during this study; therefore, there was no indication of any false-positive PCR results. None of the samples tested positive with Bartonella nuoG primers, and only one yellow fly sample was positive with the Bartonella gltA gene nested PCR test. All other positive results were obtained with either the Bartonella ITS primers or Bbsl flaB primers. These results are further described below.
Fly testing
During the early fall of 2022, as part of a pilot project, our laboratory began testing yellow flies for Bartonella species and Lyme Borrelia DNA. All of the flies tested negative for Bbsl flaB DNA. Surprisingly, however, Bartonella DNA was detected in 5 of 64 flies from the aforementioned patient's property. One fly (YF24) was negative with the ITS PCR assay, but positive with the gltA gene PCR assay; the sequence (GenBank acc. no. OQ191933) was 81% identical to a Bartonella sequence from a rodent in Japan (GenBank acc. no. AB290288.1) and 80% similar to several sequences from rodents in China (e.g., GenBank acc. no. MW161100.1).
The other four flies were negative with the gltA gene test, but positive for Bartonella ITS DNA. One fly sequence (YF2) was ∼86% identical to multiple GenBank sequences, including a Bartonella sp. sequence (GenBank acc. no. NG13-019) from a bat fly in Nigeria, clone 60 from an Aedes vexans mosquito from Europe (GenBank acc. no. MK301294.1), a lone star tick from Virginia (clone Hill-02-285, GenBank acc. no. EF559315.1), and two Bartonella tamiae sequences from human patients in Thailand (GenBank acc. nos. EF605284.1 and EF605283.1) (Table 1).
Description of Yellow Fly, Lone Star Tick, and Human Case Patient Specimens; Positive PCR Results for Bartonella and Borrelia burgdorferi; and Similarity (%) to Closest GenBank Match Using Basic Local Alignment Search Tool
ClarkAa1N, Amblyomma americanum nymph removed from patient's cat; KCAa1, A. americanum adult female removed from the patient; KCDBS or KLCDBS, dry blood spot sample from the patient; KCDBSARM, dry blood spot sample from patient's arm; KCFSPLA, plasma sample from patient's finger-stick blood; KCSASW, saliva swab sample from the patient; KLC12.5WB, 12.5-μL whole blood sample from the patient; KLCAa1M.4.2017, A. americanum adult male removed from the patient; KLCAa1M.5.2014, A. americanum adult male collected by patient; ND, not determined; NocAa1, A. americanum adult female from Nocatee Preserve; PS, poor sequence not submitted to GenBank; YF, yellow fly.
The other three yellow fly sequences (YF10, YF19, and YF45) were identical to each other and ∼99% identical to clone 60 and clone Hill-02-285, but only 89% identical to Ba. tamiae Th339 and Th307 (Table 1).
Tick testing
Finding Bartonella DNA in several flies led the authors to test previously extracted DNA samples from 24 lone star ticks collected by the case patient over the previous 12 years from the same residential property, from the UNF campus wildlife sanctuary, or from a nearby site in St. John's County (Nocatee Preserve). Three of the ticks, including a nymph removed from the patient's cat (ClarkAa1N) and an adult female removed from his skin (KCAa1)—both acquired from his residential property—and an adult female (NocAa1) from Nocatee Preserve, tested positive for Bartonella ITS DNA and the sequences were 99% identical to those from YF10, YF19, and YF45, the previously reported lone star ticks from Virginia, and the A. vexans from Europe (Table 1).
Two different ticks, including one (KLCAa1m.5.2014) removed from the patient's clothing in May 2014 and another (KLCAa1M.4.2017) removed from his scalp in April 2017, both acquired at either the patient's residence or the UNF campus, tested positive for Bbsl flaB DNA (GenBank acc. nos. OQ158861 and OQ158862). Those sequences were 99–100% identical to Bo. burgdorferi sensu stricto reference strains, B31 and N40, GenBank acc. nos. CP019767.1 and CP002228.1, respectively (Table 1).
The case patient
The 54-year-old, immunocompetent male patient had a 12-year history of chronic relapsing–remitting symptoms that started in April 2010 following several lone star tick bites (including one tick acquired from his property and several more from Fayette County, Georgia) in the week before symptom onset. Initial symptoms included only lateral cervical lymphadenopathy (Fig. 1A) and malaise. That event was followed months later with other symptoms, including severe fatigue, headache, somnolence, sleep disturbance, and occasional brain fog and stiff neck.

Those symptoms continued for years, remitting after various courses of oral antibiotics, then returning within weeks or a few months. Relapses occurred less often after several years. Bouts of fatigue and headache continued, sometimes occurring once monthly or every few months. After ∼10 years, he was taking antibiotics (usually 1–2 weeks of doxycycline) only once or twice per year, when symptoms were more severe or lasted longer than a week.
During this 12-year period, the patient maintained a regular regimen of vigorous exercise and he had no evidence of other underlying illness and rarely experienced other infections; he experienced influenza once and common cold symptoms only three times. Over the years, he regularly experienced bites from lone star ticks, and sometimes American dog ticks (Dermacentor variabilis), most of which were encountered at his property or the UNF campus.
During the fly and tick testing period of this study in the fall of 2022, coinciding with a period of high stress and poor sleep, the patient began experiencing severe chronic fatigue, headaches, multiple enlarged bilateral axillary lymph node swelling, and intermittent low-grade fever with chills. Some lymph nodes grew large and painful (Fig. 1B). His physician prescribed a 10-day course of ciprofloxacin. The lymph nodes returned to normal size, but fatigue lingered over the following two weeks.
Then, bilateral axillary lymphadenopathy returned, with multiple enlarged nodes on one side (Fig. 1C), again accompanied by headache, fatigue, and occasional fever and chills. He completed a weeklong course of doxycycline without dramatic improvement, but lymph node swelling slowly decreased to normal size over a period of another week. Following that course of antibiotics, lymph node swelling did not recur for the next two months.
Human patient testing
These events led to testing stored blood samples collected from the patient during the past 10 years. These samples were collected as part of ongoing investigations into his own illness and were typically collected during periods of relapse. None of the samples were positive in the Bartonella gltA test, but seven, including six DBS samples and one saliva swab, were positive for Bartonella ITS DNA (Table 1). The positive samples spanned the period from January 2012 to October 2022.
All of the samples were confirmed by sequencing and BLAST analysis to contain Bartonella species DNA. The patient sequences were nearly identical and were ∼99% similar to Bartonella sp. clone 60 from A. vexans and clone Hill-02-285 from A. americanum (Table 1). Nine of the same patient samples collected from January 2012 to January 2020 were positive for Bbsl flaB DNA. These included six DBS extracts, two additional frozen whole blood samples, and one frozen plasma sample (Table 1).
Three flaB sequences were submitted to GenBank; they were each 99–100% identical to the Bo. burgdorferi strain B31 or N40. Two DBS samples, one collected on January 5, 2012, and one collected on November 28, 2015, were positive for both Bartonella and Bo. burgdorferi DNA.
Bartonella ITS DNA sequence analysis
A multiple sequence alignment was performed with the yellow fly, tick, and human patient Bartonella ITS DNA sequences to demonstrate the relationship of sequences obtained in this study with the most similar Bartonella strain sequences in GenBank. This comparison included several aforementioned unpublished sequences from GenBank, the two A. americanum-derived sequences from Virginia, two Ba. tamiae sequences, and reference sequences of Bartonella apihabitans and Bartonella choladocola derived from honeybees (Fig. 2).

Multiple sequence alignment of Bartonella intergenic spacer sequences obtained from yellow flies, lone star ticks, blood samples from the case patient, and reference sequences. Sequences were aligned with Clustal. Asterisks indicate areas of agreement among all sequences. GenBank accession numbers follow the sequence ID names. Sequence ID names for sequences derived in this study are as follows: ClarkAa1N, Amblyomma americanum nymph removed from patient's cat; KCAa1, A. americanum adult removed from the patient; KCDBS, various dry blood spot samples from the patient; KCSASW, saliva swab sample from the patient; NocAa1, A. americanum adult from Nocatee Preserve; YF, yellow fly.
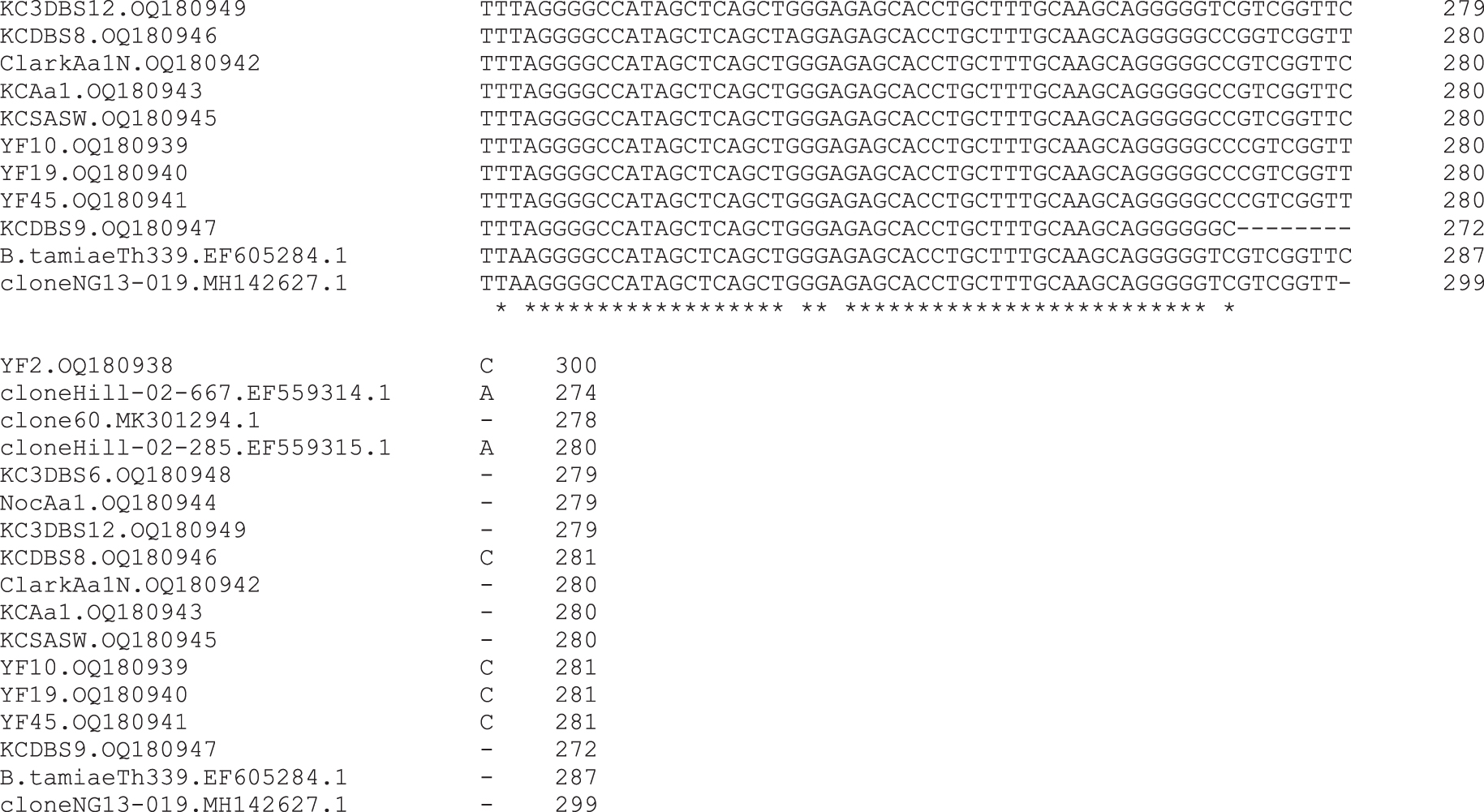
The ITS sequences from flies, ticks, and the human patient ranged from 279 to 281 bp, except for the YF2 sequence, which was 300 bp. These are some of the smallest ITSs identified among Bartonella species sequences in GenBank, but similar to the lengths of the spacers for the bee-associated and Ba. tamiae strains. Except for YF2, the yellow fly, tick, and patient sequences contained a similar, unique, single nt deletion at position 80 (using YF2 numbering as the referent) and a unique 19-nt gap starting at nt position 199.
These two unique features were shared with the sequences, clone 60 from A. vexans and clone Hill-02-285 from A. americanum (Fig. 2). Unfortunately, ITS sequences available in GenBank that are flanked by the utilized PCR primer set for other Bartonella species were too dissimilar to obtain an accurate multiple sequence alignment that included more Bartonella species.
For this same reason, we were not able to conduct a phylogenetic comparison of this study's Bartonella ITS sequences that included most previously characterized Bartonella species. However, a phylogenetic comparison was conducted using the maximum likelihood method to further demonstrate the relationships of newly discovered Bartonella sequences with the most closely related ones available in GenBank (Fig. 3).

Evolutionary analysis by maximum likelihood method. The evolutionary history was inferred by using the maximum likelihood method and Kimura 2-parameter model. The bootstrap consensus tree inferred from 500 replicates is taken to represent the evolutionary history of the taxa analyzed. Branches corresponding to partitions reproduced in less than 50% bootstrap replicates are collapsed. The percentages of replicate trees in which the associated taxa clustered together in the bootstrap test are shown next to the branches. Initial tree(s) for the heuristic search were obtained automatically by applying neighbor-join and BioNJ algorithms to a matrix of pairwise distances estimated using the maximum composite likelihood approach and then selecting the topology with superior log likelihood value. A discrete gamma distribution was used to model evolutionary rate differences among sites (five categories [+G, parameter = 0.1770]). This analysis involved 20 nucleotide sequences. There were a total of 304 positions in the final dataset. Evolutionary analyses were conducted in MEGA11.
The yellow fly, tick, and human patient ITS sequences reported here appear to be most closely related to each other, with low bootstrap support for separation among these sequences. They cluster most closely to the clone 60 sequence from an A. vexans in Europe and to one (clone Hill-02-285) of the two sequences obtained from lone star ticks in Virginia. The ITS sequences from most of our samples are more distantly related to YF2, Ba. tamiae, and honeybee-associated strains: Ba. apihabitans and Ba. choladocola.
Discussion
The name “yellow fly” refers to several species of biting flies in the family Tabanidae, including Diachlorus and Chrysops spp. One species, D. ferrugatus (Fabricius), is considered the true yellow fly (Fairchild and Weems, 2014), and our collections and references to yellow flies refer to D. ferrugatus specifically.
Within the United States, this species is reportedly distributed from Texas to New Jersey, but specific details on distribution and abundance within this range are lacking. This fly is a voracious blood feeder and persistent pest of humans, dogs, cattle, and other mammals. The bites are painful and can cause large extremely itchy welts (Fairchild and Weems, 2014). We could find no reports that D. ferrugatus is a known vector of any arthropod-borne pathogen.
There are a few records of Bartonella spp. detected in some biting flies, including Haematobia spp. and Stomoxys spp., from California (Chung et al., 2004). Our findings appear to be the first report of Bartonella in D. ferrugatus or other yellow flies. We were surprised when the Bartonella DNA sequences obtained from the flies matched closely with those previously reported from two lone star ticks from Virginia.
Those Bartonella sp. sequences were the only ones we could find in GenBank from any lone star ticks. They were initially described (Billeter et al., 2008) as being similar to Ba. tamiae, a species first isolated from three human patients in Thailand (Kosoy et al., 2008). Ba. tamiae DNA was later detected in multiple chigger mite pools and one Haemaphysalis tick pool from rodents in Thailand (Kabeya et al., 2010).
The human Ba. tamiae isolates were later shown to induce multiorgan pathology, including axillary lymphadenopathy, in immunocompetent mice, providing further evidence of Ba. tamiae's pathogenic potential. Although the Bartonella sp. detected in our study and strains from lone star ticks in Virginia are somewhat similar to Ba. tamiae, they likely represent a distinct sp.
Finding Bartonella DNA in yellow flies from the case patient's residence led us to test several dozen lone star ticks collected by him over the years, acquired either at his residence or nearby sites. Finding three A. americanum strains containing Bartonella ITS sequences nearly identical to that in the flies suggests a possible common host animal species present in the area. The patient had a history of numerous tick bites, mostly, but not exclusively, lone star ticks, as well as mosquito and yellow fly bites.
A Bartonella ITS sequence equally genetically similar to those from the yellow flies, lone star ticks, and the patient in Florida came from an A. vexans mosquito in Europe (unpublished GenBank submission). These findings raise questions regarding the ability of this as yet uncharacterized Bartonella sp. to possibly utilize multiple hosts and vectors.
A study of Bartonella in 80 dogs and fleas infesting them in Florida (Yore et al., 2014) did not report finding this species. Furthermore, it was not reported in multiple studies describing Bartonella detected in human patients in the United States (Breitschwerdt et al., 2020; Breitschwerdt et al., 2008; Breitschwerdt et al., 2007). Perhaps this Bartonella sp. has a restricted or highly focal host and vector distribution in the United States.
In the present study, we were unable to amplify Bartonella gltA or nuoG gene fragments from all except one gltA-positive sample using published primers for both genes. We surmise that this is due to significant mismatch of the identified strains with those primers. We improved the sensitivity of Bbsl flaB PCR compared with our previous studies (data not shown) by using PCR primers that amplify a smaller DNA fragment; additionally, sensitivity of the Bartonella ITS and Bbsl flaB PCRs was apparently improved by increasing the primer annealing and extension times in the first 10 cycles of amplification. It is possible that our DNA extraction and PCR methods allowed more sensitive detection of both organisms.
Detecting DNA sequences in flies or ticks does not prove the presence of live organisms or that the arthropods are able to maintain or transmit the organism (i.e., vector competence). Such findings might represent sensitive detection of dead organisms from a remnant bloodmeal. All of the arthropods that tested positive in our study were apparently flat bloodmeal-seeking species at the time of collection. A possible explanation for any positive PCR results is DNA amplicon contamination (false-positive results).
During this study, we utilized only single-reaction PCR assays and observed no positive results in any of our negative control samples. We obtained sequence-confirmed positive PCR results for Bartonella and Bo. burgdorferi from DNA samples extracted with different methods and reagents. We never found any similar Bartonella sp. in this laboratory before September 2022.
Finding similar Bartonella sp. DNA sequences in multiple specimens collected from the case patient over a 10-year period suggests that the organism was present and had established a persistent infection. Nevertheless, detecting the Bartonella sp. in the human specimens does not prove that it was responsible for the patient's long-term or more recent symptoms.
Complicating an explanation for the patient's clinical history was discovering the simultaneous presence of Bo. burgdorferi DNA in multiple samples covering the same 10-year period. Published evidence proving persistent Bo. burgdorferi infection post-treatment is lacking (Verschoor et al., 2022). There is also a lack of published evidence for chronic arthropod-borne coinfections in patients diagnosed with chronic LD (Lantos and Wormser, 2014). This could be partially due to the difficulty in detecting low concentrations of disseminated organisms in chronic infections.
The case reported here had suspected LD and had suffered chronic symptoms consistent with bartonellosis or Lyme borreliosis for over 12 years. Bartonella species are known for exhibiting relapsing low-level bacteremia in chronic infections, and therefore Bartonella DNA may be present in very low copy numbers in the blood of human patients with persistent signs and symptoms (Breitschwerdt et al., 2020; Maggi et al., 2020). Lyme Borrelia are similarly rarely present in the bloodstream following initial dissemination (Hyde, 2017), after which they establish persistent infection in tissues.
For human samples tested in this study, we utilized a salting-out DNA extraction method capable of high DNA recovery from DBSs and small DNA fragments (Cai et al., 2011; Molteni et al., 2013). Regardless of where in the body the identified organisms likely were present, either low numbers of viable cells or cell-free circulating DNA fragments of those organisms may have been present and detectable in peripheral blood specimens.
This patient had been treated with numerous brief rounds (1–2 weeks or months) of antibiotics such as doxycycline during the past 12 years. Even though symptoms would resolve for weeks or months following treatment, certain symptoms would return. Barring culture isolation of live Bartonella or Borrelia, and additional evidence for pathological effects, the cause of this patient's symptoms and the pathogenicity of the detected Bartonella sp. and Bo. burgdorferi strain(s) remain uncertain.
Ongoing studies involving testing of additional human patients from the southeastern United States with chronic, Lyme-like syndromic illness have led to detection of similar Bartonella sp. and Bbsl DNA sequences in some patients (unpublished observation); we anticipate reporting those data in separate articles.
In addition to the Bartonella sp., we confirmed Bo. burgdorferi DNA in two other adult A. americanum strains collected by the case patient. There is an ongoing debate over the ability of lone star ticks to acquire, maintain, and transmit Bo. burgdorferi (Stromdahl et al., 2018; Stromdahl et al., 2015). Bo. burgdorferi DNA has been detected in A. americanum from different states in multiple studies (Clark, 2004; Clark et al., 2013; Hudman and Sargentini, 2016; Rudenko et al., 2016; Stromdahl et al., 2001).
Rudenko et al. (2016) suggested that lone star ticks might exhibit selective compatibility with certain Bo. burgdorferi strains. Previous transmission experiments involving lone star ticks included strains different from those characterized from host-seeking lone star ticks in Florida in that study (Rudenko et al., 2016). We recommend further studies utilizing different Bo. burgdorferi strains and that those experiments be carried out to also determine the ability of adult A. americanum to transmit the spirochetes after being infected as larvae and nymphs.
There is a need for more studies on human patients with chronic undefined illness and on the presence of Bartonella and Bbsl in hematophagous arthropods and animal hosts in the southeastern United States. Some of these studies should include attempts to culture Bartonella and Borrelia spp. to fully characterize strains and provide material for transmission experiments.
Additionally, improved clinical diagnostic laboratory tests and testing capacity for Bartonella and Bbsl species are desperately needed. Recently, a new, droplet, digital PCR test for Bartonella detection in human clinical samples was described (Maggi et al., 2020). As human testing improves, it is likely that the spectrum of Bartonella and Borrelia species detected in acute and chronically ill human patients will grow.
Footnotes
Authors' Contributions
K.L.C. collected yellow flies, ticks, and patient blood specimens. K.L.C. and J.V.N. conducted PCR testing and DNA sequence analysis of specimens. Both authors drafted and edited the manuscript, table, and figures.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This work was supported by multiple donations from the Florida Lyme Disease Association and other private donors to K.L.C.'s UNF Foundation Research Account.