
Abstract
Background:
Cryptosporidium spp. are a type of protozoan parasite responsible for causing diarrheal illness worldwide. They infect a broad range of vertebrate hosts, including both non-human primates (NHPs) and humans. In fact, zoonotic transmission of cryptosporidiosis from NHPs to humans is frequently facilitated by direct contact between the two groups. However, there is a need to enhance the information available on the subtyping of Cryptosporidium spp. in NHPs in the Yunnan province of China.
Materials and Methods:
Thus, the study investigated the molecular prevalence and species of Cryptosporidium spp. from 392 stool samples of Macaca fascicularis (n = 335) and Macaca mulatta (n = 57) by using nested PCR targeting the large subunit of nuclear ribosomal RNA (LSU) gene. Of the 392 samples, 42 (10.71%) were tested Cryptosporidium-positive.
Results:
All the samples were identified as Cryptosporidium hominis. Further, the statistical analysis revealed that age is a risk factor for the infection of C. hominis. The probability of detecting C. hominis was found to be higher (odds ratio = 6.23, 95% confidence interval 1.73–22.38) in NHPs aged between 2 and 3 years, as compared with those younger than 2 years. Sequence analysis of the 60 kDa glycoprotein (gp60) identified six (IbA9 n = 4, IiA17 n = 5, InA23 n = 1, InA24 n = 2, InA25 n = 3, and InA26 n = 18) C. hominis subtypes with “TCA” repeats. Among these subtypes, it has been previously reported that the Ib family subtypes are also capable of infecting humans.
Conclusion:
The findings of this study highlight the genetic diversity of C. hominis infection among M. fascicularis and M. mulatta in Yunnan province. Further, the results confirm that both these NHPs are susceptible to C. hominis infection, posing a potential threat to humans.
Introduction
C
Cryptosporidium spp. is also one of the major causes of waterborne and foodborne illness outbreaks in industrialized countries (Chalmers and Giles, 2010; Efstratiou et al., 2017; Ryan et al., 2018). Up to now, small sub-unit rRNA (SSU rRNA), TRAP-C2, hsp70, cowp, and actin genes could be used for genotyping of the Cryptosporidium spp. (Hadfield et al., 2011; Jex et al., 2008; Roellig and Xiao, 2020).
The SSU has a multi-copy nature, thus it has been particularly commonly used to detect Cryptosporidium spp. (Le Blancq et al., 1997). However, some PCR assays may not achieve high specificity when applied to DNA samples of feces originating from various animals, suggesting that the primer sets designed are not entirely specific for Cryptosporidium spp. (Koehler et al., 2017). Thus, the novel bioinformatic approach-based sequencing of the LSU-PCR assay method is utilized for the identification of Cryptosporidium spp. This technique enhances the accuracy and efficiency of the identification process (Koehler et al., 2017).
Currently, over 44 species of Cryptosporidium have been identified, with the taxonomic classification of over 120 genotypes remaining uncertain (Feng et al., 2018; Ryan et al., 2021), of which C. hominis and C. parvum cause most of the infections reported in humans and non-human primates (NHPs) globally (Helmy and Hafez, 2022; Jia et al., 2022). Other Cryptosporidium species have been identified in NHPs, including C. muris, C. andersoni, C. ubiquitum C. suis, and C. meleagridis (Du et al., 2015; Feng and Xiao 2017; Karim et al., 2014; Li et al., 2017; Parsons et al., 2015; Ye et al., 2012).
The 60 kDa glycoprotein (gp60) as the genotyping marker is usually used in the laboratory to identify Cryptosporidium spp. subtypes. At present, eight subtype families of C. hominis (Ia, Ib, Id, Ie, If, Ii, Im, and In) have been described in NHPs. While Ia-If families are reported in humans, similar families such as Ib and Id are identified in cattle. Further, Ik is observed in horses and donkeys. whereas Ii, Im, and In families seem NHPs-adapted and are very rarely or not at all seen in humans (Abeywardena et al., 2012; Chen et al., 2019; Connelly et al., 2013; Giles et al., 2009; Jian et al., 2016; Karim et al., 2014; Laatamna et al., 2015; Li et al., 2019; Razakandrainibe et al., 2018; Ye et al., 2012; Zahedi et al., 2018).
A new family of Io subtype was reported in long-tailed macaques and rhesus macaques in Hainan Island, southern China by Zhao et al. (2019). However, there is controversy surrounding this subtype family, with some researchers suggesting that it should be classified as Im or In (Chen et al., 2019).
Macaca fascicularis and Macaca mulatta are two species of NHPs that are commonly used in biomedical research due to their genetic similarity to humans (Li et al., 2017; Zhang et al., 2014). The presence of Cryptosporidium spp. in these two NHPs heightens the likelihood of transmitting cryptosporidiosis to humans, posing a greater risk of infection. Thus, the molecular epidemiological investigation of Cryptosporidium spp. in NHPs is indispensable.
The objective of the present study was to determine the prevalence and subtypes of Cryptosporidium spp. infection in captive M. fascicularis and M. mulatta in Yunnan province. We aim at enhancing our understanding of the potential risks associated with Cryptosporidium spp. transmission for both humans and NHPs, ultimately contributing to improved public health measures.
Materials and Methods
Specimen collection
In June 2022, a total of 392 fecal samples were collected from M. fascicularis and M. mulatta. These samples were randomly collected from a commercial animal facility located in both Yuanjiang county and Kunming city in Yunnan province. It is important to note that all of these samples were collected after the monkeys had excreted their waste. Also, six monkeys experienced diarrhea in this study. All the monkeys were raised in individual cages.
In this experiment, the primates were collected from fecal samples and were reared separately according to their respective species. It was important to collect feces immediately after excretion and ensure that they were fresh and uncontaminated by feed or other disturbances. In addition, information about primates with diarrhea was recorded. At least 3–5 g fecal samples were collected from each monkey.
Then, the fecal samples were preserved in a sterile centrifuge tube (5 mL) with 2.5 mL DNA preservation solution (Phygene Biotechnology Co, Lid, Fu Zhou). Then, the collected fecal samples were quickly sent to the laboratory and stored in the −20°C refrigerator for DNA extraction in subsequent experiments. The data of fecal samples, such as ID numbers, sampling time, region, gender, age, and breed, were recorded.
Extraction of genomic DNA
Before performing DNA extraction, the fecal samples were centrifuged at 8000 g for 4 min to remove the DNA storage solution. Then, about 200 mg of feces were used to extract genomic DNA according to the instructions of the E.Z.N.A stool DNA Kit (OMEGA). In brief, the stool samples were first lysed with buffer and proteinase K. The resulting binding DNA was extracted using the HiBind DNA adsorption column. Finally, the DNA was eluted with an elution buffer. All the concentrations of the genomic DNA were measured and stored in a −20°C refrigerator for PCR amplification.
PCR amplification
All the DNA samples were screed for Cryptosporidium spp. by nested PCR and sequence analyses of the large subunit of the nuclear ribosomal RNA (LSU) gene. Through direct sequencing of amplicons, it was determined that there was no background or cross-amplification of a ∼500 bp product from any microbial taxon other than Cryptosporidium (Koehler et al., 2017). All primers and PCR systems used in this study are described in the literature.
The first cycle of primers was LSU2040F (forward: 5′-CGA ATA GCG TTA TCT TTG CTA TTT-3′) and LSU3020R (reverse: 5′-GTC TTC CGC GAA GAT CAG-3′) to amplify a region of ∼800 bp; the second cycle of primers was LSU2065F (forward: 5′-TTA CCATGG AAT YAG TTC AGC-3′) and LSU2557R (reverse: 5′-AAC ACC ATT TTC TGG CCA TC-3′) to amplify a region of ∼500 bp (Koehler et al., 2017). The PCR reaction was carried out in 25 μL using a standard reaction buffer, 3.0 mM of MgCl2, 200 μM of each dNTP, 50 pmol of each primer, and 1 U of Taq polymerase (MangoTaq, Bioline, London, UK).
The primary template was 2 μL, and the secondary template was 1 μL. The primary PCR was conducted with the following temperature settings: initial denaturation was 94°C for 5 min, followed by denaturation by 35 cycles of 94°C for 30 s, annealing by 58°C for 30 s, and 72°C for 50 s (extension) with a final extension of 72°C for 5 min. The conditions of secondary PCR were the same step, except that the extension step was 30 s instead of 50 s (Koehler et al., 2017).
Further, subtyping of C. hominis samples was amplified by the gp60 gene by the nested PCR method. The first cycle of primers was AL-3531 (forward: 5-ATA GTC TCC GCT GTA TTC-3) and AL-3534 (reverse: 5′-GCA GAG GAA CCA GCA TC-3′), and the second cycle of primers was AL-3532 (forward: 5-TCC GCT GTA TTC TCA GCC-3) and AL-3533 (reverse: 5′-GAG ATA TAT CTT GGT GCG -3′), which was used to amplify an ∼850-bp fragment of the gp60 gene (Peng et al., 2001).
If amplification failed, the C. hominis samples were further attempted for the gp60 gene using the AL-3531/AL-3533 and AL-3532/LX-0029 primer pairs (Sulaiman et al., 2005). The first cycle of primers is AL-3531 (forward: 5-ATA GTC TCC GCT GTA TTC-3) and AL-3533 (reverse: 5′-GAG ATA TAT CTT GGT GCG-3′); the second cycle of primers are AL-3532 (forward: 5-TCC GCT GTA TTC TCA GCC-3) and LX-0029 (reverse: 5′-CGA ACC ACA TTA CAA ATG AAG T-3′) to amplify a ∼400 bp fragment of the gene (Sulaiman et al., 2005).
The PCR reaction was carried out in 50 μL containing a standard reaction buffer, 3.0 mM of MgCl2, 200 μM of each dNTP, 200 nmol of each primer, 2.5 U of Taq DNA polymerase (MangoTaq, Bioline, London, UK), and 2 μL of nonacetylated BSA (10 mg/mL). The primary template and secondary template were 2 μL. The primary PCR temperature was set as follows: initial denaturation was 95°C for 3 min, followed by denaturation by 35 cycles of 94°C for 45 s, annealing by 59°C for 45 s, and 72°C for 60 s (extension) with a final extension of 72°C for 10 min.
The conditions of secondary PCR were the same step, except that the extension step was 50°C instead of 59°C (Feltus et al., 2006). The positive and negative controls for each test were added. Secondary PCR products were detected by electrophoresis on 1% (w/v) agarose gels containing ethidium bromide.
Sequence and phylogenetic analyses
All positive secondary PCR products were sequenced for the (LSU) gene by Sangon Biotech (TSING KE, Xi'an, China) in both directions, using an ABI 3730 Genetic Analyzer (Applied Biosystems, Thermo Fisher Scientific, Foster City, CA). The obtained sequences were assembled by the software ChromasPro 2.1.5.0 and aligned with corresponding sequences from the GenBank database using the Clustal × 1.83 to identify Cryptosporidium species.
The maximum-likelihood method was used to establish the phylogenetic tree using MEGA 7.0 with the Kimura 2-parameter model. The reliability of the tree was assessed using bootstrapping with 1000 replicates.
Statistical analysis
Statistical analysis was performed using a Chi-square (χ 2) test in SPSS version 26.0 for Windows (SPSS, Inc., Chicago, IL). A Chi-square test was conducted to evaluate the correlation between the Cryptosporidium spp. infection and the age, breed, gender, and location of the NHPs. The differences with p < 0.05 were considered statistically significant. The odds ratios with 95% confidence intervals (CIs) were performed to assess the strength of risk factors of Cryptosporidium spp. infection.
Results and Discussion
In this study, a total of 392 fecal samples were collected from NHPs. Among these samples, 42 (10.71%, 95% CI: 7.65–13.78) were found to be positive for C. hominis. All the C. hominis positive samples were detected in Yuanjiang county (Table 1). The prevalence of C. hominis in NHPs was found to vary among different age groups. Notably, the prevalence was observed to be higher in NHPs aged 2–3 years as compared with other age groups, with a statistically significant difference (p < 0.01) (Table 1).
Factors Associated with the Prevalence of Cryptosporidium hominis in Non-Human Primates
CI, confidence interval; OR, odds ratio.
The study also found that the prevalence of C. hominis was highest in female NHPs (13.38%, 95% CI: 7.78–18.98), whereas the lowest prevalence was observed in male NHPs (9.20%, 95% CI: 5.62–12.78). However, the difference in prevalence between the sexes was not found to be statistically significant (p > 0.05) (Table 1). To identify the C. hominis species in NHPs in Yunnan province, all the positive samples were selected for sequencing and analysis of the large subunit LSU rRNA gene.
Only C. hominis was distributed in all age groups of NHPs, showing strong host specificity. In addition, based on gp60 primers of two lengths, three C. hominis samples amplified ∼850 bp fragments, and the remaining 30 C. hominis samples amplified ∼400 bp fragments. Overall, 33 C. hominis samples were classified into six subtypes (Fig. 1), including the Ii family subtype (IiA17, n = 5), Ib family subtype (IbA9, n = 4), and In family subtype (InA23, n = 1, InA24, n = 2, InA25, n = 3, and InA26 n = 18) according to the “TCA” repeats.
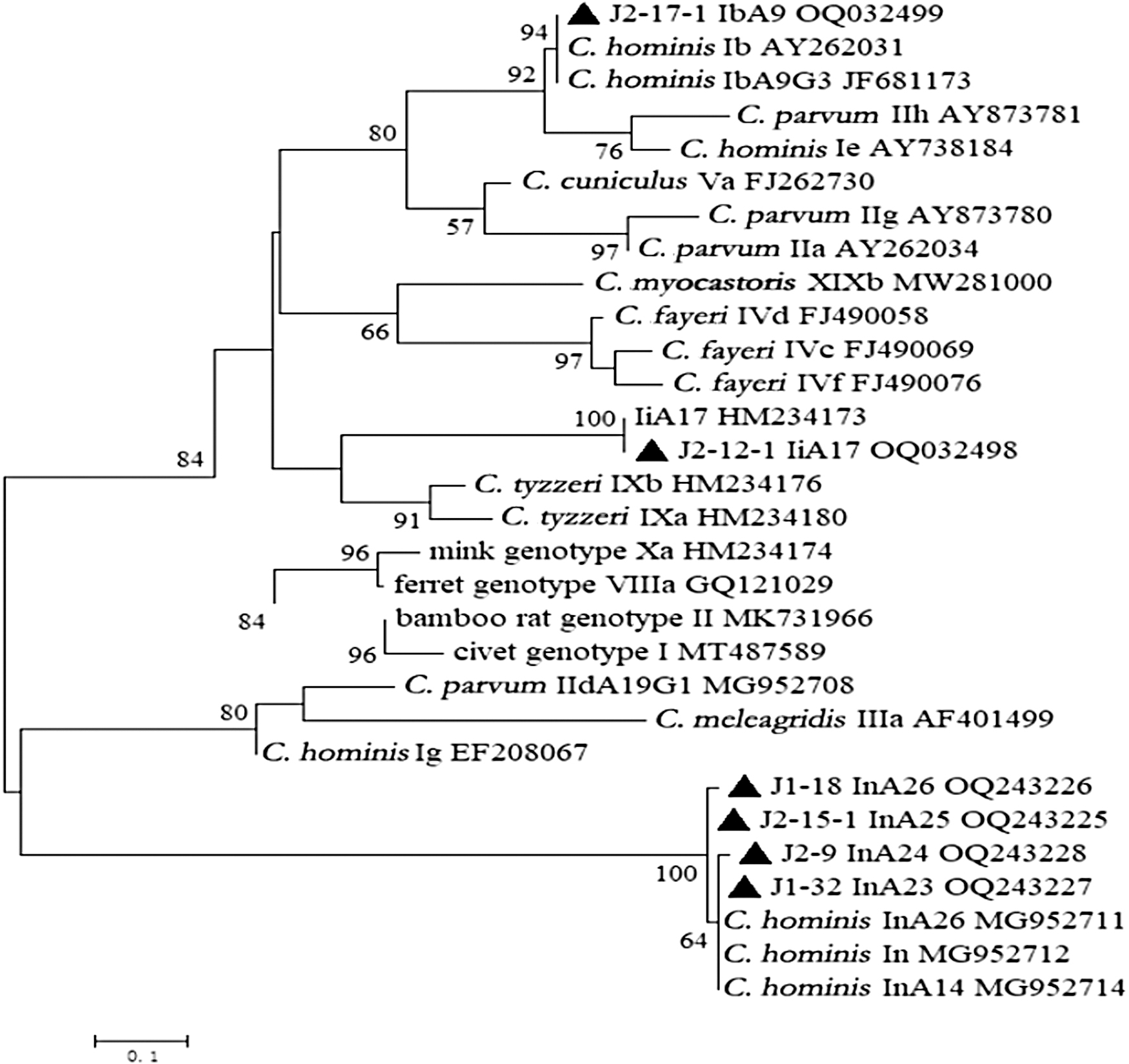
Phylogenetic analysis of Cryptosporidium spp. based on sequences of the gp60 gene fragment using the ML analysis method. Bootstrap analysis to assess the branch reliability, with 1000 replicates, and values above 50% are shown on nodes. Representative sequences obtained from this study are indicated with black filled triangles. ML, maximum-likelihood.
Based on statistical analysis, the prevalence of Cryptosporidium spp. detected in NHPs in China ranged from 0.6% to 14.4% (Chen et al., 2019; Du et al., 2015; Jia et al., 2022; Karim et al., 2014; Li et al., 2021; Shu et al., 2022). It was previously considered that only humans were susceptible to infection with C. hominis. However, due to the high genetic similarity between captive and free-living NHPs and humans, there are now predictions that they may also be susceptible to this infection.
The epidemiological investigation reveals that C. hominis is commonly found in chimpanzees, baboons, and various species of macaques (Chen et al., 2019; Feng et al., 2011; Karim et al., 2014; Li et al., 2011; Parsons et al., 2015; Widmer et al., 2020; Ye et al., 2012). Notably, monkey-adapted genotypes of the Cryptosporidium spp. primarily infect captive animals, as demonstrated in Table 2.
Reports Describing the Occurrence and Molecular Diversity of Cryptosporidium hominis in Captive and Free-Living Non-Human Primates Global
The prevalence of Cryptosporidium spp. in NHPs in the current study was found to be higher than previously reported rates in various regions of China, including Henan province (0.6%, 5/786), Guangxi Zhuang autonomous region (1.0%, 11/1,079), Guangdong province (1.8%, 1/57), Shanghai city (0.7%, 2/290) (Karim et al., 2014), Qinling Mountains (3.0%, 6/197) (Du et al., 2015), Yunnan province (0.6%, 2/348) (Shu et al., 2022), Guiyang city (0.7%, 18/2402) (Jia et al., 2022), and Hainan province (9.1%, 132/1452) (Chen et al., 2019).
In addition, the prevalence in the current study was also higher than that reported in captive NHPs from the chimpanzee in Comoé National Park, Côte d'Ivoire (0.8%, 1/124) (Köster et al., 2022 ), the captive NHPs from six European zoos (0.9%, 4/454) (Köster et al., 2022 ), a wild chimpanzee population in Dindefelo, Senegal (0.9%, 2/234) (Köster et al., 2021), but it was lower than that in Guangxi autonomous region in China (14.4%, 52/360) (Li et al., 2021).
Previous studies demonstrate that many factors may contribute to the Cryptosporidium spp. infection, including but not limited to breeds, detection methods, geographical location, sample sizes, age, consumption of drinking water, and exposure to recreational water sources (Hailu et al., 2022; Osman et al., 2016; Ryan et al., 2021; Thompson and Ash, 2019). The elevated incidence of Cryptosporidium spp. identified in M. fascicularis and M. mulatta within the study area implies that additional precautions should be implemented to prevent the transmission of C. hominis from NHPs to humans.
Similarly, in another study, Cryptosporidium spp. was commonly distributed in all age groups of NHPs. The highest prevalence of C. hominis was detected in NHPs aged 2–3 years old (25.35%, 95% CI: 15.23–35.47), and the difference was statistically significant (p < 0.01, Table 1). The NHPs aged between 2 and 3 years old are at a higher risk of contracting C. hominis infection due to their transition from breast milk to other foods that lack maternal antibodies.
This lack of immunity increases their susceptibility to the infection. As a result, the prevalence of C. hominis is lower in NHPs under 2 years old who are still nursing and receiving maternal antibodies (Table 1). Moreover, there is no C. hominis positive found in NHPs in Kunming city (Table 1). This difference may be associated with the different feeding conditions and lower sample sizes (n = 47).
Up to now, 10 species (C. parvum, C. hominis, C. cuniculus, C. andersoni, C. bovis, C. felis, C. meleagridis, C. muris, C. suis, C. ubiquitum) of Cryptosporidium spp. have been reported in NHPs (Hailu et al., 2022; Karim et al., 2014; Sak et al., 2014; Sak et al., 2013). In this study, only C. hominis was identified in M. fascicularis and M. mulatta. C. hominis is typically considered a species that is unique to humans, but recent research has shown that it is being detected more frequently in other animals such as NHPs, horses, pet red squirrels, dairy cattle, yaks, and donkeys (Deng et al., 2020; Feng et al., 2018; Qi et al., 2015; Zhang et al., 2015).
The detection of C. hominis in this study highlights the potential risk of cross-species transmission between M. fascicularis and M. mulatta. This finding suggests that these NHPs may serve as a reservoir for the infection and could potentially transmit the disease to humans or other animals (Widmer et al., 2020; Zhao et al., 2019). To prevent the spread of C. hominis infection in NHPs, animals, and humans, it is crucial to implement effective strategies and measures.
Among six subtypes, the Ib family of C. hominis was reported in captive baboons in Kenya (Li et al., 2011; Ye et al., 2012), which also was found to be the cause of an outbreak of cryptosporidiosis in a pediatric hospital in China (Feng et al., 2012). IbA9 is a subtype belonging to the Ib family of C. hominis, which has been detected in M. fascicularis and M. mulatta during this study.
These results suggest that there is a possibility of transmission of this subtype (IbA9) from NHPs to humans. Another subtype IiA17 was previously identified in a rhesus macaque in the USA (Feng et al., 2011) and long-tailed macaques and rhesus macaques in Hainan Island of China (Zhao et al., 2019). In addition, the IiA17 was also found in humans (Lebbad et al., 2018). Phylogenetic analysis revealed that the IiA17 subtype formed a distinct cluster within the tree (Fig. 1), indicating that it may represent a unique subtype among NHPs.
More studies are needed for investigating the IiA17 in NHPs and other animals throughout different geographical regions. Moreover, four new In family subtypes (InA23-InA26) were identified in M. mulatta in this study (Fig. 1). The In subtype family of C. hominis was identified as a new subtype in M. fascicularis in the Hainan province of China in 2019 (Chen et al., 2019). However, as far as current knowledge goes, the ln family appears to only infect NHPs. Further systematic epidemiological studies will be required to confirm whether these subtypes have the potential to infect humans.
Conclusion
The findings of the present study revealed the infection of human-pathogenic C. hominis in M. fascicularis and M. mulatta in Yunnan province, China. The total prevalence of C. hominis was 10.71% (42/392), which was significantly higher than those reported in previous studies on NHPs. C. hominis infection is associated mainly with diarrhea, nausea, and vomiting.
Therefore, greater emphasis should be placed on investigating C. hominis infection in NHPs in future studies. The findings of this study have provided additional evidence for the existence of six distinct genotypes of C. hominis, highlighting the genetic variability of C. hominis infection among NHPs in China.
Availability of Data and Material
The datasets presented in this study have been uploaded in GenBank with the accession numbers: OQ207290-OQ207294 for C. hominis. The obtained sequences based on the gp60 gene were uploaded, and the accession numbers were OQ243225-OQ243228, and OQ032498-OQ032499.
Ethical Approval
The present study was reviewed and approved by the Animal Ethics Committee of Lanzhou Veterinary Research Institute, Chinese Academy of Agricultural Sciences. Fecal samples were collected from M. fascicularis and M. mulatta with the permission of the breeders and managers. All procedures were conducted strictly by the requirements of Animal Ethics Procedures and Guidelines of the People's Republic of China.
Footnotes
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
Project support was provided by The Natural Science Foundation of Gansu Province, China (no. 21JR7RA027), the Gansu Agricultural University, grant no. 32160843, and the National Natural Science Foundation of China (32172878).